
Abstract
Adult-onset neuronal ceroid lipofuscinosis (ANCL), one of the neuronal ceroid lipofuscinosis (NCLs), is an inherited neurodegenerative disorder with progressive neuronal dysfunction. Recently, mutations in the DNAJC5 gene that encodes cysteine-string protein alpha (CSPα) have been reported to be associated with familial autosomal-dominant ANCL (AD-ANCL). This study constructed an ANCL transgenic zebrafish model expressing the human mutant DNAJC5 (mDNAJC5) gene under the control of a zebrafish neuron-specific promoter. To investigate whether gene therapy based on genome-editing technology could treat ANCL, a panel of TALEN and Cas9 nucleases was designed to disrupt the mDNAJC5 gene in this transgenic animal model. By screening these nucleases, it was found that one nuclease that targeted the 5′ coding region efficiently alleviated mDNAJC5 protein aggregates in the affected neurons. Therefore, this study provides a gene therapy strategy via the use of the CRISPR/Cas9 system to treat neural genetic diseases.
Introduction
N
Zebrafish has been successfully used in a broad range of research, including evolution, development, toxicity, and human diseases. 7,8 Researchers have developed a series of toolkits to modify the zebrafish genome with gene disruptions or transgenes. Tol2, a transposon developed from medaka fish, is the most widely used tool in making transgenic zebrafish because of its high genomic integration efficiency. 9 The introduction of a multisite gateway toolkit for Tol2 vector construction enables the quick and efficient assembly of multiple elements in a single Tol2 transposon backbone, which significantly facilitates transgenic research. 10 –12
Importantly, in recent years, powerful genome editing tools, such as transcription activator-like effector nucleases (TALENs) and clustered regularly interspaced short palindromic repeats (CRISPR)/Cas9, have enabled investigators to modify genomes in a wide range of cell types and organisms. TALENs are paired chimera nucleases, each of which is composed of the programmable sequence-specific DNA-binding module and the nonspecific DNA-cleavage domain derived from the Fok1 endonuclease. 13 Upon binding to their targets, TALENs form heterodimers and produce DNA double-strand breaks (DSBs) in regions between their binding sites. Unlike TALENs, the CRISPR system binds and cleaves the DNA through a programmable short guide RNA pairing one strand of its target and a Cas9 protein recognizing a protospacer-adjacent motif (PAM) that is usually a unique sequence adjacent to the RNA binding site of the non-target strand. 14 DSBs can be repaired either by non-homologous end-joining (NHEJ) to yield indels 15,16 or by homologous directed recombination (HDR) if homologous donor templates are provided. 17,18 Due to their abilities to be reprogrammed to recognize a relatively large variety of DNA sequence, these engineering nucleases have been widely used in basic research 19 –31 and hold great promise for clinical applications, especially in the treatment of genetic diseases. 22,32 –34
This study generated a transgenic ANCL zebrafish model in which the mutant human DNAJC5 gene was expressed under the control of the HuC neuron promoter. In order to eliminate the disease-specific mDNAJC5 protein aggregates, a series of TALENs and Cas9/sgRNAs were designed to destroy DNAJC5 expression. It was found that targeting the mutant site of the DNAJC5 gene could efficiently induce DSBs and indel mutations, which led to the truncations of the mDNAJC5 protein but did not eliminate protein aggregates, suggesting that the N-terminal portion of the DNAJC5 protein is responsible for protein misfolding and aggregation. Targeting the 5′-coding region caused a significant reduction of protein aggregates, which were demonstrated due to nonsense-mediated mRNA decay mechanism. Thus, this study provides a gene therapy strategy to treat neural genetic diseases via the use of genome editing technology.
Materials and Methods
Zebrafish care
Zebrafish used in this experiment were AB line. Adult fish and embryos were raised and maintained according to Kimmel et al. 35 Healthy 3- to 5-month-old adult female and male fish were selected for mating once a week.
Vector constructions
For the construction of pDestTol2pA-HuC-EGFP-DNAJC5 and pDestTol2pA-HuC-mCherry-mDNAJC5 plasmids, a DNAJC5 fragment was amplified by polymerase chain reaction (PCR) using following primers: attB2F—5′-GGGGACAGCTTTCTTGTACAAAGTGGACATGGCAGACCAGAGACAGCG-3′; and attB3R2—5′-GGGGACAACTTTGTATAATAAAGTTGATTAGTTGAACCCGTCAGTGT-3′. mDNAJC5 was generated by overlapping PCR described by Ho et al. 36 The site-directed mutagenesis primers was mF2: 5′-TTCTGCGGCCTCACGTGCTGCTACTGCTGCT-3′ and mR1: 5′-AGTAGCAGCACGTGAGGCCGCAGAAGACAAA-3′. To generate the pDestTol2pA-HuC-EGFP-DNAJC5 and pDestTol2pA-HuC-mCherry-mDNAJC5 plasmids, multisite recombination reactions were performed, as described by Kwan et al. 10 The entry vectors pDestTol2pA, p5E-HuC, pME-EGFP, and pME-mCherry were kindly gifted from Shuo Lin. p3E-DNAJC5 and p3E-mDNAJC5 plasmids were constructed according to the protocol provided by Life Technologies. Briefly, equimolar amounts of destination vectors pDestTol2pA, p5E-HuC, pME-EGFP (or pME-mCherry), and p3E-DNAJC5 (or p3E-mDNAJC5) vectors were incubated in a final volume of 5 μL reactions in the presence of LR Clonase II Plus Enzyme Mix. The ligation products were then transformed into Escherichia coli Top10.
For the construction of pDestTol2pA-HuC-mCherry-mmDNAJC5 plasmid, the site-directed plasmid mutagenesis method was used according to Liu et al. 37 In brief, the whole-length pDestTol2pA-HuC-mCherry-mDNAJC5 plasmid was amplified by PCR using primers containing the mutagenesis site. Next, the PCR product was transformed into E. coli Top10 after being treated with 5 units of DpnI at 37°C for 2 h. Five colonies were selected for sequencing to determine whether desired mutations were generated. The site-directed plasmid mutagenesis primers were: mmF—5′-AGACAGCGCTCACTGTCTACTCTGGGGAGT-3′; mmR—5′-GTAGACAGTGAGCGCTGTCTCTGGTCTGCC-3′.
Generation of transgenic zebrafish
To create transgenic zebrafish, pDestTol2pA-HuC-EGFP-DNAJC5 or pDestTol2pA-HuC- mCherry-mDNAJC5 plasmid was mixed with Tol2 transposase mRNA to microinject into one-cell stage fertilized zebrafish embryos by using an electronically regulated air-pressure micro-injector (Harvard Apparatus; PL1-90). Larvas with EGFP or mCherry expression in the spinal cord were selected at 3 days post fertilization (dpf) after injection and raised to adulthood. Individual F0 founders were out-crossed with wild-type zebrafish, and their F1 progenies were identified for the presence of EGFP or mCherry expression. Finally, F1 fish were used to generate stable transgenic zebrafish line. These transgenic zebrafish were used for further experiments.
Design TALENs and Cas9 for targeting the mDNAJC5 gene
Potential TALEN target sites were designed with an online tool, TALENT (
For CRISPR/Cas9, the targets sites were designed with an online tool, ZIFIT Targeter (
In vitro mRNA synthesis
Cas9 and TALEN expression vectors were linearized by XbaI and NotI, respectively. Next, the purified linearized templates were in vitro transcribed to mRNA with mMESSAGE mMACHINE T7 ULTRA kit (Ambion), and the purified sgRNAs PCR templates with T7 promoter were transcribed by T7 polymerase (NEB). RNAs were purified by LiCl acetate precipitation and re-dissolved in RNase-free water. The RNA quality and concentration were analyzed by electrophoresis and nucleic acid spectrometer, respectively.
Zygote microinjection
Healthy 3- to 5-month-old wild-type female fish and mDNAJC5 transgenic male fish were crossed, and the resulting embryos were collected for microinjection at one-cell stage. For injection of TALENs, equal amount of left and right TALEN mRNAs (∼300 ng/μL) were mixed and injected together into the cytoplasm of one-cell stage zebrafish embryos. For CRISPR/Cas9 injection, Cas9 mRNA (300 ng/μL) and sgRNA (60 ng/μL) were co-injected into one-cell stage embryos. For transient expression, plasmids pDestTol2pA-HuC-mCherry-mDNAJC5 or pDestTol2pA-HuC-mCherry-mmDNAJC5 mixed with pDestTol2pA-HuC-EGFP-DNAJC5 were co-injected into one-cell stage wild-type AB embryos.
Mutation analysis
Genomic DNA was extracted using the alkaline lysis method. Briefly, about five 48–72 h zebrafish embryos were collected in 30 μL of alkaline lysis buffer (50 mM of NaOH) and incubated at 95°C for 10 min and cooled on ice. Next, 1/10 volume neutralization buffer were added to neutralize the alkaline (1 M of Tris-HCl, pH 8.0). The target sites of TALEN and Cas9/sgRNAs were amplified by PCR with specific primers. The PCR products were subjected to Sanger sequencing to identify potential indel mutations. The primers were: DNAJC5 sF—5′-CCCACAACGAGGACTACACC-3′; and DNAJC5 sR—5′-GCCTTGGGCTTACACTTCCC-3′.
Real-time PCR for analyzing mDNAJC5 gene expression
Real-time PCR (RT-PCR) was performed with SsoAdvanced SYBR Green Supermix (Bio-Rad) by using a Bio-Rad CFX96 Real-Time system. The PCR conditions were programmed as follows: 40 cycles of 10 s at 95°C and 20 s at 60°C after initial denaturing for 30 s at 95°C. The primers for RT-PCR were: RTF—5′-ACACCTACTTCGTGCTGTCC-3′; and RTR—5′-GTATGACGATCGGCGTGTCT-3′.
Results
Construction of transgenic zebrafish with neuron-specific expression of mutant DNAJC5
In order to establish an ANCL animal model, transgenic zebrafish with neuron-specific expression of mDNAJC5 were generated (Fig. 1A). As shown in Fig. 1B and Supplementary Fig. S1, the transgenic vectors were constructed with the multisite gateway system. The wild-type DNAJC5 and mDNAJC5 were fused directly to the C-terminal of GFP or mCherry, respectively, and controlled by HuC promoter. The endo- and RNase-free plasmids of HuC-mCherry-mDNAJC5 or HuC-EGFP-DNAJC5 were injected together with Tol2 transponase mRNAs into zebrafish embryos at the one-cell stage. In embryos injected with HuC-EGFP-DNAJC5, EGFP-DNAJC5 was expressed ubiquitously in the neurons, while in embryos injected with HuC-mCherry-mDNAJC5, the mCherry reporter was expressed as aggregates, which is consistent with previous observations in mammalian cells (Fig. 1C). The mCherry-positive embryos were collected and raised to adult. They were then out-crossed with wild-type AB fish to identify potential founders. Two independent founders harboring mCherry-mDNAJC5 transgene were identified, and the one with a stronger mCherry-mDNAJC5 expression was kept for further experiments.
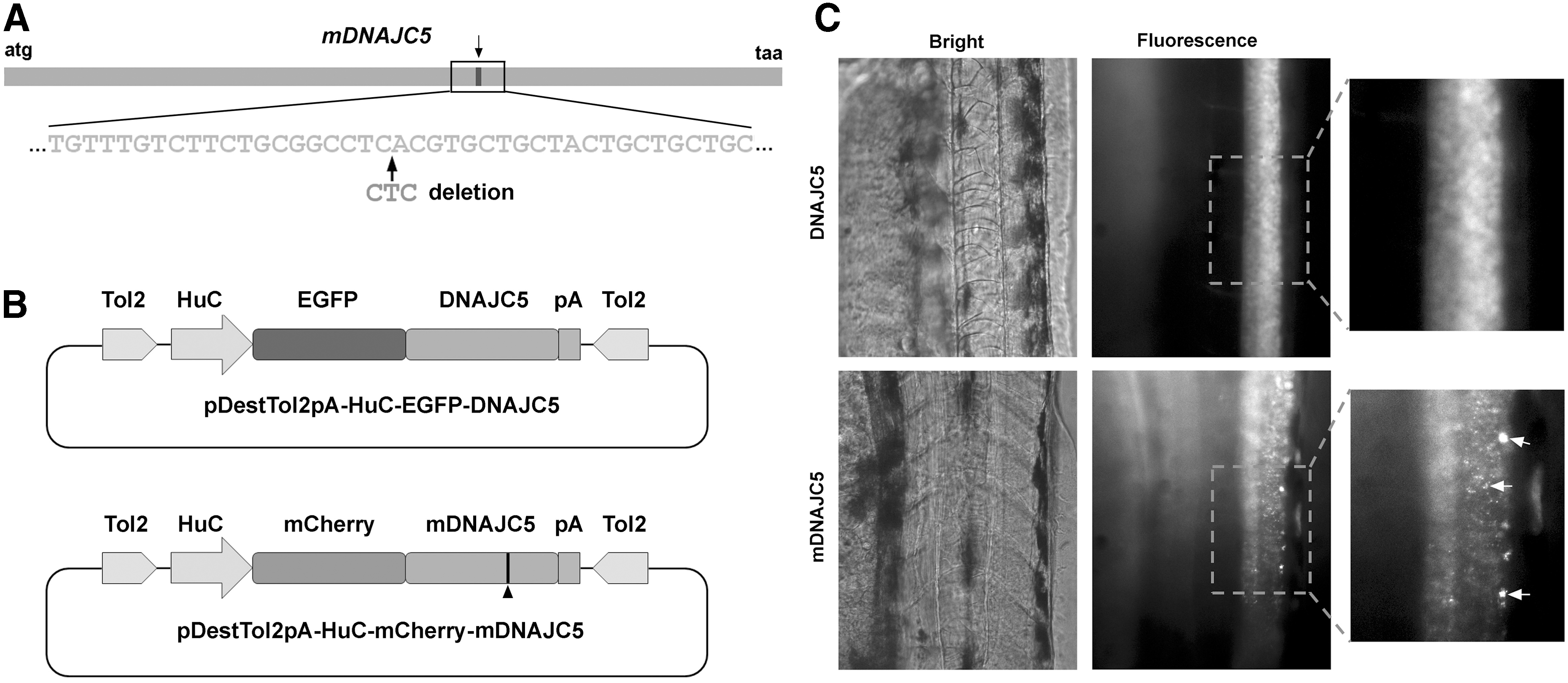
Construction of adult-onset neuronal ceroid lipofuscinosis (ANCL) zebrafish model.
Targeting mDNAJC5 by TALEN
To eliminate the mutant protein aggregates in ANCL zebrafish model, an attempt was made to disrupt the mutant gene by a pair of TALENs, one of which is specific to the mutation site. It was hypothesized that removing the C-terminal amino residues from mDNAJC5 protein would increase the solubility of mDNAJC5 protein and thus reduce protein aggregates. The TALEN targeting sites were designed as shown in Fig. 2A. The left TALEN arm recognizes a 17 bp DNA motif, which covers the mutation site (ΔCTC), and the right arm recognizes another 17 bp motif downstream of the left arm. The space sequence is 15 bp, which has been demonstrated to be optimal for the activity of TALEN with a 63 amino-acid linker between TALE module and FOK1 domain. It was expected that this pair of TALENs could induce indel mutations within its target site and lead to mDNAJC5 proteins losing their C-terminal amino residues and becoming soluble. mRNAs for each TALEN arm were in vitro transcribed and were microinjected in pairs into heterozygous mDNAJC5 transgenic zebrafish embryos. Following injection, zebrafish embryos at 2 dpf were examined under a fluorescence microscope to see if mCherry-DNAJC5 aggregates become soluble. However, beyond expectations, none of the TALENs-injected embryos showed any reduction in the level or territory of protein aggregates (Fig. 2B). Next, the study examined if this pair of TALENs induced DSBs and indel mutations within its target site. Genomic DNA of injected ACNL zebrafish embryos was extracted and subjected to PCR to amplify the TALEN target site. Sanger sequencing of the PCR products revealed that the TALEN worked very well and induced multiple indel mutations within mDNAJC5 genes (Fig. 2C). Theoretically, two-thirds of these indels result in frameshift mutations in the mDNAJC5 coding region, which will eliminate the expression of its C-terminal amino residues. Thus, this observation suggests that the C-terminal region is dispensable for protein aggregation and that the strategy designed to remove the C-terminal region of mDNAJC5 cannot relieve the ACNL phenotype.

ANCL zebrafish model treated by transcription activator-like effector nucleases (TALENs).
Targeting mDNAJC5 by CRISPR/Cas9
Next, the strategy was changed to target the upstream region of mDNAJC5 by using CRISPR/Cas9. Five sgRNAs targeting non-overlap sites that are sequentially upstream of TALEN target site were designed (sgRNA 1–5). Each of the five sgRNAs was microinjected together with Cas9 mRNA into mDNAJC5 transgenic embryos at the one-cell stage. Next, at 48 hpf, genomic DNA was extracted from a pool of five injected embryos and analyzed by PCR and sequencing. It was found that sgRNA3 and sgRNA5 induced significant indel mutations in their target sites, as evidenced by the presence of a composite sequence trace in the sequencing graph (Fig. 3A–C). The remaining three sgRNAs did not induce any detectable indels. Next, the focus was on these two effective sgRNAs to see if they could help to reduce protein aggregates. As with the TALEN-injected embryos, no reduction in protein aggregates was found in sgRNA3-injected embryos, as evidenced by mCherry expression (Fig. 3D), suggesting that removal of the C-terminal portion that is downstream of the sgRNA3 target site is not enough to improve the solubility of the mDNAJC5 protein. In sgRNA5-injected embryos, it was observed that 63.5% of them showed a significant reduction in protein aggregates (Fig. 3E). Unexpectedly, these protein aggregates nearly disappeared rather than became soluble. The remaining fish that had no visible reduction in protein aggregates had little or no indel mutations, which could be the result of injection failure (Supplementary Fig. S2).

ANCL zebrafish model treated by CRISPR/Cas9.
Because mCherry was fused to the N-terminal of mDNAJC5 in the transgenic zebrafish, indel mutations in mDNAJC5 coding region were not supposed to interrupt mCherry expression. The remaining elements within the transgenic vector that were necessary for mCherry expression were checked, including the HuC promoter, mCherry coding region, and ployA signal, to see if they harbored potential sgRNA5 off-target site and therefore were destroyed by sgRNA5. The sequence composed of 3′ 10nt portion of sgRNA5 (TCTACCTCTG) and spCas9 PAM (NGG) was used for searching potential off-target sites. However, no such off-target site was found in the DNA sequences of these elements, suggesting that the elimination of mCherry-DNAJC5 is likely due to indel mutations in sgRNA target site within the DNAJC5 gene.
Cas9 Targeting the 5′ coding region of mDNAJC5 decrease its mRNA level
Because random indel mutations induced by sgRNA5 can cause a premature stop codon in mDNAJC5, it was hypothesized that the reduced expression of mCherry-mDNAJC5 is due to a nonsense-mediated mRNA decay pathway. This pathway is an important mRNA surveillance mechanism in eukaryotic cells, which selectively degrades mRNAs carrying a premature translation termination codon to prevent generating the harmful truncated proteins. To test this hypothesis, RT-PCR was performed to detect the relative expression level of mDNAJC5 in sgRNA5-injected embryos. The result showed that the mDNAJC5 mRNA level declined ∼40% in the embryos that received the Cas9/sgRNA5 injection (Fig. 4A). To confirm this observation further, a mutant mDNAJC5 (mmDNAJC5) plasmid was constructed that harbors a 1 bp deletion in the upstream coding region of mDNAJC5 (31 del C), which leads to a premature stop codon and genocopies frameshift indels induced by sgRNA5 (Fig. 4B). mCherry-mmDNAJC5 or mCherry-mDNAJC5 plasmid was mixed with an equal amount of EGFP-DNAJC5 plasmid, and they were co-injected into zebrafish embryos at the one-cell stage. It was found that embryos injected with mCherry-mDNAJC5 and EGFP-DNAJC5 plasmids showed comparable EGFP and mCherry expression in the neurons, while embryos injected with mCherry-mmDNAJC5 and EGFP-DNAJC5 plasmids displayed strong EGFP expression and yet a very weak mCherry expression, indicating that a frameshift mutation in this site resulted in decreased expression (Fig. 4C). In addition, the sgRNA5-injected embryos were raised to adulthood and a one-line harboring frameshift mutation was obtained around the sgRNA target site (mDNAJC5Δ1, Fig. 4D). The offspring of this line have an extremely weak mCherry signal (Fig. 4F). This observation could not be explained by the redistribution of the mCherry proteins into the cytoplasm. More importantly, compared to mDNAJC5 embryos, mDNAJC5Δ1 embryos had significant reduced DNAJC5 mRNA expression (Fig. 4E). Together, these results suggest that the reduction of mcherry-mDNAJC5 protein aggregates was due to the decreased mRNA level, which was like that which resulted from the nonsense-mediated mRNA decay pathway.

Cas9/sgRNA5 decreased DNAJC5 mRNA level.
Discussion
ANCLs are a common group of inherited neurodegenerative diseases with an onset age range from 25 to 46 years. 6 As with other inherited neurodegenerative diseases, the aggregation and deposition of misfolded proteins are accumulated in the affected neurons to induce endoplasmic reticulum stress and reactive oxygen species toxin, which eventually leads to dysfunction or death of the neuron. 6,38 Many of these misfolded proteins are due to the genetic mutations in their coding regions. Therefore, gene therapies by correction or removal of these genetic lesions hold the promise of curing such disorders.
Recent advances in engineering nucleases have provided a great opportunity for the treatment of such genetic disorders. The development of a series of genome editing tools such as ZFNs, TALENs, and CRISPR/Cas9 has made it possible for both in vivo and in vitro genome editing in an efficient way. Recently, Yin et al. adopted a hydrodynamic delivery way to inject Cas9 plasmid and sgRNA along with a repair template into the liver of an adult mouse model of tyrosinemia to correct the mutant Fah gene, and they rescued about 1/250 cells expressing the wild-type Fah protein. 34 In addition, the CRISPR/Cas9 system has been employed to correct a dominant mutant Crygc gene that causes cataracts in the mouse by co-injection into embryos of Cas9 mRNA and sgRNA. 18 The same research group has successfully used the CRISPR/Cas9 system to correct the disease-causing gene Crygc –/– in spermatogonial stem cells to produce healthy pups. 32
In one of the ANCLs, the DNAJC5 gene was identified to be frequently mutated at two positions (L115R and ΔL116), which were likely associated with disease occurrence and progression. 39 –41 Both mutations result in DNAJC5 protein misfolding and aggregation. To evaluate the feasibility of engineering nuclease-mediated gene therapy for ANCL, a transgenic zebrafish line was constructed with ectopic expression of mutant DNAJC5 in the neurons. As expected, massive protein aggregates were found in the neurons of transgenic zebrafish, which phenocopied ANCL patients. Next, a series of engineering nucleases were designed to target the DNAJC5 gene and to eliminate these protein aggregates. These engineering nucleases, including TALEN and Cas9, did function effectively and introduced indels in their target sites. These indels are supposed to induce a frameshift mutation, leading to DNAJC5 protein truncation, which might eliminate the wrong folding and increase the solubility of this protein. Unfortunately, none of these engineering nucleases made mutant DNAJC5 protein soluble. An attempt was also made to correct the mutation by providing a wild-type DNAJC5 template when TALENs were injected that were specific to the mutant DNAJC5 gene and not supposed to cut the wild-type one. However, co-injection of the wild-type DNAJC5 templates and TALEN mRNAs did not produce any detectable neurons that expressed soluble DNAJC5, which might be the result of the low activity of HDR in higher vertebrates.
Among these engineering nucleases, it was noticed that a sgRNA targeting the 5′ coding region of mDNAJC5, sgRNA5, greatly inhibited mDNAJC5 expression rather than made it soluble. Next, it was shown that the mDNAJC5 mRNA level was significantly reduced in sgRNA5 injected embryos, which could be the result of nonsense-mediated mRNA decay. In agreement with this, a transgenic vector mCherry-mmDNAJC5, harboring a 1 bp deletion within the sgRNA5 target site, also showed much less mCherry expression than mCherry-mDNAJC5 did, though they had the same promoter and were injected with the same dose. Furthermore, the offspring of sgRNA5-treated zebrafish, which had frameshift indels, had fewer levels of mCherry-mDNAJC5 mRNA and protein expression. However, embryos injected with DNAJC5 TALENs and sgRNA3 harbored frameshift mutations, but none of them had an obvious hint of nonsense-mediated mRNA decay, as evidenced by mCherry expression, suggesting that the position of premature stop codon is critical for nonsense-mediated mRNA decay. Together, these results suggest targeting the 5′ coding region is practicable in designing gene therapy against the DNAJC5 mutation. Recently, adeno-associated virus (AAV)-mediated delivery of Cas9 has achieved great success in mouse models. 42 Considering that several AAV subtypes have high tropisms for neural tissues, it will be promising to use AAV-carried Cas9 to remove DNAJC5 mutant genes from neurons. 43 Though targeting the 5′ coding region of DNAJC5 disrupted both the wild-type and mutant DNAJC5 gene, this can be compensated by introducing Cas9-resistant wild-type variant. Therefore, this study is the first step toward ANCL gene therapy utilizing the CRISPR/Cas9 system, and it is believed that such gene therapies will be clinically transformed in the near future.
Footnotes
Acknowledgments
This work was supported by the Hi-Tech Research and Development (863) Program of China 2015AA020309 and the National Natural Science Foundation of China (No. 81372143, No. 31000646, and No. 81123003).
Author Disclosure
We declare that we have no conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.