
Abstract
The Sleeping Beauty transposon system has been extensively tested for integration of reporter and therapeutic genes in vitro and in vivo in mice. Dogs were used as a large animal model for human therapy and minimally invasive infusion of DNA solutions. DNA solutions were delivered into the entire liver or the left side of the liver using balloon catheters for temporary occlusion of venous outflow. A peak intravascular pressure between 80 and 140 mmHg supported sufficient DNA delivery in dog liver for detection of secretable reporter proteins. Secretable reporters allowed monitoring of the time course of gene products detectable in the circulation postinfusion. Canine secreted alkaline phosphatase reporter protein levels were measured in plasma, with expression detectable for up to 6 weeks, while expression of canine erythropoietin was detectable for 7–10 days. All animals exhibited a transient increase in blood transaminases that normalized within 10 days; otherwise the treated animals were clinically normal. These results demonstrate the utility of a secreted reporter protein for real-time monitoring of gene expression in the liver in a large animal model but highlight the need for improved delivery in target tissues to support integration and long-term expression of Sleeping Beauty transposons.
Introduction
N
Hydrodynamic DNA delivery is a promising approach for nonviral gene transfer to the liver in large animals. However, there are significant challenges to adapt hydrodynamic delivery to large animal models, such as development of a minimally invasive, liver-directed technique. We focused on dogs because existing canine models of several genetic diseases could be used to model human genetic disease. Previously, hydrodynamic delivery of plasmids in dogs has been achieved by a surgical clamping technique to occlude the liver prior to IVC or bile duct infusion, 10 or by occlusion of the hepatic vein of the target lobe. 11 Application to some diseases (e.g., hemophilia) will require a minimally invasive procedure. Accordingly, we developed a minimally invasive system that allows for both occlusion of the entire liver as well as infusion of DNA into the left hepatic vein. Balloon catheters have also been used for hepatic occlusion with pressurized delivery of plasmid DNA in rabbits 12 and in pigs. 13 –16 These previous studies demonstrated the extended applicability of hydrodynamic delivery from rodents to large animals using short-term reporter (luciferase) gene expression.
To achieve sustained gene expression after hydrodynamic delivery of plasmids, we employ the Sleeping Beauty (SB) transposon system, 17 which consists of a transposon comprised of inverted terminal repeat sequences that flank the genetic cargo plus an SB transposase enzyme. 18,19 SB transposase excises the engineered SB transposon from its plasmid donor and inserts it into the target genome. In mice, hydrodynamically delivered SB transposons have been used successfully to correct murine models of inherited genetic diseases, including those for hemophilia A, 20 hemophilia B, 21,22 tyrosinemia type 1, 23,24 and mucopolysaccharidosis types 1 and 7. 25,26
Evaluation of the effectiveness of the SB system to mediate stable chromosomal integration requires the ability to track transgene expression over an extended period of time by sampling the blood of individual animals. Accordingly, we isolated and characterized the coding sequences for canine secretable reporters, canine erythropoietin (cEPO) and canine secreted alkaline phosphatase (cSEAP), 27 which allowed us to monitor longitudinal transgene expression and secretion into the blood. In addition, since canine EPO and SEAP are canine proteins, we expected them to be less immunogenic than other reporter proteins such as either luciferase 28 or green fluorescent protein (GFP). 29 Our results demonstrate that secreted reporters can be used to monitor the effectiveness of hydrodynamic delivery of plasmids to the liver in large animals.
Materials and Methods
Plasmids
The canine erythropoietin (EPO) cDNA sequence was amplified by PCR from dog cDNA and the 5′-sequence engineered to encode authentic canine EPO. A canine heat-stable alkaline phosphatase coding sequence was assembled from a combination of cDNA, genomic, and synthetic sequences. Truncation at codon 521 was predicted using a glycosylphosphatidylinositol prediction algorithm 30 to interrupt cellular retention, allowing the polypeptide to be secreted. Plasmid pCMV-SB100X 31 was a kind gift from Dr. Zoltan Ivics (Paul Ehrlich-Institut, Langen, Germany).
pKT2/mCAGGS-cEPO//Ub-SB11
A 1780 bp PvuII fragment containing the human ubiquitin promoter sequence and the SB11 coding sequence was isolated from pUb-SB11 32 and ligated into the SwaI site of pKT2/mCAGGS (created from assembly of pKT2/CaL 33 ) to form pKT2/mCAGGS//UbSB11. The 688bp cEPO coding sequence was then isolated from TOPO4.0-cEPO as an EcoRI/AscI fragment, and cloned between EcoRI and AscI sites downstream of mCAGGS in pKT2/mCAGGS//UbSB11 to generate pKT2/mCAGGS-cEPO//Ub-SB11 (Fig. 1).
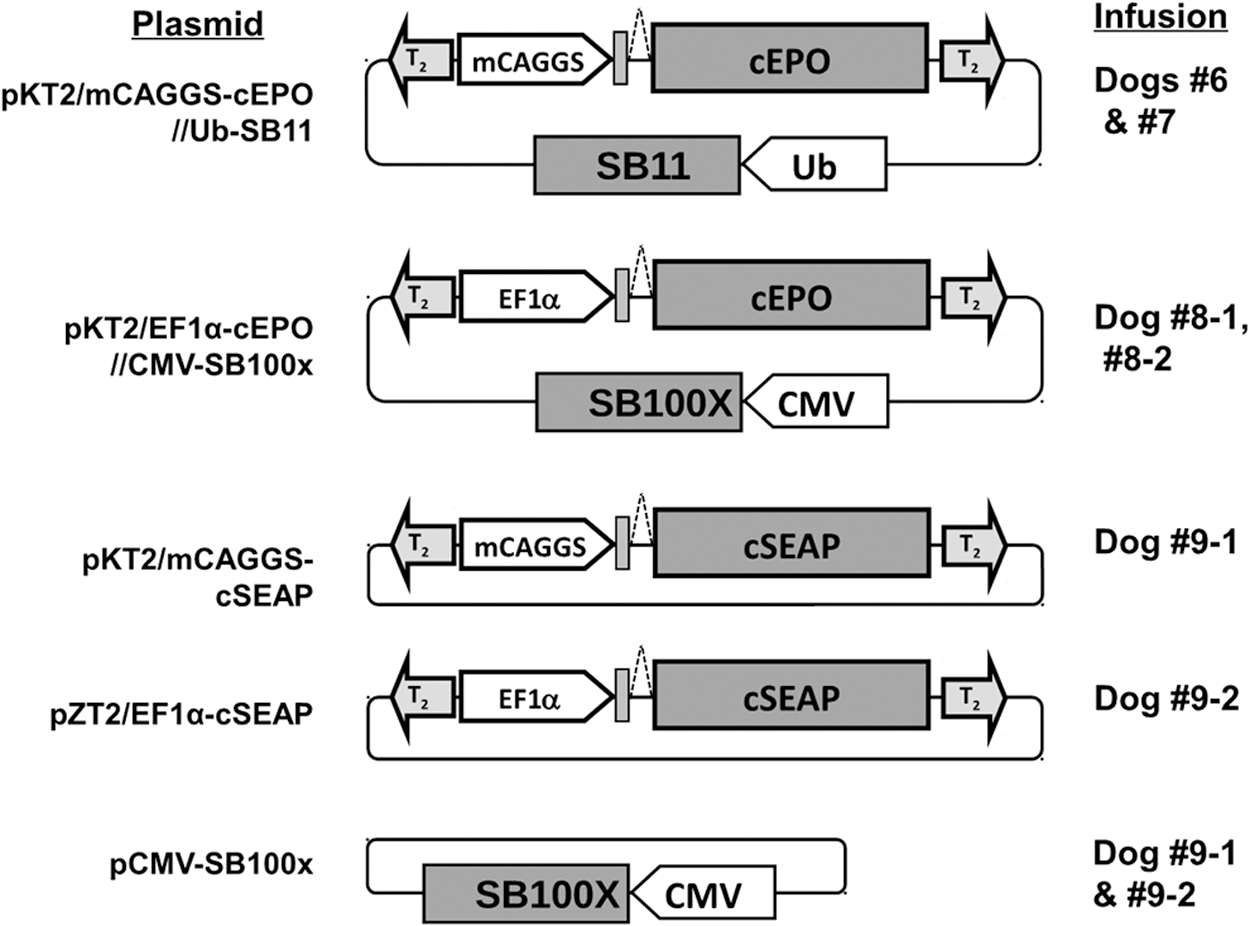
Transposon- and transposase-encoding plasmids. Transposons contain T2 inverted-terminal repeats (shaded arrows) and a reporter gene sequence (shaded boxes) encoding either canine erythropoietin (cEPO) or canine secreted alkaline phosphatase (cSEAP). Reporter gene expression was regulated either by the miniCAGGS (mCAGGS) promoter or by a CpG-less variant of elongation factor 1 alpha (EF1α) (unshaded block arrows). pKT2/mCAGGS-cEPO//Ub-SB11 and pKT2/EF1α-cEPO//CMV-SB100X also contain a ubiquitin promoter–regulated SB11 gene or a cytomegalovirus (CMV) promoter-regulated SB100X gene, respectively, exterior to the cEPO-encoding transposon. For the other transposons, Sleeping Beauty (SB) transposase was provided in trans by co-infusion of a CMV promoter-regulated SB100X expression plasmid (pCMV-SB100X).
pKT2/EF1α-cEPO//CMV-SB100X
An 824 bp EcoRI/EagI fragment containing the cytomegalovirus (CMV) promoter/enhancer sequence was isolated from pCMV-SB11 34 and ligated into pKT2/mCAGGS-cEPO//Ub-SB11 in place of the ubiquitin promoter to generate pKT2/mCAGGS-cEPO//CMV-SB11. A modified CpG-less elongation factor 1 alpha (EF1α) promoter sequence (CLP, InvivoGen, San Diego, CA) was isolated from pKT2/EF1α-GFP (similar to pKT2/EF1α(CLP)-Luc 33 ) as an NcoI/BglII fragment and cloned between NcoI and BglII in place of the CAGGS promoter of pKT2/CAGS-cEPO//CMV-SB11 to generate pKT2/EF1α-cEPO//CMV-SB11. Finally, a 3318 bp SacI/PvuI fragment containing the SB100X coding sequence and the rabbit β-globin polyadenylation signal was isolated from pCMV-SB100X 31 and ligated into pKT2/EF1α-cEPO//CMV-SB11 between SacI and PvuI sites downstream of the CMV promoter to generate pKT2/EF1α-cEPO//CMV-SB100X.
pKT2/mCAGGS-cSEAP
The truncated cSEAP coding region was cloned into pKT2/mCAGGs 20 between EcoRI and AscI downstream of the mCAGGS promoter to generate pKT2/mCAGGS-cSEAP.
pZT2/EF1α-cSEAP
pZT2/MCS (provided by Dr. Andrew Wilber, Southern Illinois University School of Medicine, Springfield, IL) was generated by ligation of a 671 bp BamHI (blunt)-KpnI (blunt) fragment (consisting of multiple restriction sites flanked by T2 IR/DRs) excised from pKT2/SE 35 with a 730 bp PacI fragment of pCpG-MCS (InvivoGen, San Diego, CA) containing CpG-free plasmid sequences. The cSEAP coding sequence was excised from pKT2/mCAGGS-cSEAP as a 1627 bp EcoRI fragment and inserted into pKT2/EF1α-SV40pa downstream of the EF1α sequence. EF1α and cSEAP sequences were then excised as a 990 bp BamHI-HindIII fragment and an 1818 bp HindIII-SalI fragment, respectively, and ligated into pZT2/MCS between BamHI and XhoI to generate pZT2/EF1α-cSEAP. Endotoxin-free plasmid DNA was either prepared using the Endofree Plasmid Maxiprep kit from Qiagen or in quantities larger than 10 mg by Aldevron Corp (Fargo, ND).
Hydrodynamic plasmid infusions in dogs
All procedures and animal care were approved by the University of Minnesota's Institutional Animal Care and Use Committee with the Animal Welfare Assurance No. A3456-01 and following the Guide for the Care and Use of Laboratory Animals. Female (dog 6) and male (all others) beagle dogs were procured from a US Department of Agriculture class-A vendor for the described procedures. Test animals were fasted for 12 h prior to anesthesia, sedated with acepromazine 0.4 mg/kg subcutaneous (SQ) and atropine 0.05 mg/kg SQ or torbugesic at 0.05–0.5 mg/kg SQ, and then anesthesia-induced with propofol at about 4 mg/kg intravenously (i.v.) followed by intubation and general anesthesia using isoflurane 1–2% with 3–4 L/min of oxygen at approximately 15 cc/kg tidal volume. Animals were continuously monitored by visual assessment, ECG, and intravascular venous pressure recording. A warming water blanket and an air blanket were used to maintain the animal's body temperature, and lactated Ringer's solution or 0.9% NaCl was given at a rate of 5–10 mL/kg/hour during anesthesia. With the animal under general anesthesia and using sterile technique, a cut down to the femoral vein and/or an external jugular vein was performed, and an appropriately sized introducer sheath was placed into the vessel using Seldinger technique.
Three different strategies (three catheter, double balloon, and single balloon) were used for balloon-mediated occlusion of the liver with subsequent infusion of DNA into the hepatic venous circulation (Fig. 2). Three catheter system: (i) A 6 Fr single-balloon wedge pressure or Berman angiographic balloon catheter (Arrow No. AI-07126 or AI-07131) was introduced through the right femoral vein to the IVC immediately below the liver; (ii) a second 6 Fr single-balloon catheter was introduced through the jugular vein to the IVC immediately above the liver; and (iii) an 8 Fr high-flow Cordis pigtail catheter (No. 801-618) or a 4 Fr Vanguard Dx Medrad pigtail (No. 155, 1200 PSI) was introduced through the left femoral vein past the lower balloon-catheter into the region of the hepatic veins in the IVC. Double balloon system: Custom-designed double-balloon catheters were assembled by Innovations in Medicine (Inver Grove Heights, MN (or by Minnesota MedTec, Inc., (Minneapolis, MN). Dog 7 was infused using a 5 Fr catheter containing appropriately positioned compliant balloons and multiple ports spaced between the balloons for delivery of DNA solution at 20–40 mL/s, 300–600 pounds per square inch (PSI). Dog 8 (both infusions) and dog 9 (infusion 1) were infused using a similarly designed 9 Fr catheter that included a third lumen. The catheters were introduced through the right femoral vein into the IVC and a second pressure-sensing catheter was introduced through the left femoral vein with the end of the catheter positioned between the two balloons. Single balloon system: dog 9 (infusion 2) was infused using a custom-designed 8Fr catheter containing a single compliant balloon and multiple ports positioned distal to the balloon for delivery of DNA solution at up to 20 mL/s, 333 PSI. This catheter was introduced through the right femoral vein to the IVC and positioned just inside the left hepatic vein (LHV). A second pressure-sensing catheter was introduced alongside the balloon catheter from the right femoral vein with the end of the catheter positioned distal to the balloon.
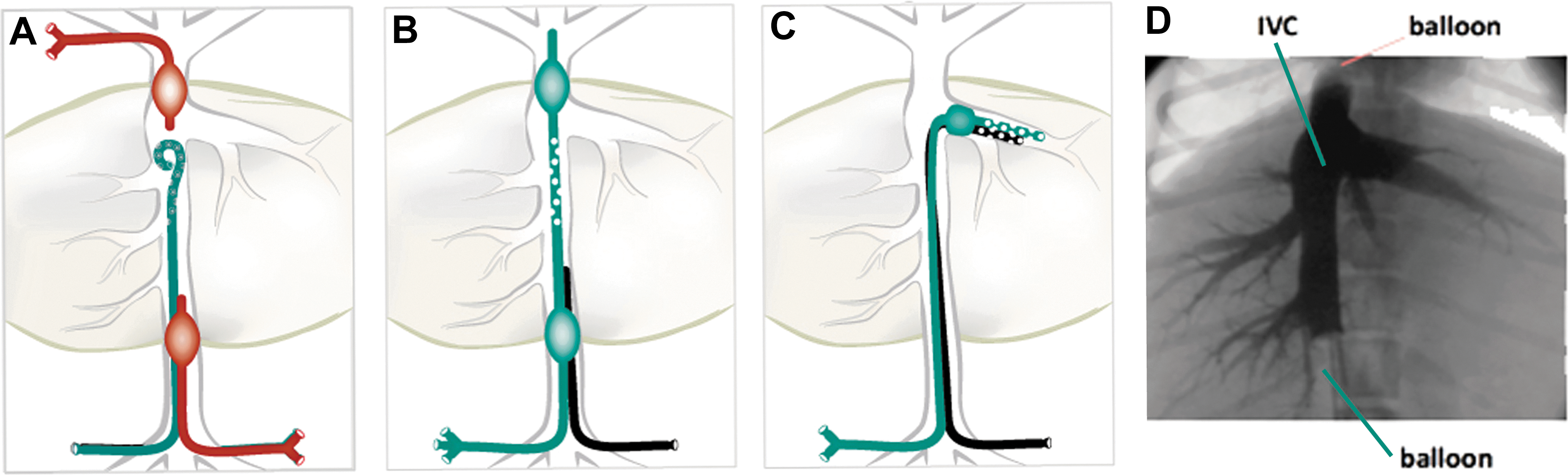
Strategies for catheter-mediated DNA delivery to dog liver.
Fluoroscopy was used to position the catheters, and then the balloons were briefly inflated and a small amount of contrast dye was infused by hand to verify correct catheter placement, occlusion of the IVC, and access to the hepatic venous circulation (Fig. 2D; see Supplementary Video S1 for video recording of infusion; Supplementary Data are available online at
At the end of each experiment, animals were anesthetized with propofol at 2–6 mg/kg i.v. and given heparin at 150 units/kg i.v. Approximately 15 min later, the animals were euthanized with 1 mL Euthasol (390 mg pentobarbital +50 mg phenytoin) per 4.5 kg. The livers were perfused with 0.5–1 L sterile saline to remove residual blood. A gross general necropsy was performed and samples of liver and other organ tissues were collected for molecular analyses.
cEPO and cSEAP activity assays
Plasma samples from animals infused with cEPO-encoding plasmids were assayed by ELISA for canine EPO using the Quantikine IVD Human Erythropoietin kit from Bio-Techne (Minneapolis, MN). To determine assay suitability, human EPO standard was assayed in parallel with purified recombinant canine EPO (Bio-Techne, Minneapolis, MN). Canine EPO amounts are reported in milli-International Units per mL (mIU/mL). Plasma samples from animals infused with cSEAP-encoding plasmids were heated for 10 min. at 65°C and then assayed for alkaline phosphatase activity using Tropix Phospha phospholuminescent substrate (Applied Biosytems, ThermoFisher) with human SEAP as a standard for calculation of values in ng/mL of plasma.
Quantitative real-time PCR assay for canine SEAP sequences
Approximately 30 liver samples were pulverized in liquid nitrogen, and then the DNA was isolated by phenol-chloroform extraction. DNA was extracted from dog plasma (50 μL) using the ZR-96 Quick-gDNA kit (Zymo Research, Irivine, CA) and eluted with equal volume 1 × TE buffer. PCR reactions (20 μL) were prepared from ABGene Absolute Blue Probe master mix (Fisher Scientific); 600 nM each primer: forward (5′-TGGTCCTCTGCCCTCAACA-3′) and reverse (5′-CCTCCTCAGCTGGGATGATGC-3′); 200 nM Universal Probe 3 (5′-FAM labeled; Roche Applied Science, Indianapolis, IN); and 100 ng dog liver DNA or 2 μL DNA extracted from plasma. PCR cycling conditions: 95°C for 15 min followed by 45 cycles of 95°C for 15 s and 60°C for 15 s, using a LightCycler 480 (Roche). Samples were quantitated by comparison to pKT2/-EF1α-cSEAP3II//PGK-SB100X plasmid, diluted serially in dog genomic DNA (Zyagen, San Diego, CA) in 10 mM TRIS, 0.1 mM EDTA, pH 8.0 (TE). The assay lower limit was defined as the amount of DNA in the standard curve that scored positive 95% of the time, and was equivalent to 2 × 10−4 copies per diploid genome equivalent in dog liver and 2 × 103 copies per mL plasma. Negative controls were included in each assay: TE and canine genomic DNA purchased from Zyagen. The negative controls always had a threshold cycle value less than the assay lower limit.
Results
Secreted canine reporter protein EPO
A key factor in the strategy of these studies was the ability to monitor transgene expression by periodic blood sampling. For this purpose, a transposon was assembled encoding the canine EPO gene regulated by the mini CAGGS (mCAGGS) promoter 20 and containing an ubiquitin promoter-regulated SB11 sequence in cis (Fig. 1). All transposon preparations were hydrodynamically infused into mice to assure the integrity and transgene expression potential of the constructs. Control groups of mice were infused with pKT2/mCAGGs-murine EPO 36 or pKT2/mCAGGS-cEPO plasmid lacking the SB11 sequence. Expression of murine or canine EPO in mice was detected for up to 5 weeks, which elicited an extended increase in hematocrit, two times above normal levels for 5 months (Supplementary Fig. S1).
Hepatic venous delivery of plasmid DNA
Although a 1.5- to 2-fold transient increase in liver volume is optimal for hydrodynamic delivery in mice, 8 we were limited to an infusion volume of 200 mL, the maximum amount delivered using the MEDRAD ProVis power injector. Therefore, 5–6 kg dogs were procured in order to approximate a 1:1 ratio of the DNA infusion volume to liver volume, estimated at 3% total body weight. 37 Experiments were performed in the first five dogs to evaluate conditions for catheter placement and plasmid DNA infusion into the liver. Effective conditions were identified in dog 6, with the use of an off-the-shelf, three-catheter system (Fig. 2A, Table 1) to infuse a cEPO-encoding plasmid (pKT2/CAGS-cEPO//Ub-SB11; Fig. 1). Supplementary Video S1 shows both vascular access and hydrodynamic infusion. cEPO was detectable in the peripheral blood until 4 days postinfusion, with expression peaking one day post-infusion (Fig. 3A). However, the delivery procedure did not result in extended transgene expression sufficient for a therapeutic effect or for long-term monitoring.
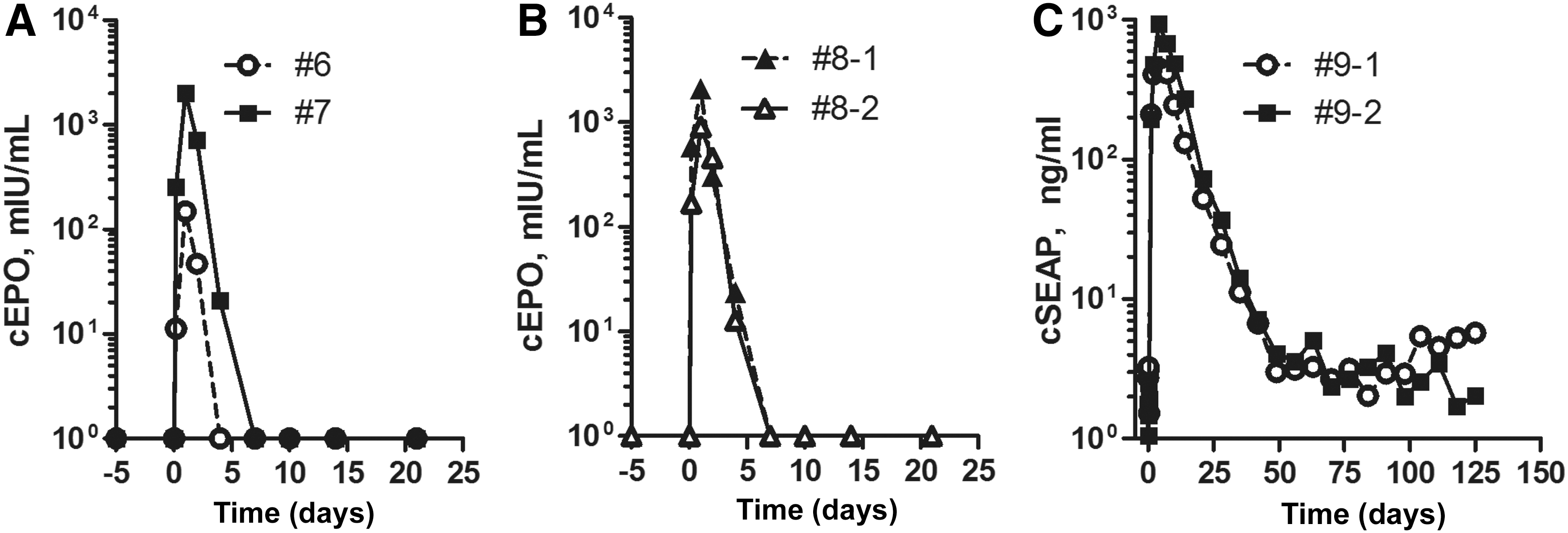
Secretable reporter expression after hydrodynamic DNA infusion in dogs. Plasma samples were collected before, during, and after DNA infusion and were assayed for canine erythropoietin (cEPO) by ELISA
Summary of balloon catheter-mediated DNA delivery to liver of dogs
Infusion route: venous infusion to the whole liver via inferior vena cava (IVC); venous infusion into the left half of the liver through the left hepatic vein (LHV).
pKT2/mCAGGS-cEPO//Ub-SB11.
pKT2/EF1alpha-cEPO//CMV-SB100X.
pKT2/CAGGS-cSEAP +2.2 mg pCMV-SB100X.
pZT2/EF1alpha-cSEAP +2.0 mg pCMV-SB100X.
ALT, alanine aminotransferase; AST, aspartate aminotransferase; DNA, deoxyribonucleic acid; EPO, erythropoietin; ID, identification; mmHg, millimeters of mercury; NM, not measured; SEAP, secreted alkaline phosphatase; Tn, transposon.
Double-balloon catheter for hepatic venous DNA delivery
To simplify subsequent infusions, we designed a double-balloon catheter for occlusion of the IVC and infusion of DNA solution from the same catheter between the two balloons (Fig. 2B). This double-balloon catheter system provided effective occlusion as shown by fluoroscopy (Fig. 2D). In dog 7, IVC intravascular pressure was slightly elevated at about 2–5 mmHg upon inflation of the balloons, increased dramatically upon initiation of the DNA infusion (Supplementary Fig. S2), decreased after the infusion, and dropped to normal upon deflation of the balloons. A computed tomography (CT) scan of the liver conducted 1 hour after the infusion was normal. Canine EPO expression in dog 7 followed a pattern similar to dog 6, peaking 1 day postinfusion, with expression detectable until 7 days postinfusion (Fig. 3A). No change in hematocrit was observed (Supplementary Fig. S3).
Subsequently, we conducted two infusions to test whether silencing of the mCAGGS promoter caused loss of transgenic cEPO expression. We first infused a transposon encoding a variant EF1α promoter with enhancer sequences lacking CpG sites that could be targets for methylation and subsequent down-regulation (dog 8-1 with triluminal catheter, Fig. 1B). We additionally hypothesized that blockade of inflammation prior to and post infusion would improve the duration of expression. The same animal was infused six weeks later (dog 8-2) with the same plasmid (pKT2/EF1α-cEPO//CMV-SB100X) at a reduced dose (2 mg/kg), and with administration of 1 mg/kg dexamethasone for 5 days, starting 1 day prior to delivery of the DNA. For both infusions of dog 8, the peak level of cEPO was observed 1 day postinfusion and cEPO was rapidly extinguished by day 10–14 (Table 1, Fig. 3B).
The double-balloon catheter thus provided a simplified surgical intervention for infusion of DNA. However, expression of the cEPO transgene was transient after all infusions, despite the use of two different transcriptional regulatory sequences. A reduction in the DNA dose by nearly 80% resulted in only a 50% decrease in cEPO gene expression, indicating that the plasmid dose of ∼2 mg/kg was close to saturating.
Canine SEAP
As expression of cEPO was transient, we created an alternate reporter, a heat-stable cSEAP gene from a combination of intestinal alkaline phosphatase cDNA and synthetic sequences derived from the dog genome. The cSEAP sequence contained 11 silent mutations at the 5′-end that permitted selective amplification by PCR when mixed with the endogenous sequence in genomic DNA. A mCAGGS-regulated cSEAP transposon was assembled (pKT2/mCAGGS-SEAP, Fig. 1), and first infused into mice, where it supported expression of ∼4.0 × 104 ng/mL heat-stable SEAP activity in the plasma one day after hydrodynamic injection. Dog 9 was infused with a 5:1 ratio of pKT2/mCAGGS-cSEAP (2 mg/kg): pCMV-SB100X 31 (0.4 mg/kg; Fig. 1) using the double-balloon catheter system (Fig. 2B). The dose of transposase gene was reduced because SB100X has increased transposase activity compared to SB1131 and because there is reduced transposition efficacy at 1:1 ratio of transposon to SB100X plasmids, likely due to overexpression inhibition (our unpublished observations). Heat-stable cSEAP activity in the peripheral blood peaked on day 4 postinfusion and was clearly detectable through 6 weeks (42 days) postinfusion (Fig. 3C). The half-life of transgenic cSEAP was approximately 4.5 days compared with about 1 day for cEPO.
A second infusion was carried out in dog 9, which had grown over the intervening five months to 9.2 kg. The transposon pZT2/EF1α-cSEAP (Fig. 1) contained the cSEAP gene in a CpG-less plasmid backbone, including the EF1α promoter. In order to maintain approximately a 1:1 infusion volume to target tissue ratio, a single-balloon catheter was used to occlude the major LHV for DNA infusion limited to the left side of the liver (Fig. 2C). A 6:1 ratio of plasmids pZT2/EF1α-cSEAP:pCMV-SB100X was infused into the LHV through infusion ports distal to the balloon (Fig. 2C, Table 1). The animal's liver was evaluated by ultrasound immediately after infusion, and by CT scan 2 days after infusion; neither test indicated significant injury to the liver. After the infusion, heat-stable cSEAP activity was detected with an identical time course to the first infusion using pKT2/mCAGGS-cSEAP (Fig. 3C), suggesting that the CpG-less promoter did not prevent the ultimate reduction of cSEAP reporter gene expression.
Impact of hydrodynamic infusion on serum biochemistry and hematology
In all dogs, there was an increase in circulating liver enzymes, which returned to normal after 2 days (aspartate transaminase [AST]) or by 14 days (alanine transaminase [ALT]) (Fig. 4). In all infusions, dogs exhibited a transient increase in glucose, creatine kinase, and lactate dehydrogenase levels that returned to normal 1 day later (Supplementary Fig. S3). Levels of creatinine, albumin, hematocrit, and hemoglobin all exhibited a decrease postinfusion that rapidly returned to normal levels (Supplementary Fig. S3). This could be influenced by the volume and number of blood samples obtained in a short interval. In dogs infused with cEPO-encoding plasmid, only one in four infusions resulted in alkaline phosphatase levels outside the normal range (Supplementary Fig. S3). Gamma-glutamyl transpeptidase levels were not altered outside of the normal range (Supplementary Fig. S3), indicating that bile flow was not negatively impacted by DNA infusion. There were small fluctuations outside of the normal range in numbers of circulating levels of monocytes, red blood cells and mature neutrophils after DNA infusion, which returned to normal by around 5 days postinjection (Supplementary Fig. S4). Platelet counts were decreased immediately after infusion and returned to normal by 30 days postinjection, while lymphocyte counts remained elevated in 2 of 6 infusions at 30 days (Supplementary Fig. S4).
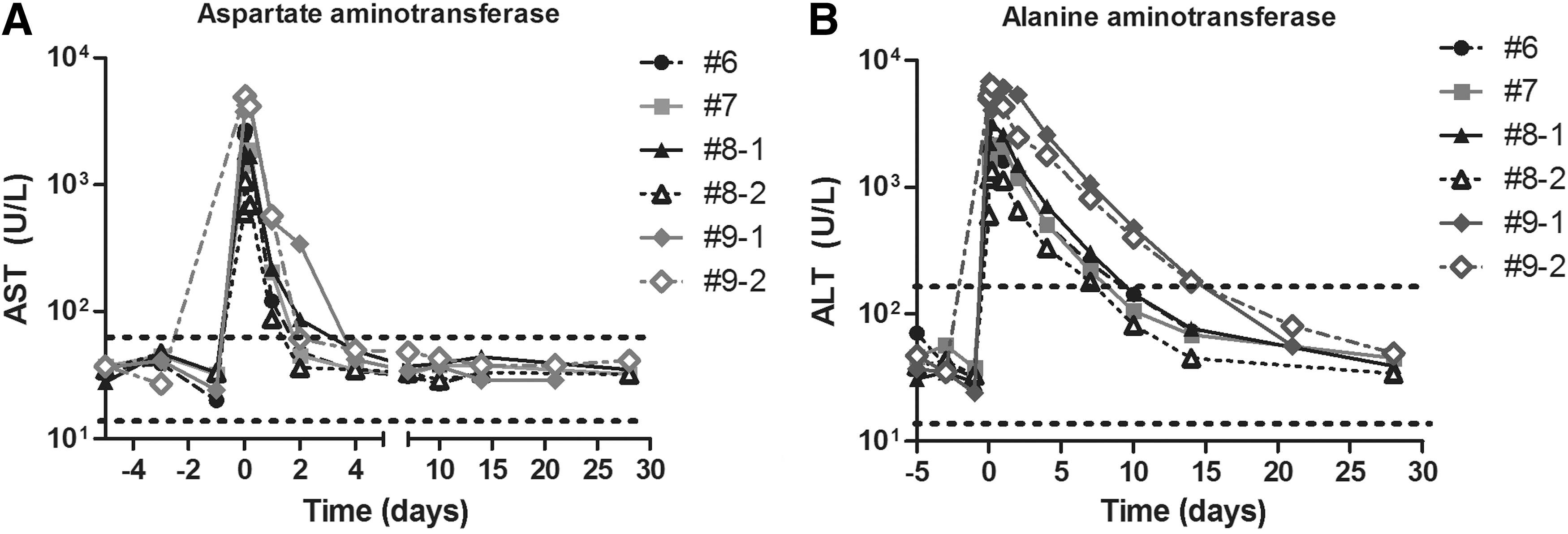
Serum transaminase levels after hydrodynamic infusion. Serum samples collected before, during, and after hydrodynamic infusion were assayed for
Assessment of cSEAP-encoding plasmid in the plasma collected after both infusions showed a peak of about 1011 copies/mL in plasma immediately postinfusion that was rapidly cleared from the circulation, reaching background levels by 4 days (Fig. 5A). Within 5 hours, there was about a 1000-fold reduction in the number of plasmids in the plasma. Sequences encoding cSEAP were detectable in the left lateral lobe of dog 9 at 4.6 months after the second infusion (Fig. 5B). The rapid clearance of plasmid from the circulation was similar to observations in mice, in which hydrodynamically injected plasmid DNA (0.5 mg/kg) was cleared at 30 min postinfusion. 38 The results from these experiments thus confirm the value of cSEAP as a reporter and demonstrate the effectiveness of hydrodynamic delivery to either the entire liver or to the left side of the liver through the left hepatic vein.
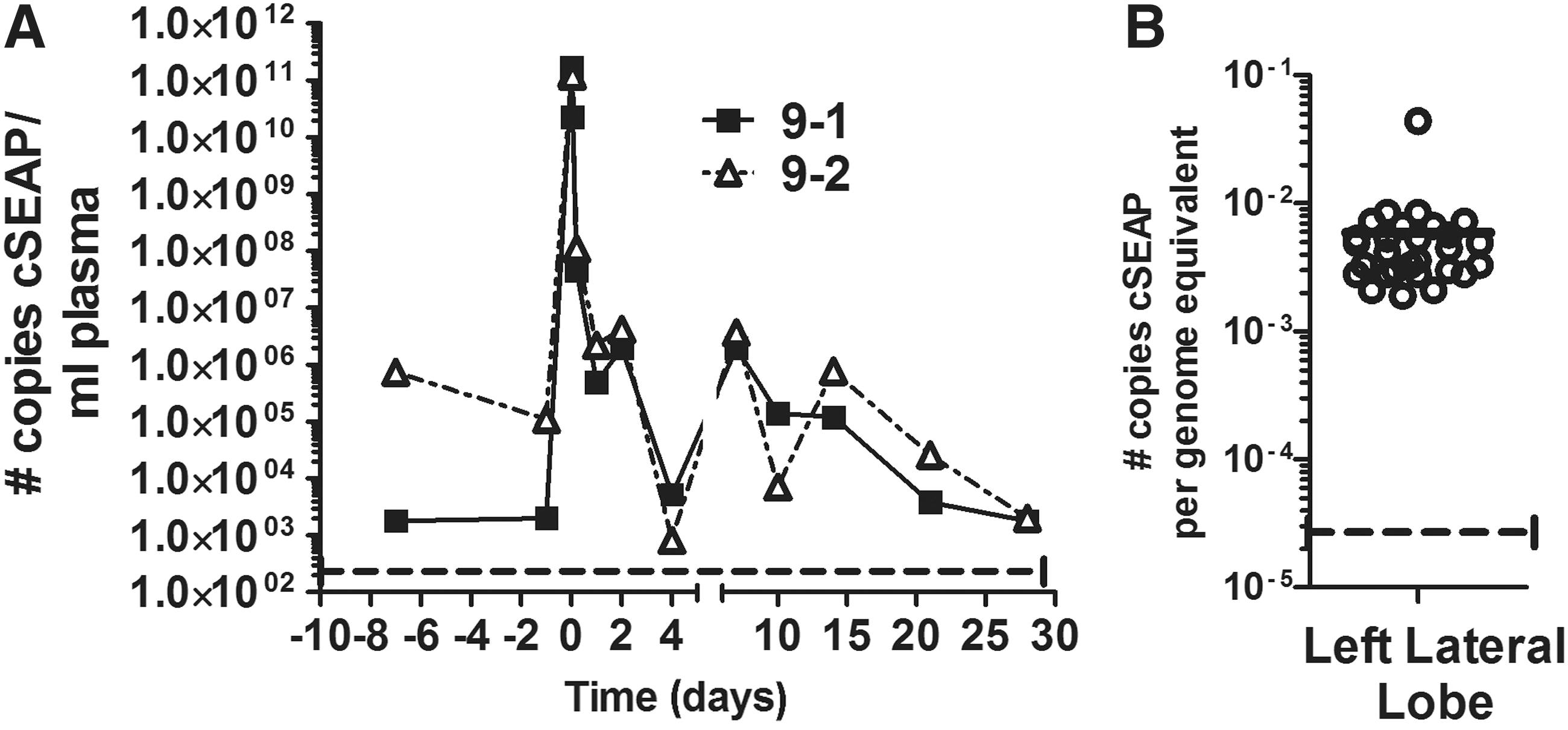
Clearance of infused plasmid from circulation and detection in the liver.
Discussion
We have developed novel secreted reporter proteins to track long term gene expression in large animals. Of the two secreted reporter proteins, expression of cSEAP by canine liver was detected for a longer duration than cEPO expression. In mice, cEPO levels dropped from the peak level of expression one week after infusion of cEPO transposons, followed by stabilization of expression (Supplementary Fig. S1A). However, cEPO expression in dogs reached about 1000-fold above background, in contrast to 104 to 105 units above background in mice. Hence, our data are insufficient to distinguish whether we were unable to detect long-term expression due to reduced efficiency of delivery or reduced stability of expression. However, the extremely short duration of detectable cEPO expression, 1 week or less, explains the lack of alteration in hematocrit for dogs infused with cEPO-encoding transposons, since an increase in hematocrit required at least 2 weeks of canine EPO injections. 39 The time course of reporter gene product detected in the plasma was remarkably similar after each of four cEPO plasmid infusions, regardless of the promoter regulating reporter gene expression, the activity level of SB, co-delivery of the SB gene in cis or in trans, or the overall dose of plasmid DNA infused (2 mg/kg vs.10 mg/kg). It is unlikely that the extinction of cEPO or cSEAP was the result of adaptive immunity because identical expression profiles were obtained after a second infusion of cEPO transposon in dog 8 (Fig. 3A, B) and of cSEAP transposon in dog 9 (Fig. 3C).
We used a minimally invasive system with catheter-mediated occlusion of the IVC for delivery of plasmid DNA to whole liver or temporary occlusion of the hepatic vein for delivery to the left side of the liver in dogs. We achieved gene expression after DNA infusion without blocking outflow through the portal vein or hepatic artery, which was previously reported using balloons or clamping in pigs. 13,15,40 Blocking outflow of DNA solution via the portal vein may not be necessary because we and others demonstrate the effectiveness of hydrodynamic infusion without such blockade in dogs, 11 pigs, 14,16 and rabbits. 12 The hydrodynamic procedure was well tolerated by the animals; by 6–24 h post anesthesia and procedure, the dogs were eating, drinking, and exhibiting normal activity. No long-term adverse effects were observed.
Effective hydrodynamic delivery depends on increased intrasinusoidal pressure for extravasation of DNA-containing solution across the sinusoidal endothelial lining and entry into hepatocytes. 41,42 There have been varying reports on the relationship between intravascular pressure and effective DNA delivery; 43 we found that a peak intravascular pressure between 80 and 140 mmHg supported sufficient DNA delivery in dog liver for detection of circulating reporter proteins. Most infusions have been conducted at a constant infusion rate with varying pressure. Liu et al. use a computer-assisted infusion technique that maintains constant pressure during hydrodynamic injection. 11,15,16
The intravascular pressure achieved during DNA infusion results in permeabilization of hepatocyte membranes and leakage of liver transaminases AST and ALT into the circulation. 7,44 AST has a shorter half-life than ALT, which explains why it takes longer for circulating levels of ALT to return to normal. 45 While elevations in circulating AST, ALT, and lactate dehydrogenase are indicators of chronic liver disease, in our experiments increased levels of the enzymes were transient and returned to normal after 2–10 days.
In conclusion, we have engineered a reporter protein, canine secreted alkaline phosphatase, which can be measured by periodic blood sampling. In all dogs, we found an increase in circulating levels of innate immune cells, neutrophils and monocytes, along with an increase in lymphocytes, cells of the adaptive immune system. Neutrophils are released from bone marrow reserves in response to increased inflammatory cytokines and other mediators during tissue inflammation or damage. 46 Dexamethasone is a glucocorticoid that blocks expression of pro-inflammatory cytokines. Dog 8, infused with a cEPO-encoding transposon plasmid, was given dexamethasone before and during the second DNA infusion. However, there were no changes in the kinetics of circulating EPO expression and no difference in the numbers of circulating monocytes and neutrophils between the first and second infusions of dog 8. Resident liver macrophages (Kupffer cells) are exposed to DNA immediately after infusion, since they line the walls of the sinusoids, and activated Kupffer cells secrete inflammatory cytokines and other mediators during an innate immune response. 47 Canine Kupffer cells are more reactive than Kupffer cells from rat or monkey, and secrete greater levels of cytokines interleukin-1β and interleukin-6 in response to lipopolysaccharide. 48 Depletion of Kupffer cells does not negatively impact DNA delivery or gene expression after hydrodynamic delivery in mice. 38 Future experiments will determine whether a drug or combination of drugs that block Kupffer cell activity and/or prevents the release of circulating immune cells is effective at achieving a longer duration of expression.
Footnotes
Acknowledgments
We thank Greg Brucker (Innovations in Medicine) and Brian Packard (Minnesota MedTec, Inc.) for catheter assembly and Dr. Steven Kass (University of Minnesota) for valuable discussions, Julie Arnold (Bio-Techne) for artwork, and the staff of the Department of Radiology (University of Minnesota) for assistance with infusions during the early phase of this work. Infusions were carried out in the Kurt Amplatz Interventional Laboratory, Department of Radiology, University of Minnesota, Minneapolis, MN.
Author Disclosure
This work was supported by US Public Health Service grants R44 HL072539 (R.S.M.) and R41 DK081249 (P.B.H.) to Discovery Genomics, Inc., and grant R01 DK082516 to the University of Minnesota (P.B.H.) from the National Institutes of Health. P.B.H., R.S.M., K.A.H., and E.R.O have equity in Immusoft, Inc. (formerly Discovery Genomics, Inc.). E.R.O. and R.S.M. are employees of Immusoft, Inc. E.L.A, J.B.B, M.U.R., R.G., and D.W.H. have no competing financial interests.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.