
Abstract
Adeno-associated virus (AAV) vectors are one of the most frequently applied gene transfer systems in research and human clinical trials. Since AAV vectors do not possess an integrase activity, application is restricted to terminally differentiated tissues if transgene expression is required long term. To overcome this limitation and to generate AAV vectors that persist episomally in dividing cells, AAV vector genomes were equipped with a scaffold/matrix attachment region (S/MAR). After a mild antibiotic selection, cells transduced with AAV-S/MAR established colonies that maintained long-term transgene expression (>50 population doublings) from replicating AAV vector episomes in the absence of further selection. Unexpectedly, with a lesser but still significant efficiency, the control vector (AAV-ΔS/MAR), a standard single-stranded AAV vector, also established stable transgene-expressing colonies, most of which were maintained as replicating episomes rather than integrated vector genomes. Thus, based on the result in HeLa cells, it is concluded that AAV vector genomes per se possess the ability to establish episomal maintenance in proliferating cells, a feature that can be enhanced by incorporation of a foreign genomic element such as an S/MAR element.
Introduction
A
In non-viral gene delivery, genomic elements, such as Scaffold/matrix attachment regions (S/MARs) are routinely used to establish long-term gene expression from plasmids. In the pioneer S/MAR plasmid vector pEPI, the S/MAR element mediates interaction with the nuclear matrix via matrix protein SAF-A, thus maintaining episomal transgene expression over hundreds of generations following an initial phase of antibiotic selection. 14,15 Furthermore, the general applicability of S/MAR-based episomes for therapeutic gene transfer is well accepted. 16 –20 However, as efficiency of non-viral gene transfer is low, S/MAR elements are combined with the high transduction efficiency of viral vectors. For example, a high-capacity adenoviral vector delivering a S/MAR element containing vector genome mediated long-term transgene expression in vitro (6 weeks) and in vivo (6 months in murine liver). 21 Likewise, non-integrating lentiviral vectors (anchored non-integrating lentivirus [aniLV]) when equipped with S/MAR elements remained stable for >100 cell divisions in the absence of selection. 22
Based on this background, this study assayed an S/MAR element in the context of AAV vectors, aiming to establish a strategy to equip AAV vectors with the ability to modify proliferating cells permanently in the absence of vector genome integration.
Materials and Methods
Cell culture
The human embryonic kidney cell line HEK293 (ATCC CRL-1573) and the human cervix carcinoma cell line HeLa (ATCC CCL2) were cultured in Dulbecco's modified Eagle's medium with GlutaMAX-I (Invitrogen, Karlsruhe, Germany) supplemented with 10% fetal calf serum (Invitrogen), 100 IU/mL of penicillin (Invitrogen), and 100 μg/mL of streptomycin (Invitrogen). For replication experiments, cells were synchronized in G1/S-phase using a double thymidine block. Briefly, cells were grown to 50% confluency, and 2 mmol/L of thymidine was added for 18 h. Subsequently, cells were released in fresh medium for 9 h followed by a second thymidine block (2 mmol/L) for 15 h. 23 Synchronization was verified by quantifying DNA content by flow cytometry. 24
Production of AAV-S/MAR and AAV-ΔS/MAR
The transgene expression cassette of pGFP 25 was exchanged for the hCMV/EF1P-eGFP-IRES-blasticidin resistance gene (BSD)-S/MAR cassette of pEPito-eGFP-IB (kindly provided by R. Haase), 26 thereby generating plasmid pAAV-S/MAR (Fig. 1A). A control vector plasmid, pAAV-ΔS/MAR, lacking the S/MAR sequence was constructed by deleting the S/MAR of pAAV-S/MAR using HpaI (Fig. 1B). AAV2 vectors were produced and purified as described 27 using the AAV helper plasmid pRC, 28 the adenoviral helper plasmid pXX6-80 (kindly provided by J. Samulski), 29 and the vector plasmid pAAV-S/MAR or pAAV-ΔS/MAR. Viral vector genomic particle titers were determined by isolating DNA from vector preparations, followed by quantitative polymerase chain reaction (qPCR) using green fluorescent protein (eGFP)-specific primers. 30 Transducing titers were determined by transduction of HeLa cells with a serial dilution of the vector preparations. The percentage of eGFP-expressing cells was determined 48 h post-transduction by flow cytometry (FACS Canto II; BD Biosciences, Franklin Lakes, NJ). 31

Schematic representation of
Transduction and colony forming assay
HeLa cells were incubated with a genomic particle per cell ratio (GOI) of 5,000 of either AAV-SMAR or AAV-ΔSMAR. Twenty-four hours after transduction, the percentage of eGFP-positive cells was determined by flow cytometry (BD FACSCanto™; BD Biosciences, Heidelberg, Germany). Cells were then either maintained as mixed populations in the presence of 2.5 μg/mL of blasticidin for 3 weeks followed by cultivation in the absence of selection, or seeded for colony forming assays, as described below. For experiment (Exp) 1 and Exp 2, cell suspension containing a mixture of eGFP-positive and eGFP-negative cells was seeded to yield 105 eGFP-positive cells per 15 cm2 dish or 150 cm2 flask. For Exp 3, cells were sorted for eGFP expression prior to seeding 105 eGFP-positive cells per plate. Twenty-four hours after seeding, 2 μg/mL blasticidin (Invitrogen) was added. With the next medium exchange, the blasticidin concentration was increased to 2.5 μg/mL and maintained at this concentration for 3–4 weeks. Surviving cell colonies were either picked and re-cultured without blasticidin, or washed, stained with crystal violet, and counted, as described previously. 32 Crystal violet stained colonies from the independently performed Exp 1–3 were quantified using Fiji ImageJ software setting the threshold for colony size to 200 pixel 2 for consideration. Mean and standard deviation of colony establishment was calculated from counting individual dishes (Exp 1 and Exp 2: n = 5; Exp 3: n = 8).
Southern blot
Total genomic DNA 33 was isolated from indicated colonies and treated with a restriction enzyme that cuts once per AAV vector genome (XbaI for AAV-SMAR and PciI for AAV-ΔSMAR). Southern blot analyses were performed, as described before, 14,34 using pEPitoΔSMAR as a probe. The probe was labeled with α 32 P[dCTP] and α 32 P[dATP] (6,000 Ci/ mmol each; Prime-It II Random Primer Labeling Kit; Agilent, Santa Clara, CA), and signals were detected using either PhosphoImager (Bio-Rad, Hercules, CA) or conventional x-ray films.
Fluorescent in situ hybridization
Fluorescent in situ hybridization (FISH) analyses were performed using metaphase spreads from colonies transduced with either AAV-SMAR or AAV-ΔSMAR and cultivated for at least 70 days in the absence of blasticidin. Cells were treated with 0.02 μg/mL colcemid (Biochrom, Berlin, Germany) for 24 h. Cells arrested in metaphase were harvested by mitotic shake-off, and metaphase plates were prepared as described elsewhere. 35 DIG-labeled plasmid DNA (DIG DNA labeling Kit; Roche, Basel, Switzerland) of pEPitoΔSMAR was used as a probe. Hybridization was performed overnight at 37°C in a humid chamber, followed by washing steps. DIG-labeled samples were visualized by staining with mouse anti-DIG antibody (D8156; Sigma–Aldrich) and goat anti-mouse antibody conjugated with Alexa488 (A-11029; Invitrogen). Chromosomes were counterstained with 10 μM of Sytox® Orange Nucleic Acid Stain (Invitrogen).
5-bromo-2′-deoxyuridine immunoprecipitation
For detection of replicating DNA, HeLa cells stably maintaining either AAV-S/MAR or AAV-ΔS/MAR were cultivated in the presence of 5-bromo-2′-deoxyuridine (BrdU; B5002; Sigma–Aldrich). Immunoprecipitation of BrdU-labeled DNA was performed following a recently published protocol. 36 Briefly, 50 μmol/L BrdU was added to asynchronously growing cells or cells synchronized in G1/S-phase, respectively. Cells were harvested, and 2.5 × 105 cells were lysed in the presence of 0.2 mg/mL of proteinase K and 0.05 mg/mL of glycogen for 1 h at 56°C, followed by phenol–chloroform extraction and ethanol precipitation. Purified DNA was sheared to an average size of 400 bp (Bioruptor Standard, 30/90 s on/off time, 12 min) and heat-denatured for 5 min. BrdU-labeled DNA was incubated with 12.5 μg/mL of anti-BrdU antibody (mouse, #555627; BD Biosciences Pharmingen) for 20 min at room temperature (RT) followed by incubation with 20 μg of anti-mouse IgG antibody (rabbit, M-7023; Sigma–Aldrich) for an additional 20 min at RT. DNA–antibody complexes were collected by centrifugation (16,000 g, 5 min), treated with proteinase K (0.25 mg/mL), and purified using phenol–chloroform extraction and ethanol precipitation. Subsequently, samples were analyzed by qPCR (Luminaris Color HiGreen qPCR Master Mix, K0392; Thermo Fisher Scientific) using primers specific for either blasticidin (fwd 5′-TCGCGATCGGAAATGAGAAC-3′, rev 5′-AGGGCAGCAATTCACGAATC-3′) or hCMVe/EF1 promoter region (fwd 5′-CGTCAATGGGTGGAGTATTTACG-3′, rev 5′- GGTCATGTACTGGGCATAATGC-3′).
Statistical analyses
Statistical analyses and significance tests were performed applying unpaired, two-tailed t-tests using the GraphPad software. p-Values of <0.05 were considered as significant.
Results and Discussion
Presence of an S/MAR element in AAV vector genomes increases colony formation in a competing environment
The bicistronic AAV2-based vector plasmids pAAV-S/MAR and pAAV-ΔS/MAR, which both encode for eGFP and BSD were constructed (Fig. 1A and B). pAAV-ΔS/MAR corresponds to a standard single-stranded AAV vector genome construct and served as control, whereas pAAV-S/MAR additionally contained a 2 kb S/MAR element of the human β-interferon gene. These constructs were packaged as AAV2 vectors. The resulting vectors AAV-S/MAR and AAV-ΔS/MAR were characterized as follows: The amount of vector genome containing particles was measured by qPCR using transgene specific primers revealing a genomic particle titer of 2.4 × 1011/mL for AAV-S/MAR and of 6.9 × 1011/mL for AAV-ΔS/MAR. In addition, HeLa cells were transduced with a serial dilution of the vector preparation followed by flow cytometry measuring eGFP-expressing cells to determine the biological activity or transducing particle titer of the preparations (AAV-S/MAR 3.1 × 109/mL, AAV-ΔS/MAR 4.7 × 109/mL). Both genomic and transducing particle titers were comparable, indicating that incorporation of the S/MAR element interferes neither with vector genome packaging nor with infectivity.
To investigate whether the S/MAR element is functional in the context of AAV vectors, colony forming assays were performed. Specifically, HeLa cells were transduced with either AAV-S/MAR or AAV-ΔS/MAR at a GOI of 5,000. The percentage of transduced cells was determined 24 h post-transduction revealing, as expected, a comparable number of transduced cells for each preparation (Fig. 1C). A mixed cell suspension containing in total 105 eGFP-positive cells was then seeded. Forty-eight hours after transduction, blasticidin was added to the culture medium to select cells stably expressing from the vector genome. After up to 4 weeks of selection, surviving colonies were quantified. It was found that HeLa cells transduced with AAV-S/MAR resulted in a significantly higher number of blasticidin-resistant colonies compared to cells treated with AAV-ΔS/MAR (Exp 1 and Exp 2; Fig. 1C). Interestingly, when only eGFP-positive cells were seeded prior to selection, no significant difference in colony formation could be observed (Exp 3; Fig. 1C). This suggests an establishment advantage of AAV-S/MAR over AAV-ΔS/MAR in a competing environment with non-transduced cells, facilitated by the S/MAR element. However, establishment of a stable transduction in proliferating cells is, for both AAV-S/MAR and AAV-ΔS/MAR, a rare event, as <0.5% of AAV-S/MAR and <0.25% of AAV-ΔS/MAR treated cells formed colonies (Supplementary Figs. S1 and S2A; Supplementary Data are available online at
S/MAR sequence facilitates episomal maintenance of AAV genomes in proliferating cells
To analyze whether episomal or integrated vector genomes served as template for long-term transgene expression, HeLa cells were transduced with AAV-S/MAR or AAV-ΔS/MAR and seeded in a colony forming assay, as described above. Four eGFP-positive and one eGFP-negative blasticidin-resistant AAV-S/MAR-derived colonies were then picked and cultured further in the absence of blasticidin. The four eGFP-positive AAV-S/MAR colonies showed stable eGFP expression for at least 56 population doublings (70 days) in the absence of selection pressure (Fig. 2A), indicating mitotic stability of the AAV-S/MAR genome, while eGFP expression for colony #4 remained, as expected, below background level.

Mitotic stability of AAV-S/MAR vector genomes in HeLa.
Southern blot analyses were then performed to distinguish the episomally replicating AAV from integrated vector genomes. However, Southern blot analyses were challenging to perform due to the small size and low copy numbers of the vector genome. 34, 37
For eGFP-positive AAV-S/MAR-derived colonies #2, #5, and #8, a 4.6 kb band was detected, which indicated that the vector DNA prevailed either as single circular episomes or head-to-tail concatemers (Fig. 2B). Vector DNA from colony #9 formed a tail-to-tail concatemer, as shown by the 7.9 kb signal. Integrated vector DNA that should result in a 4.6 kb (one vector genome or multiple head-to-tail genomes) or 7.9 kb (multiple tail-to-tail genomes) band escorted by high-molecular genomic DNA bands 37 could not be detected. For the eGFP-negative colony #4, vector genomes were not detected, which was in line with absence of transgene expression.
As an independent method, FISH was then performed on metaphase spreads of these colonies. FISH on metaphase spreads enables detection of even rare integration events as hybridization signals on both sister chromatids. In contrast, a hybridization signal on only one of the sister chromatids is indicative of episomes, since they non-covalently associate with host chromosomes. 38 For each colony, at least three different metaphase plates were analyzed. In all cases, FISH analyses confirmed the Southern blot results. Specifically, only episomal signals were detectable for the AAV-S/MAR colonies #2, #5, #8, and #9 (Fig. 2C, white boxes). Southern blot and FISH analyses of three further AAV-S/MAR colonies obtained from another independently performed experiment showed equivalent results (Supplementary Fig. S3).
To distinguish monomeric from concatemeric episomes, qPCR analyses were performed using eGFP-specific primers. Again, AAV-S/MAR genomes were not detected in colony #4 (Supplementary Fig. S4). Since this colony survived selection, a spontaneously developed blasticidin resistance is conceivable—a phenomenon that has been observed before. 39 For the remaining colonies, an average copy number of 6 was determined (Supplementary Fig. S4). Considering the average number of one to two episomes per cell detected in metaphase FISH (Fig. 2C), these results argue for AAV-S/MAR genomes being maintained as episomal concatemers in either head-to-tail (#2, #5, and #8) or tail-to-tail (#9) conformation (Fig. 2B).
Analogously to the AAV-S/MAR-derived colonies, three eGFP-positive and one eGFP-negative, blasticidin-resistant AAV-ΔS/MAR-derived colonies were also picked for further analysis. Again, eGFP-positive AAV-ΔS/MAR-derived colonies Δ4, Δ5, and Δ6 displayed stable long-term eGFP expression in the absence of selection (Fig. 3A). Surprisingly, it was found that the observed vector genome maintenance was not necessarily the result of integration: Southern blot analyses revealed episomal signals for all three eGFP-positive colonies (Δ4, Δ5, and Δ6), either as single circular episomes or head-to-tail (2.6 kb, colonies 4 and 6) or tail-to-tail concatemers (4.8 kb, colony Δ5; Fig. 3B). In the eGFP-negative colony (Δ1), no presence of vector genomes was detected in Southern Blot analysis, despite its apparent insensitivity to blasticidin. Again, the colonies were analyzed by FISH on metaphase spreads. 37,38 In one colony (Δ6), rare integration signals were detected (1 signal in at least 10 analyzed metaphase plates; Fig. 3C) in addition to episomal signals, indicating that rare integration events in the absence of the S/MAR do occur. For all other colonies, however, hybridization signals are in line with pure episomal maintenance of vector genomes (Fig. 3C). Based on the metaphase plate analyses, an average of three AAV-ΔS/MAR signals per cell were estimated (Fig. 3C).
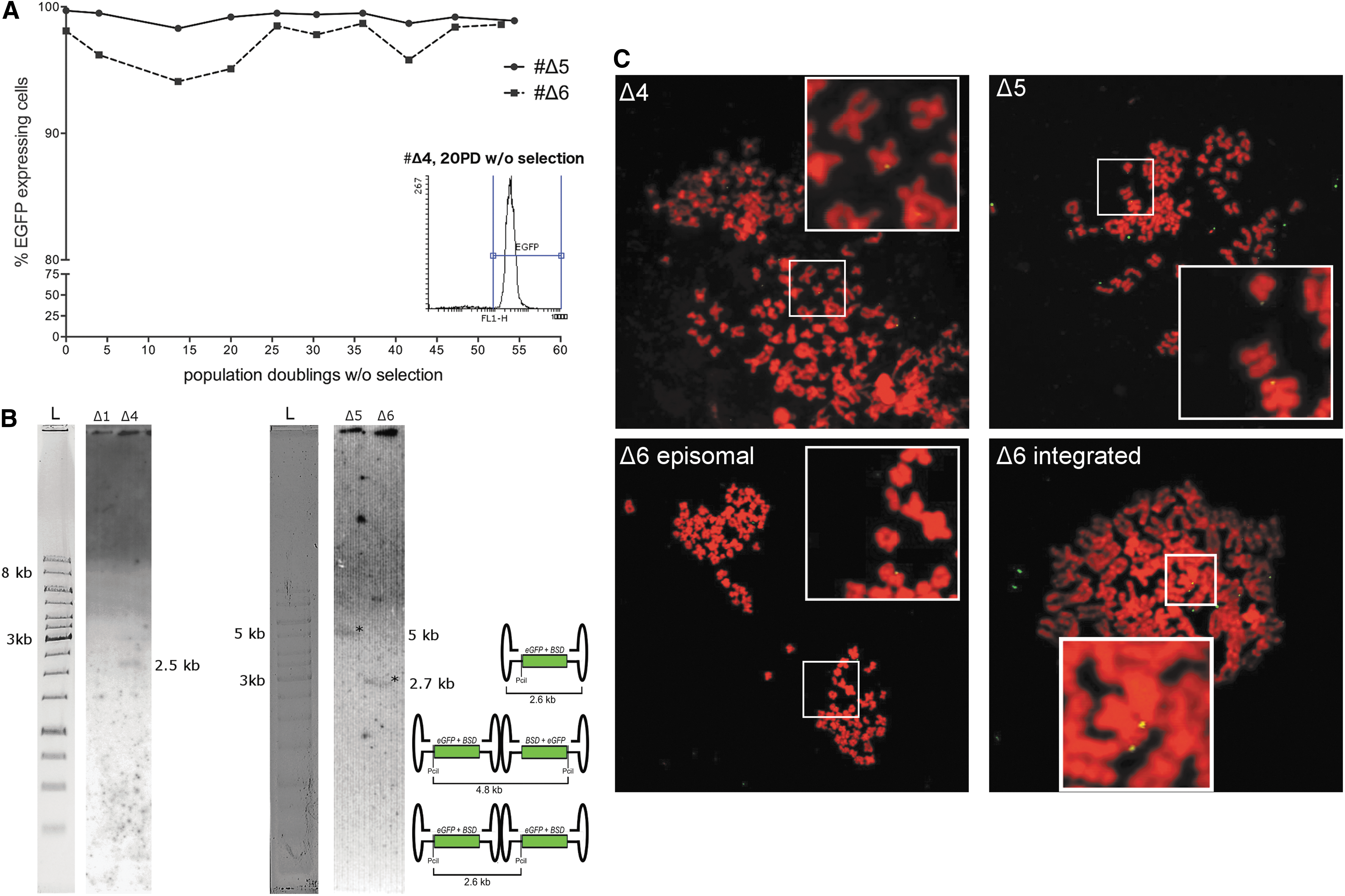
Mitotic stability of AAV-ΔS/MAR vector genomes in HeLa cells.
Repeating cell transduction and colony forming assays with lower GOIs (GOI 1,000) also demonstrated episomal maintenance of both AAV-S/MAR and AAV-ΔS/MAR vector genomes (Supplementary Fig. S2A and C).
An initial selection phase is required to establish mitotically stable AAV episomes
It has been previously shown that plasmid-based S/MAR vectors are maintained episomally. 15,40,41 Here, it was demonstrated that insertion of an S/MAR element in an AAV vector improves the efficiency but is not required for AAV episome retention in proliferating HeLa cells. In contrast to this study, Kymäläinen et al. explored a truncated S/MAR module. In that first study (and to date only report), AAV vectors, both with and without an S/MAR element, failed to establish stable colonies, and thus AAV-transduced cells were diluted out with cell divisions. 42 In contrast to our study, Kymäläinen et al. did not employ initial antibiotic selection. At least in the context of plasmid-based S/MAR vector systems, however, initial antibiotic selection pressure is known to be a mandatory step to compete successfully with an environment of non-transfected cells. 35,37 Likewise, in the present study, when transduced HeLa cells were cultured in the absence of selection pressure, non-transduced cells rapidly overgrew cells transduced with either vector (Supplementary Fig. S2B). Therefore, the importance of an initial selection period for establishing mitotically stable episomes of both AAV-S/MAR and AAV–ΔS/MAR vectors in proliferating cells is emphasized. As observed for other episomal replicons, 43 for both AAV-S/MAR and AAV–ΔS/MAR, rather low vector copy numbers per cell were found.
In post-mitotic tissues, long-term transgene expression is reported from vector genomes, which are maintained as episomal circles or high-molecular weight concatemers. In addition, as consequence of cellular DNA repair activities, AAV vector genomes can be found integrated at non-homologous sites either as single copies or concatemers. 44 Although vector genome integration is a rare event (ca. 1 × 10−4–1 × 10−5), 45 this study selected for transgene expressing colonies, thereby increasing the probability of obtaining colonies in which survival is caused by vector genome integration. In line with results obtained for plasmid-based S/MAR vectors, no integration event could be detected in AAV-S/MAR-derived colonies (Fig. 2 and Supplementary Figs. 2C and 3). Unexpectedly, AAV-ΔS/MAR-derived stable colonies also maintained episomal vector genomes, as shown by two independent methods (Southern blot and FISH; Fig. 3 and Supplementary Fig. S2C). In one of the AAV-ΔS/MAR-derived colonies (Δ6), integration was detected in addition to episomally maintained genomes (Fig. 3), proving the sensitivity of the assay system and revealing that integration of standard vector genomes does occur, albeit at a lower frequency than expected. This observation is particularly remarkable, as it contradicts previous studies that employed selection pressure as a common strategy to induce mitotic stability of AAV vector genomes in proliferating cells through integration. 7,46,47 The latter studies commonly used a rather high selection pressure, 7,46,47 while this study used moderate concentration of the antibiotic (up to 2.5 μg/mL of blasticidin, fourfold less than the manufacturer recommends for selecting HeLa cells).
AAV-S/MAR and AAV-ΔS/MAR genomes replicate in proliferating HeLa cells
Since maintenance of the extrachromosomal state in proliferating cells requires replication, we performed a BrdU-based replication assay. Briefly, the AAV-S/MAR- and AAV-ΔS/MAR-derived colonies were incubated with BrdU to label newly synthesized DNA. BrdU-labeled DNA was then immunoprecipitated and analyzed by qPCR, using unlabeled DNA of the same colonies as a control. 36 BrdU incorporation and thus AAV vector genome replication was shown for all colonies, independent of whether they were derived from AAV-S/MAR or AAV-ΔS/MAR-transduced cells (Fig. 4). Notably, in asynchronously growing cell populations, a short labeling pulse (3 h) resulted in rather low BrdU signals for AAV-ΔS/MAR-derived colonies compared to AAV-S/MAR-derived colonies (Fig. 4A). In contrast, BrdU signals of AAV-ΔS/MAR colonies were higher when cells were labeled for 18 h (Fig. 4B), possibly due to differences in replication timing of AAV-S/MAR and AAV-ΔS/MAR vector genomes. When cells were synchronized in late G1-phase and released into S-phase for 3 h (early S-phase), BrdU signals for the AAV-ΔS/MAR-derived colonies Δ4 and Δ5 were at background level (Fig. 4C), indicating replication of AAV-ΔS/MAR vector genomes in late S-phase. In contrast, in Δ6, the only AAV-ΔS/MAR-derived colony in which an integrated vector copy as well as episomes had been detected, the level of replication was found to be similar to levels measured for the AAV-S/MAR-derived colonies. Human β-globin, a late replicating gene (S/G2-phase), served as a control and showed low BrdU signals in early S-phase, while signals were comparable to AAV-S/MAR and AAV-ΔS/MAR in asynchronously growing cells (Fig. 4D). Thus, it was shown for the first time that AAV vector genomes replicate in an episomal configuration. The portion(s) of AAV vector genome serving as origins of replication in the absence of S/MAR remain to be identified. Although this remains to be proven, the most likely candidate sequences for mediating episomal replication and mitotic segregation of “common” AAV vector genomes are the ITR sequences, the sole viral remnants in AAV vector genomes. These sequences serve as origin of replication but normally require helper virus factors as well as the viral Rep proteins for this function. 8
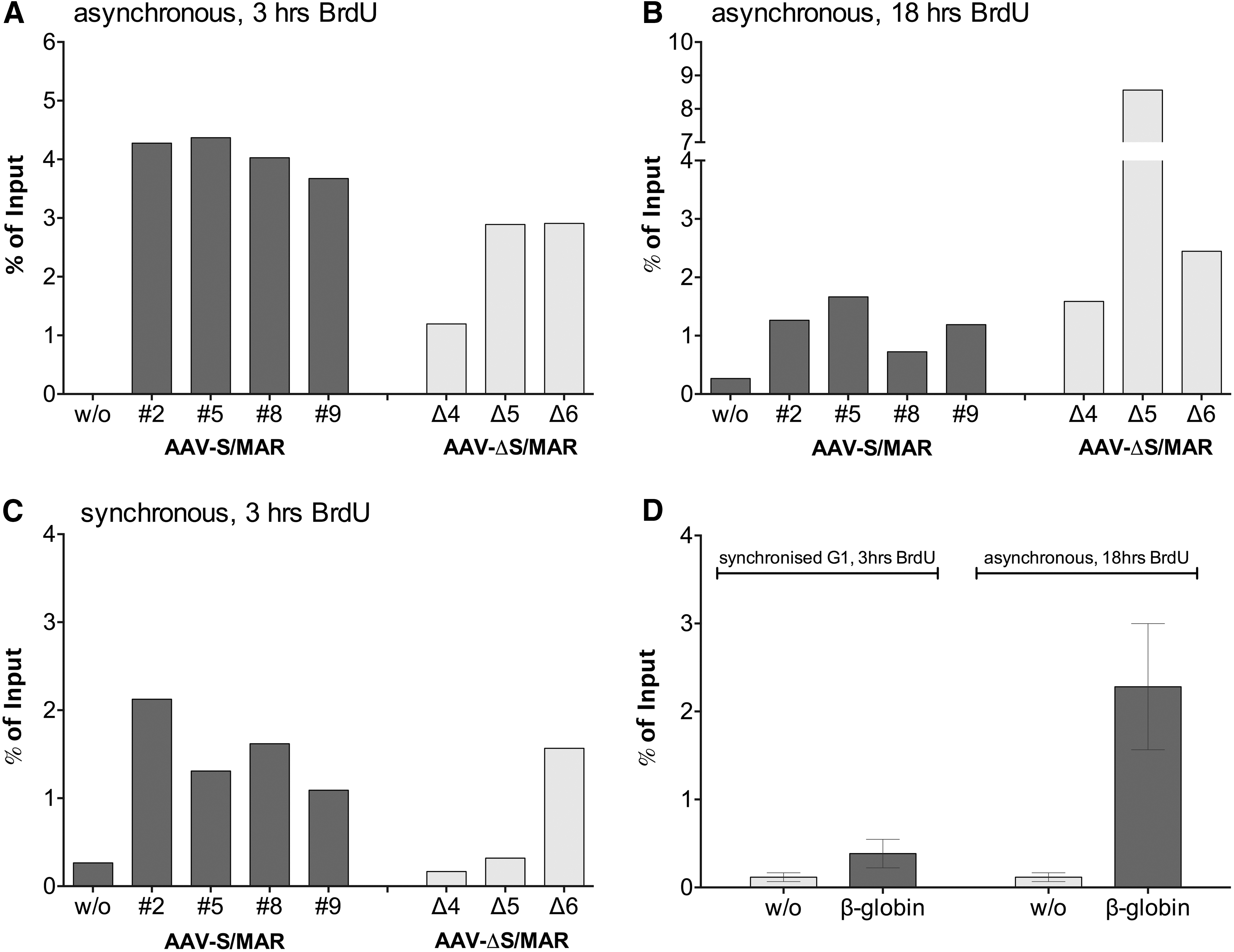
AAV vector genomes replicate in proliferating HeLa cells. Indicated colonies of AAV-S/MAR and AAV-ΔS/MAR-treated HeLa cells, stably expressing eGFP in the absence of selection pressure, were subjected to 5-bromo-2′-deoxyuridine (BrdU) replication assay. BrdU incorporation was detected for both AAV-S/MAR and AAV-ΔS/MAR vector genomes when asynchronous cells were labeled with BrdU for either
Incorporation of a foreign genomic element in addition impacts on the replication timing (Fig. 4). Specifically, episomes with S/MAR replicate in an early stage of the S-phase, while in the absence of S/MAR, replication of AAV episomes is delayed to the late S-phase suggesting that an alternative set of cellular factors contribute. One candidate—in case AAV vector and viral genome replication indeed employs the same factors—is the Mre11-Rad50-Nbs1 (MRN) complex. MRN exerts its function in S/G2 phase, possesses a DNA tethering activity, and was reported to regulate wild-type AAV replication positively. 48
In conclusion, this study proof-of-principle reports that AAV vector genomes are capable to establish as replicating episomes, at least in HeLa cells. Low establishment efficiencies (<0.25%) indicate that episomal establishment and replication of vector genomes is a possible but rare event. However, applying mild selection conditions might enable selection for these rare events. Whether episomal replication of AAV vector genomes is a phenomenon restricted to transformed, tumorigenic cell lines such as HeLa 49,50 or a more general feature remains to be elucidated. Similarly, the underlying mechanism as well as specific factors or conditions needed for replication remain unknown and should be investigated in further studies. Nevertheless, incorporation of an autonomously replicating S/MAR unit facilitates establishment of replicating AAV episomes in a competing environment, a finding that may open a new perspective for AAV vectors in gene therapeutic applications.
Footnotes
Acknowledgments
This work was supported by grants from the priority programs of the Deutsche Forschungsgemeinschaft SPP1230 (H.B., BU1310/1-2; and H.J.L., LI231/25-2), Alexander von Humbold Fundation (H.A.), Peter and Traudl Engelhorn foundation (C.H.), as well as by the Center for Molecular Medicine Cologne (CMMC), University of Cologne (H.B.; CMMC-C1). We thank Jude Samulski (University of North Carolina, Chapel Hill, NC) for providing pXX6, and Rudi Haase for providing pEPito-eGFP-IB.
Author Disclosure
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.