
Abstract
Fibrotic liver injury is a significant healthcare burden in the United States. It represents a major cause of morbidity and mortality for which there are no effective Food and Drug Administration–approved treatment strategies. Fibrosis is considered a disruption of the normal wound healing responses mediated by fibroblastic cells, which are triggered and sustained by pro-fibrotic cytokines such as transforming growth factor beta 1 (TGF-β1). TGF-β1-mediated trans-differentiation of hepatic stellate cells (HSCs) from quiescent to activated myofibroblasts is a pivotal event in the development of fibrosis. Activation is accompanied by global changes in microRNA (miR) expression. It has been previously reported that miR19b is decreased in activated HSCs and contributes to increased expression of TGF-β receptor II and connective tissue growth factor, both confirmed targets of miR19b. An adeno-associated virus serotype 2 vector (AAV2) with a miR19b transgene downstream of enhanced green fluorescent protein under the murine collage alpha 1(I) promoter was developed specifically to target HSCs. Male Sprague Dawley rats (250 g) underwent sham or bile-duct ligation (BDL) surgery. Directly after BDL, rats received AAV2-miR19b, AAV2-control, or vehicle normal saline (NS) by portal-vein injection. After 2 weeks, the animals were euthanized, and blood was collected for alanine and aspartate aminotransferase, total and direct bilirubin, and alkaline phosphatase. Tissue was collected for RNA and protein extraction and histology. Fibrosis and measures of hepatic injury were significantly reduced in AAV2-miR19b-treated rats in combination with significant improvements in total and direct bilirubin. Histological analysis of collagen by PicroSirius Red staining revealed a ∼50% reduction compared to AAV2-control or NS-injected animals. Pro-fibrotic markers, smooth-muscle alpha-actin, TGF-β receptor II, and collagen alpha 2(I) mRNA and protein were significantly decreased compared to AAV2-control and NS groups. AAV2-mediated reintroduction of miR-19b, specifically expressed in HSCs, improved liver function, inhibited fibrosis, and improved measures of hepatic injury in a BDL model.
Introduction
C
Liver injury is a dynamic process beginning with hepatocyte cell death and oxidative stress that results in recruitment of resident macrophages, Kupffer cells, and hepatic stellate cells (HSCs). 1,9 Activation of HSCs from a quiescent vitamin A–storing cell to a proliferating, contractile myofibroblast is a pivotal event during fibrogenesis. 10 HSCs maintain an extensive role in fibrotic injury as the primary source of collagen and ECM proteins, immunomodulatory cytokines, and growth factors, and they are required for hepatocyte repopulation. 11,12 HSC trans-differentiation is associated with extensive changes in protein coding mRNA expression, as well as microRNA (miR) profiles both within the HSCs and those packaged into exosomes and released into the hepatic microenvironment. 13,14 Liver injury is associated with major alterations in both hepatic and circulating miRs. Injured hepatocytes release transforming growth factor beta 1 (TGF-β1), a key cytokine in stimulating fibrotic injury and crucial for regulating HSC activation and stimulating collagen synthesis. 1,9,10 This process is tightly regulated by translational, transcriptional, and post-transcriptional pathways that include epigenetics and miRs. Changes in miR expression profiles are often cell specific, and during hepatic injury, these changes are closely associated with HSC trans-differentiation. 9,15,16 miRs are small (18–22 nt), non-coding RNA that post-transcriptionally regulate protein coding mRNA by translational inhibition or degradation through mRNA destabilization, but may also modulate transcription and DNA methylation processes. 17 Further, miRs may act as molecular switches to alter signaling pathways and redirect cell functions and phenotype. 18
The authors' laboratory has previously reported that miR19b is significantly downregulated during HSC trans-differentiation. The studies revealed that cellular loss of miR19b on HSC activation enhanced TGF-β1 signaling through disruption of miR19b's inhibitory effects on TGF-β receptor II (TGF-βRII) expression and connective tissue growth factor (CTGF) signaling during fibrosis in rodent models of hepatic injury and in human fibrosis. 13,14,19 The fibrotic response in HSCs was reversed by the forced overexpression of miR19b in vitro. 13 The current studies presented herein focused on the in vivo application of modulating miR19b expression in HSCs in a rodent model of hepatic injury to inhibit TGF-β-mediated fibrogenesis. 1 An adeno-associated virus serotype 2 vector 20,21 (AAV2), carrying either the mature miR19b transgene under control of a collagen promoter and downstream of enhanced green fluorescent protein (EGFP) or a truncated version lacking the mature miR and seed sequence (control), was injected at 1 × 1012 viral genomes (vg) via the portal vein at the time of bile-duct ligation (BDL). Two weeks following BDL, animals treated with AAV2-miR19b had decreased markers of liver injury and improved liver function, with a reduction in collagen deposition and other pro-fibrotic markers. To date there has been considerable emphasis placed on targeting the HSC to ameliorate fibrotic injury in the liver. The present data indicate that AAV2-mediated reintroduction of miR19b in a rodent model of liver injury can successfully inhibit fibrosis and improve liver function through targeted action on collagen-producing HSCs.
Materials and Methods
Plasmids
The AAV2 constructs were produced by the Vector Core at University of Pennsylvania (
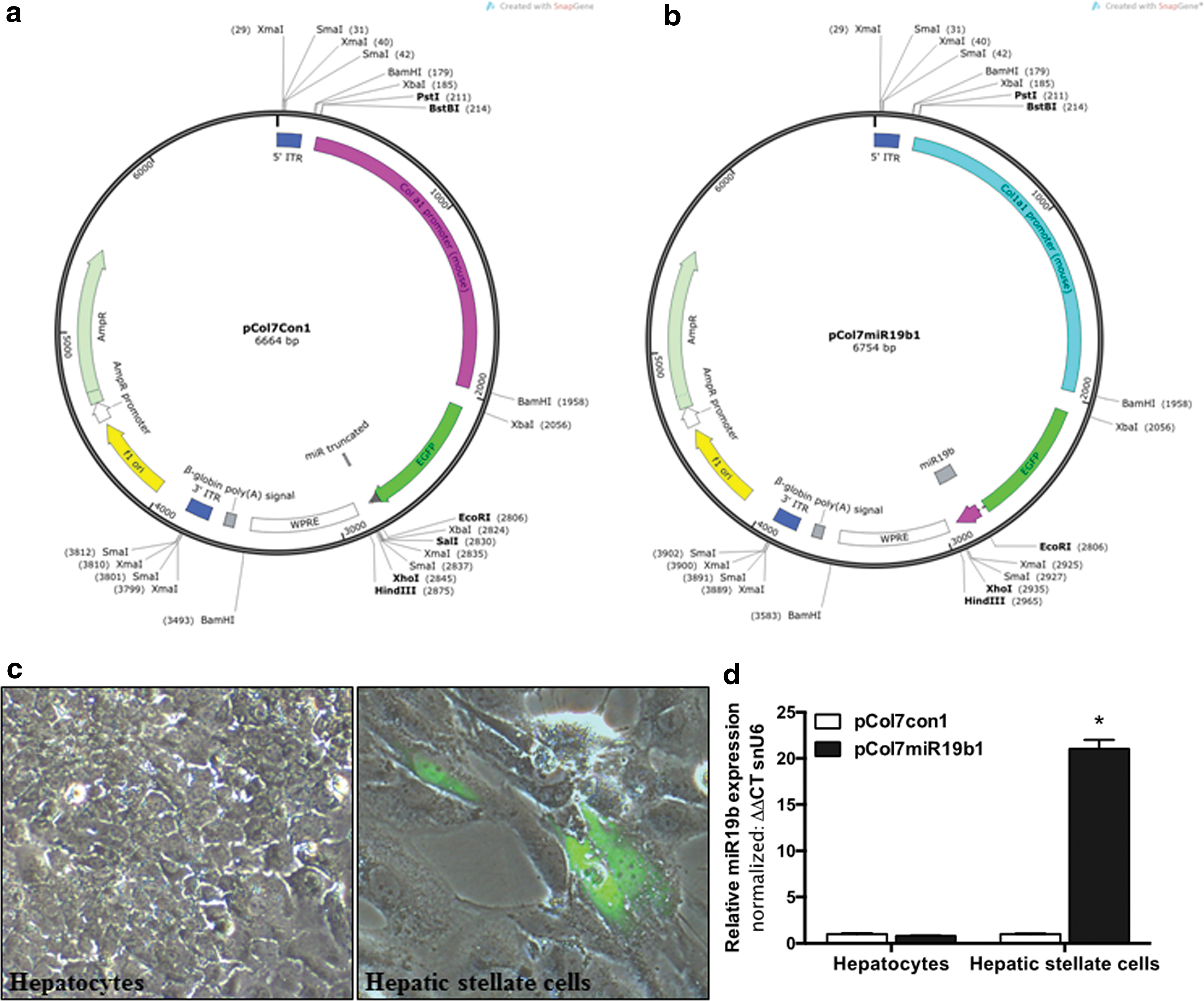
Adeno-associated virus (AAV) allows for targeted delivery of miR19b transgene to activated hepatic stellate cells (HSCs). Plasmid maps for pCol7Con1
Sequences for truncated control and miR19b transgenes
Underlined sequence marks miR19b target sequence.
Cell culture
Primary hepatocytes and HSCs were isolated from Sprague Dawley rats, as previously described. 22 Hepatocytes were maintained no more than 48 h on collagen-coated plates. 22 Cells were transfected with miR19b or control plasmid using CytoFect for hepatocytes or fibroblasts (Cell Applications, San Diego, CA) per the manufacturer's protocol.
BDL
Male Sprague Dawley rats (200–250 g) were randomly assigned to groups for BDL or sham surgery per the approved Institutional Care and Use of Animal Committee protocol and following the Guide for The Care and Use of Laboratory Animals. Animals were anesthetized by isoflurane. Using aseptic conditions, a midline laparotomy was performed, and the common bile duct located, doubly ligated, and transected. Following transection, animals were randomly assigned to receive either 100 μL of AAV2-miR19b, AAV2-control at 1 × 1012 vg suspended in normal saline (NS), or 100 μL of NS and injected into the portal vein using an ultrafine insulin syringe (31 gauge). Bleeding was staved; the abdominal wall and skin were closed with sutures. Animals were necropsied 2 weeks post BDL, blood sample drawn, and tissues flash frozen or preserved in buffered formalin followed by paraffin embedding.
Laboratory chemical analysis
Blood was collected by cardiac puncture in heparinized tubes and sent to an in-house laboratory (Carolinas Medical Center) for analyses of alkaline phosphatase, albumin, alanine aminotransferase (ALT), aspartate aminotransferase (AST), and total and direct bilirubin.
Total RNA isolation and qRT-PCR
Total RNA was prepared from flash-frozen tissue using a phenol/chloroform extraction and in-column DNase treatment (Zymo, Irvine, CA). microRNA or mRNA were analyzed by qRT-PCR using gene specific primers on a BioRad CFX96 real-time PCR thermal cycler (BioRad, Hercules, CA). cDNA was generated from total RNA using iScript cDNA synthesis kit followed by qRT-PCR using SYBR Green Super Mix (BioRad) per the manufacturer's protocols. Alternately, miR19b expression was determined from cDNA generated using NCode ViLo miRNA cDNA synthesis kit and Express SYBR-Green miR qRT-PCR kit (Life Technologies, Carlsbad, CA) per the manufacturer's protocols. Fold changes were calculated using comparative Ct analysis and normalized to GAPDH or snU6 as appropriate.
Western blot
Total protein lysates from whole liver tissue were prepared using TePer™ (Thermo Fisher Scientific, Grand Island, NY). Protein concentration was determined by BCA protein assay kit (Thermo Fisher Scientific). Equal amounts of protein were loaded on a 4–12% Bis-Tris gel (Life Technologies) and transferred to a polyvinylidene fluoride membrane (Thermo Fisher Scientific). The following primary antibodies were used: anti–smooth muscle alpha-actin (αSMA; ab21027), purchased from Abcam (Cambridge, MA); collagen1α2 (Col1α2; sc-8788), TGF-βRII (sc-220R), and glyceraldehyde-3-phosphate dehydrogenase (GAPDH; sc-48166) were purchased from Santa Cruz Biotechnology (Dallas, TX). Ponceau staining was used to confirm equal protein loading, and expression was normalized to GAPDH.
Immunohistochemistry
Sections (4 μm) of formalin-fixed paraffin-embedded liver tissue were blocked with 2.5% normal horse serum and incubated with anti-CD68 antibody at 1:250 (ab125212; Abcam) followed by diaminobenzidine (DAB) chromogen for macrophage detection. For detection of neutrophil infiltration, a Leder stain, Naphthol-ASD Chloroacetate Esterase (Sigma–Aldrich, St. Louis, MO), was used, and sections were counterstained with Gill's hematoxylin. For dual staining, paraffin-embedded liver blocks were sectioned (5 μm) and mounted followed by dual staining for GFP (38689; Abcam) and Desmin (sc-7559). Positive and negative liver slides for each antibody were stained with each antibody individually then combined for dual staining. ImmPRESS peroxidase polymer detection system (mouse) was used to detect bound secondary antibody following the manufacturer's instructions (Vector Laboratories, Burlingame, CA). Visualization of the GFP antibody–polymer complex was performed using DAB (brown chromagen). Incubation with the second primary antibody (Desmin) was followed by application of ImmPRESS goat secondary. Visualization of the Desmin antibody–polymer complex was performed using Vector VIP peroxidase substrate (purple chromagen). Stained slides were counterstained with methyl green. The dual staining was carried out according to the Multiple Antigen Labeling guide published by Vector Laboratories using ImmPress peroxidase detection systems. Representative sections were examined microscopically (40 × ) on an Olympus BX40 microscope (Olympus America, Inc., San Jose, CA) with a DP72 camera and CellSense standard 2.0 software (Olympus America, Inc.).
Statistical analyses
Data are presented as the mean ± standard error of the mean. All experiments were performed a minimum of three independent times unless otherwise stated. Statistical analyses were performed using one- or two-way analysis of variance for grouped comparisons, with Tukey's multiple comparison test or paired Student's t-test. p-Values of <0.05 were considered significant.
Results
Expression of EGFP-miR19b under the collagen promoter is specific to HSCs
Tissue-specific loss of miR19b in the liver during fibrotic injury led to the development of an AAV2 vector to reintroduce miR19b targeting the HSC. The mature miR19b transgene (pCol7miR19b1) or a truncated miR19b (pCol7Con1), lacking the mature sequence and seed region (Table 1), was fused to EGFP and cloned under the control of the murine collagen alpha 1(I) promoter (Fig. 1a and b). Initially, the study sought to confirm that expression of EGFP-miR19b transgene would be specific to HSCs. Primary HSCs were culture-activated, and on day 7, media was changed and cells transfected with the pCol7miR19b1 plasmid. Cells were transfected with a GFP plasmid under the cytomegalovirus promoter (pCMV-GFP) to monitor transfection efficiency (data not shown). Cells were transfected using cytofect for fibroblasts (Cell Applications). Primary hepatocytes were isolated and plated to collagen-coated tissue culture plates. Media was changed after 4 h and cells transfected with the miR19b or control plasmids using cytofect transfection reagent for hepatocytes (Cell Applications). EGFP expression was examined by fluorescent microscopy 24 h after transfection. EGFP expression was limited to primary HSCs transfected with pCol7miR19b1 (Fig. 1c, right panel). Similar results were observed with pCol7con1 (data not shown). EGFP expression was not detected in primary hepatocytes (Fig. 1c, left panel), despite a 65–70% transfection efficiency identified with pCMV-GFP expression in both hepatocytes and HSCs (data not shown). In addition, miR19b expression measured by qRT-PCR was significantly increased in HSCs transfected with pCol7miR19b1 compared to pCol7con1, with no increase identified in primary hepatocytes (Fig. 1d), supporting the specificity of the plasmid construct for the AAV vector.
AAV2-mediated reintroduction of miR19b inhibits pro-fibrotic gene expression in primary HSCs
To assess the efficacy of the AAV2 construct, a dose response was performed. Primary HSCs were plated and culture-activated, and on day 4 cells were treated with tryphostin-1 500 μM/mL for 4 h. 20 Media was replaced and cells transduced at a MOI of 5,000, 10,000, 20,000, or 50,000 vg/cell of AAV2-miR19b. miR19b expression was measured after 24 h by qRT-PCR. These data confirmed a significant increase in miR19b at 5,000 vg/cell, with no significant difference at 10,000 or 20,000 vg/cell (data not shown). However, expression was decreased at a MOI of 50,000 vg/cell (data not shown). To examine the effects of AAV2-mediated miR19b expression on HSC activation and the fibrogenic response, primary HSCs were plated and culture-activated, as previously described, and transduced at a MOI of 5,000 vg/cell. At 24 h, cells were examined microscopically for GFP expression (Fig. 2a), and cells were harvested for expression analysis of αSMA, TGF-βRII, Col1α2, and miR19b by qRT-PCR. Additional cells were treated with TGF-β1 (5 ng/mL) to determine the responsiveness of the murine collagen promoter on EGFP and miR19b expression. Cells were positive for EGFP at 24 h, and there was an increase in the number of EGFP-positive cells 24 h after TGF-β1 treatment (Fig. 2a). miR19b expression was also significantly increased in AAV2-miR19b transduced cells at 24 h, and further at 48 h following TGF-β1 treatment compared to respective controls (Fig. 2b). αSMA, TGF-βRII, and col1α2 were all significantly increased following TGF-β1 treatment compared to untreated cells. However, AAV2-miR19b transduction significantly inhibited TGF-βRII mRNA at 24 h, and all pro-fibrotic markers were significantly decreased at 48 h following TGF-β1 stimulation compared to control and compared to TGF-β1-stimulated cells transduced with AAV2 control (Fig. 2c), suggesting increased activity of the collagen promoter with subsequent decreases in pro-fibrotic responses.
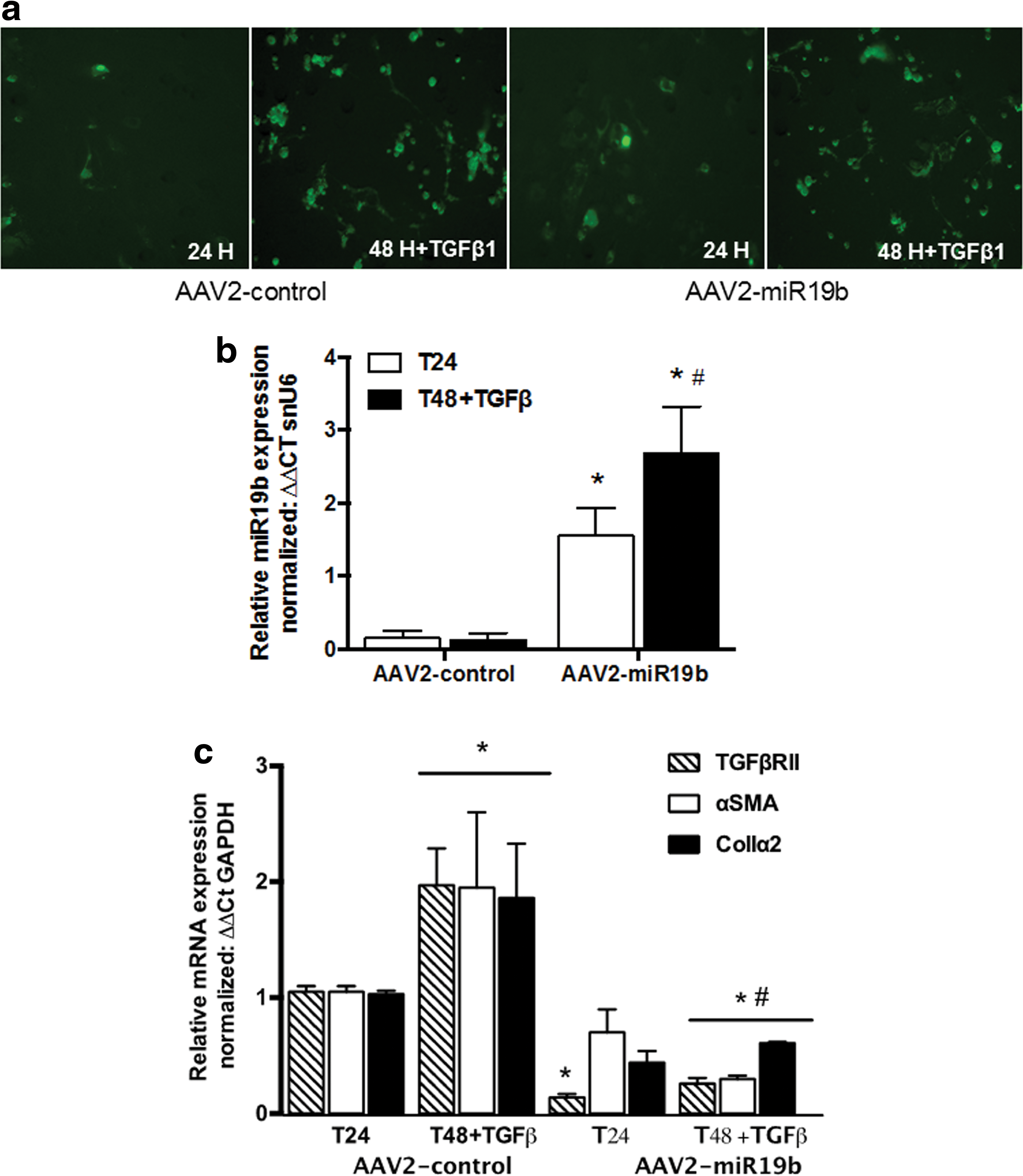
Transduction of LX2 cells with AAV2-miR19b inhibited HSC activation and fibrotic response.
AAV2-mediated expression of miR19b inhibits collagen deposition in a rodent model of BDL injury
An initial pilot study was conducted to determine a dose response for the AAV2 vector. Twenty-four Sprague Dawley rats (250 g, three per group) were randomly assigned to receive sham or BDL surgery. BDL rats were treated with vehicle or with 5 × 1011, 1 × 1012, or 2 × 1012 vg of AAV2-control or AAV2-miR19b administered in 100 μL of NS via the portal vein at the time of BDL. Increased expression of miR19b with inhibition of col1α2 mRNA was identified at a dose of 1 × 1012 (data not shown), and this dose was selected for further study. Sprague Dawley rats (250 g, six per group) were randomly assigned to undergo sham or BDL surgery and receive vehicle, AAV2-control, or AAV2-miR19b (1 × 1012 vg in 100 μL of NS) via portal-vein injection at the time of BDL. On gross examination prior to necropsy, all rats were active and displayed no signs of distress. Rats that underwent BDL had yellow ears and tail, with dark yellow urine and pale feces. On necropsy, it was noted that the visceral and parietal peritoneum were jaundiced, livers were enlarged with punctuated surfaces, and bile ducts were dilated. Microscopic examination of hematoxylin and eosin (H&E)-stained tissue (200 × ) revealed no structural changes in hepatic architecture, portal triad, or central vein in sham tissue (Fig. 3a, H&E left panel). However, histological examination of tissue from BDL rats receiving NS or AAV2-control revealed extensive damage, with alterations in hepatic architecture. Bile ducts were engorged, sinusoids distended, with numerous necrotic cells and extensive connective tissue bridging along portal tracts, accompanied by a marked increase in immune cells. Despite obvious injury and distention of the bile ducts, changes in the hepatic architecture were not as extensive in AAV2-miR19b-treated rats, with fewer invading immune cells observed. Sections were also stained with PicroSirius Red for visualization of collagen I and III. A minimum of five fields per section were imaged (100 × ) and positive staining quantified by ImageJ. 23,24 Collagen staining (Fig. 3a, right panel) was significantly increased in all BDL groups compared to sham animals, but AAV2-miR19b-treated animals measured a significant reduction in collagen staining compared to BDL-NS and BDL AAV2-control-treated animals (Fig. 3b). Analyses of col1α2 mRNA by qRT-PCR revealed similar significant increases in all BDL groups compared to sham animals, but was significantly decreased in BDL AAV2-miR19b-treated animals compared to BDL-NS and BDL AAV2-control animals (Fig. 3c). These data suggest that reintroduction of miR19b reduces fibrosis and minimizes derangements in hepatic architecture.
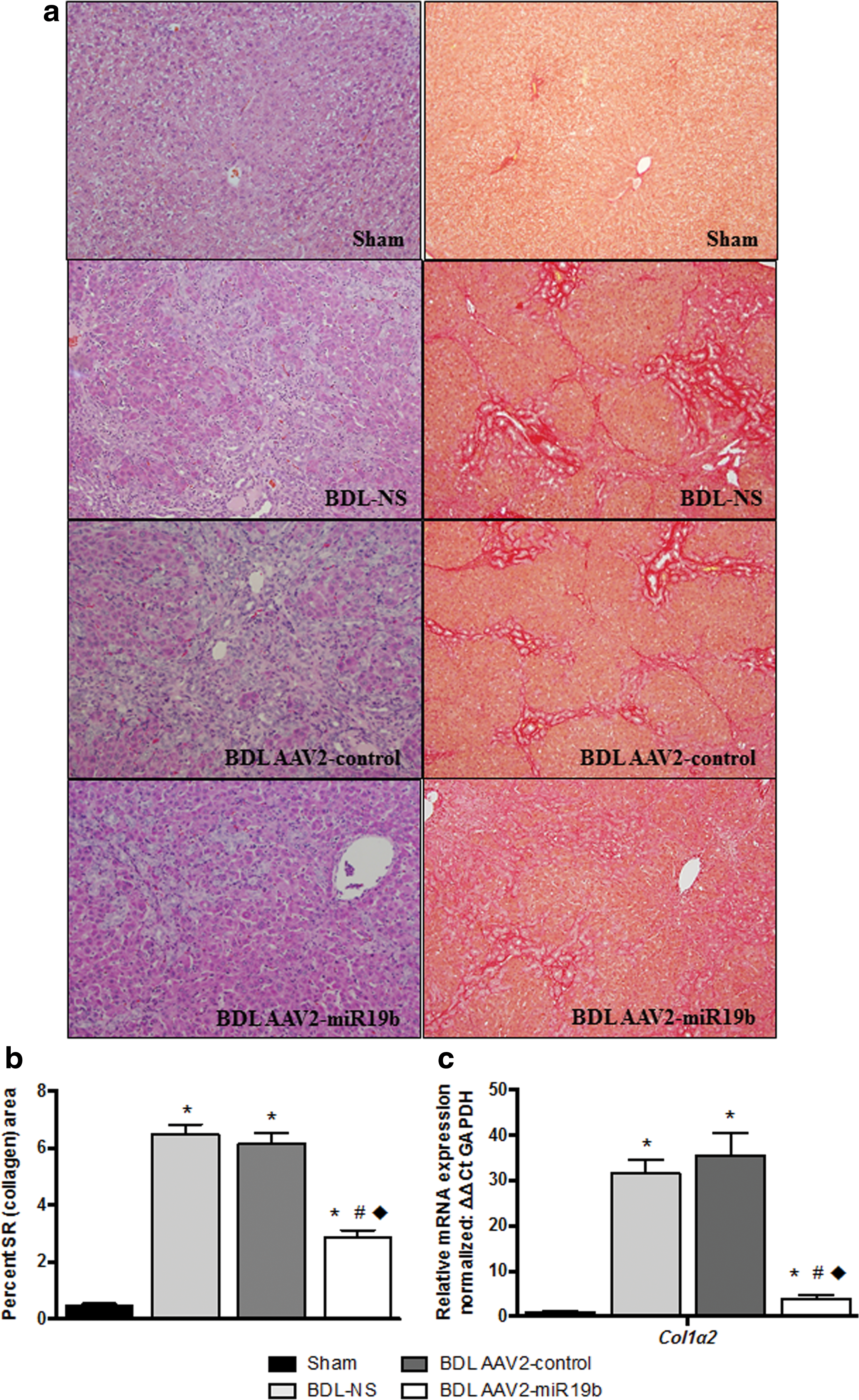
Histological examination of livers from bile-duct ligated (BDL) rats showed decreased injury and collagen deposition.
Measures of liver injury and liver function were improved in AAV2-miR19b-treated BDL rats
At necropsy animals and livers were weighed and the liver-to-body-weight ratio calculated. All ratios were significantly increased in BDL groups compared to sham animals. Body weight did not differ significantly in BDL-NS or BDL AAV2-control-treated animals compared to sham. However, body weight was significantly lower in BDL AAV2-miR19b-treated rats compared to sham (Table 2). A complete liver panel was performed by the Laboratory at Carolinas Medical Center and revealed ALT and AST were significantly increased in BDL rats compared to sham. However, AST was significantly reduced in BDL-AAV2-miR19b-treated rats compared to pair-matched BDL-AAV2-control. In addition, total (unconjugated) and direct (conjugated) bilirubin levels were determined to be significantly elevated (p < 0.05) in all BDL groups compared to sham animals. However, in BDL-AAV2-miR19b animals, measures of total and direct bilirubin were significantly reduced compared to levels in BDL-NS and BDL-AAV2-control (Table 2). AAV2-mediated reintroduction of miR19b was associated with improved liver function and decreased markers of hepatic injury.
Characteristics and liver panel data
p < 0.05 vs. sham; ** p < 0.05 vs. BDL-NS; *** p < 0.05 vs. BDL AAV2-control.
Table values represent mean ± SEM.
BDL, bile-duct ligation; NS, normal saline; AAV, adeno-associated virus; AAV2, adeno-associated virus serotype 2; ALT, alanine aminotransferase; AST, aspartate aminotransferase.
With AAV2-mediated reintroduction of miR19b expression, pro-fibrotic markers are significantly reduced in rats after BDL
Initially, expression of miR19b in total liver tissue from sham and BDL groups was examined. Expression was significantly reduced in both BDL-NS and BDL AAV2-control and increased in AAV2-miR19b-treated rats compared to sham. Expression of miR19b was significantly increased in AAV2-miR19b-treated rats compared to BDL-NS and BDL AAV2-control-treated rats (Fig. 4A). To confirm that expression of miR19b was directed by AAV2 transgene and was limited to the liver, EGFP mRNA was measured by qRT-PCR from RNA extracted from total liver, pancreas, heart, lung, kidney, ilium, and brain. No off-target expression of EGFP was identified in AAV2-miR19b-treated animals. EGFP was only detected in liver tissue from BDL AAV2-miR19b-treated rats and AAV2-control. However, no miR19b was associated with this increase in AAV2-control animals (Fig. 4B). Further confirmation of miR19b expression in HSCs was conducted by dual immunohistochemistry analysis for GFP (brown staining, marker for miR19b expression) and Desmin (purple staining, specific HSC marker). Sections were visualized and imaged by microscopy (40 × ) with a minimum of 10 fields per liver. No GFP-positive staining was detected in the livers of sham or BDL-NS rats. However, extensive GFP-positive staining was detected in HSCs, with the majority of the staining visualized along the portal tracts in AAV2-control miR and to a lesser degree in AAV2-miR19b animals. There was undetectable staining in hepatocytes (Fig. 4C).

BDL rats treated with AAV2-miR19b had increased expression of miR19b with decreased HSC activation.
To determine if AAV2-mediated expression of miR19b influenced the fibrotic response, mRNA for αSMA, a marker of HSC activation, and TGF-βRII, a key component of TGF-β signaling, were measured by qRT-PCR and protein by immunoblot. αSMA and TGF-βRII mRNA were significantly increased in all BDL groups compared to sham. However, mRNA for both pro-fibrotic markers were significantly decreased in BDL-AAV2-miR19b-treated rats compared to BDL-NS and BDL-AAV2-control (Fig. 5A). Immunoblots of αSMA and TGF-βRII confirmed protein expression was also significantly increased in BDL rats from NS and AAV2-control groups, with no significant difference identified in BDL AAV2-miR19b-treated rats when compared to sham animals (Fig. 5B). However, αSMA and TGF-βRII were significantly decreased in BDL AAV2-miR19b-treated rats compared to BDL-NS and to BDL AAV2-control.

Reintroduction of miR19b targeted to HSCs attenuated expression of mRNA and protein of pro-fibrotic markers.
Infiltrating macrophages and neutrophils were significantly reduced in AAV2-miR19b-treated BDL rats
In order to assess the impact of AAV2-mediated miR19b expression on immune cell infiltration in the BDL model, immunohistochemical analysis of tissue sections was performed with a primary antibody for macrophage marker CD68 followed by DAB chromagen and Leder stain for enzymatic detection of neutrophils. Counts are expressed as the mean number of macrophage or neutrophils present per field based on independent counts of five areas per slide by two independent investigators. Representative images show increased infiltration of both neutrophils (left panel) and macrophages (right panel) in BDL tissue compared to sham (Fig. 6A). Counts confirmed a significant increase in macrophages and neutrophils in all BDL tissue when compared to tissue from sham animals. Neutrophils (Fig. 6B) were significantly reduced in AAV2-miR19b-treated BDL animals by 51–53%, and macrophages (Fig. 6C) were significantly reduced by 29–35% compared to BDL-NS and BDL AAV2-control. Infiltrating immune cells (e.g., neutrophils and macrophages) play a significant role in the development and progression of fibrotic injury. Changes in fibrotic response by reintroduction of miR19b decreased tissue neutrophil and macrophage infiltration.
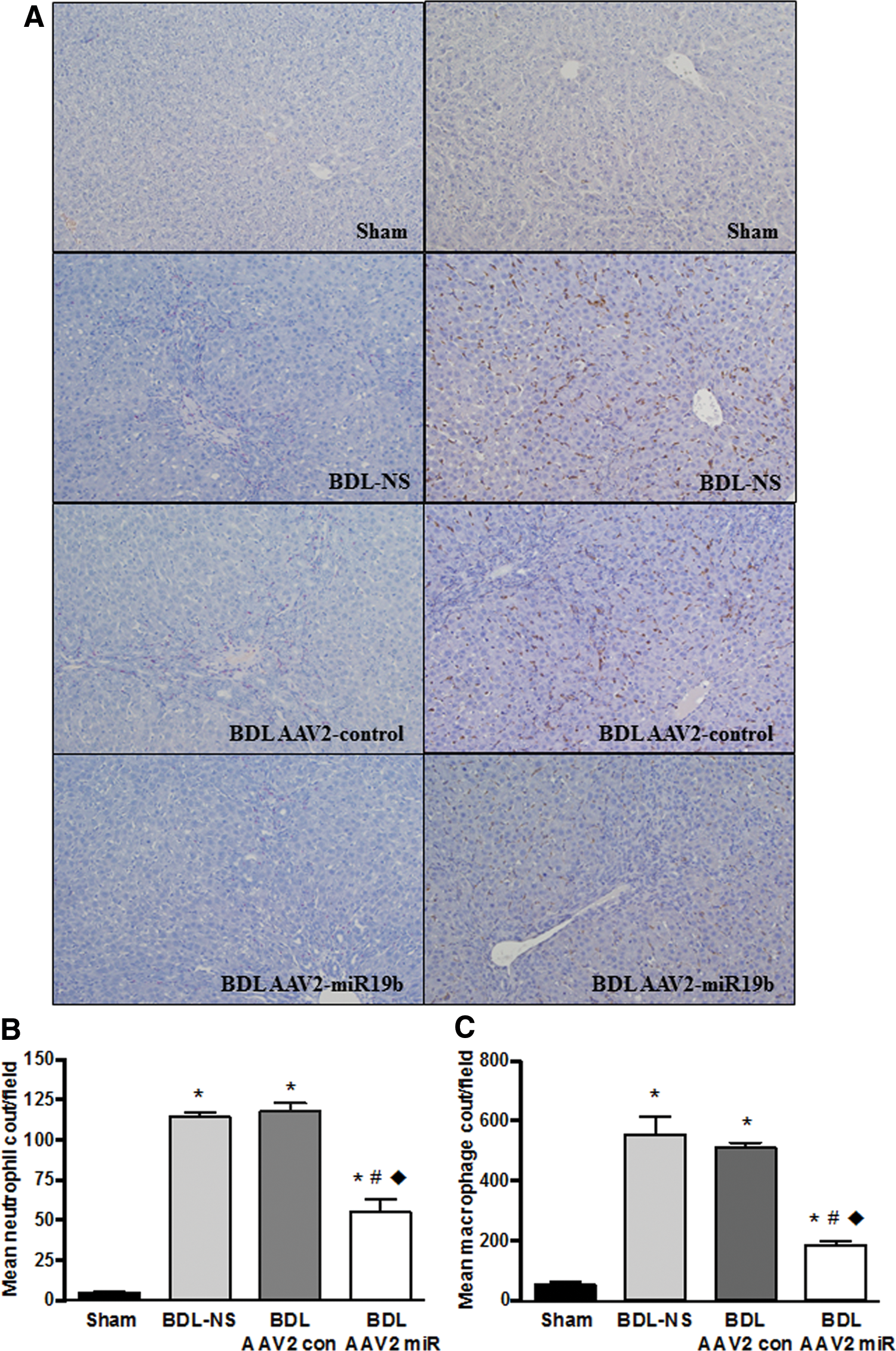
Infiltration of macrophages and neutrophils was inhibited in AAV2-miR19b treated rats.
Discussion
Fibrosis results from initial cell injury due to toxic, mechanical, or infective insult, which activates myofibroblast cells and inflammatory immune responses to promote wound healing. Loss of miR19b occurs in liver tissue from models of hepatic injury and is attributed to changes in HSC trans-differentiation. Previously, the authors' laboratory identified that miR19b was significantly downregulated during trans-differentiation of HSCs from a quiescent to an activated myofibroblast type cell, and cellular loss of miR19b on HSC activation enhanced TGF-β1 signaling through disruption of miR19b's inhibitory effects on TGF-βRII expression, and CTGF signaling during fibrosis in rodent models of hepatic injury and in human fibrosis. 13,14 This suggests miR19b replacement in activated fibroblasts may be a viable target to inhibit fibrotic injury. The present study focused on targeted re-expression of miR19b in activated HSCs to examine the effects on pro-fibrotic responses in vitro and in a rodent model of severe injury using BDL in vivo. HSC activation is a pivotal event during fibrogenesis. 11,12 TGF-β is a potent stimulator of collagen, and ECM synthesis in activated HSCs while promoting and sustaining HSC activation. 1,11 TGF-β is associated with major changes in protein coding mRNA and miR expression in HSCs that stimulates collagen deposition, inflammatory immune responses, and hepatic repopulation.
AAV2-mediated reintroduction of miR19b attenuated hepatic injury in vivo by inhibiting HSC activation. BDL produced severe mechanical injury in the model, with expected increases in markers of liver injury and decreases in bilirubin clearance, jaundice, and hepatomegaly. There was no significant difference in the body weight of BDL rats compared to those that underwent BDL with NS or AA2-control treatment. However, the body weights of rats in the BDL-AAV2-miR19b group were significantly reduced compared to sham animals. The liver governs energy metabolism and is metabolically connected to a variety of tissues. 25 To determine a possible explanation for this disparity, a role for miR19b in metabolic pathways was examined. miRs are critical regulators of protein coding mRNAs through promiscuous binding to the 3′-UTR. This regulatory mechanism also allows individual mRNA to interact with multiple miRs. A gene ontology analysis of miR19b identified >50 genes associated with metabolic processes that have binding sequences in the 3′-UTR, including acetyl-coA carboxylase (ACC1), a key regulator of both fatty acid oxidation and fatty acid synthesis. 25,26 In addition, long-chain fatty acids (LCFA) are activated and converted to LCFA-CoA by long-chain acyl-CoA synthetase (ACSL) ACSL1. The 3′-UTR of AAC1 and ACSL1 mRNA both contain consensus sequences for miR19b. Disruption of ACC enzymes is associated with increased leanness and decreased body weight in mice and has been reported to reduce hepatocyte lipid accumulation and insulin resistance in rats. 26 Disruption of ACC1 and/or ACSL1 in the liver of BDL rats treated with AAV2-miR19b may have produced alterations in lipid synthesis or oxidation pathways that resulted in impaired weight gain that remains to be further characterized.
HSCs are a direct target of mediators that activate nuclear factor kappa-light-chain enhancer of activated B cells, regulating several key factors in HSC biology, including activation, survival, and inflammatory responses, thereby contributing to a cycle of injury, inflammation, collagen deposition, and hepatic repopulation. 27 Inhibition of HSC activation can exert direct effects on the fibrogenic response, but also inhibit chemotactic factors involved in recruitment of macrophages and other immune cells. 28,29 Neutrophils enhance tissue damage, accumulating in sinusoids and sinusoidal venules during hepatic injury. Chemotactic signals from parenchymal cells promote neutrophil migration and adherence to hepatocytes where they can cause cell injury through formation of reactive oxygen species, releasing proteases into the hepatic milieu, which perpetuates inflammatory and wound repair responses. 30 Reintroduction of miR19b targeted to HSCs reduced these signals, resulting in impaired recruitment of neutrophils but also inhibited expansion of the Kupffer cell population. In the present model, rats receiving NS or AAV2 had higher infiltration of these inflammatory immune cell populations in response to BDL injury.
The burden of fibrosis and cirrhosis on the healthcare system is burgeoning, with cirrhosis claiming more than one million lives worldwide in 2013. 3 Clinically, fibrosis remains one of the only diseases for which there are no approved effective therapeutics. The economic burden for healthcare, diminished quality of life, and loss of workforce productivity remain staggering. 31 The molecular pathogenesis of fibrosis/cirrhosis involves multiple cell types responding to parenchymal damage, collagen and ECM deposition, inflammation, and oxidative stress. In the liver, activated HSCs are the dominate cell type responsible for collagen and ECM accumulation that leads to derangement of the hepatic architecture and scarring characteristic of cirrhosis, which has made them an attractive target for therapeutic intervention. 32,33 TGF-β is closely associated as a key signaling pathway in fibrotic injury. 4 In this model of severe mechanical injury, AAV2-miR19b was injected concurrent with BDL surgery to establish vector expression as the injury rapidly unfolded in response to TGF-β signaling. In treating a chronic injury model, tail injection 1–2 weeks after toxic insult should produce similar inhibitory effects on HSC activation and fibrogenic response and remains to be tested. Dual immunohistochemistry demonstrated miR19b expression in HSCs in AAV2-miR19b and AAV2-control-treated animals, although fewer GFP-positive activated HSCs were observed in AAV2-miR19b-treated BDL rats. The reduction of GFP-positive HSCs is due to the anti-fibrogenic functional effects of the AAV2-miR19b (i.e., miR19b is reducing HSC activation with concomitant decreases in miR19b expression due to decreased collagen promoter activity in the AAV2 vector). AAV2-miR expression in other hepatic cells, including endothelial cells, Kupffer cells, and hepatocytes was not test for experimentally. However, expression in these cells is not likely, as previous studies have shown AAV2 preferential infection/transgene expression in stellate cells with little detection in endothelial cells and hepatocytes. 21 Furthermore, even if other hepatic cells are infected by AAV2, transgene expression will only occur in collagen-producing cells, as the expression is driven by the collagen promoter. Reintroduction of miR19b in a rodent model of severe hepatic injury attenuated TGF-β signaling, HSC activation, hepatic remodeling, and collagen deposition. Concomitant with decreases in injury markers were improvement in hepatic function. However, metabolic changes may result in impaired weight gain and energy metabolism. AAV2-mediated reintroduction of miR19b targeted to HSCs appears to offer promise as a treatment for fibrotic pathologies (e.g., cardiac, renal, pulmonary) responsive to TGF-β signaling. Although the study herein focuses on miR19b as impeding the development of fibrosis, future studies should also evaluate the efficacy of miR19b therapy on other models of liver injury (e.g., alcohol, obesity), as well as determining if miR19b therapy could be effective in reversing fibrosis. The activated myofibroblasts are crucial to the development of fibrosis, making them an attractive target for therapeutic intervention. 32,33
Minor changes in the relative abundance of a few miRs can have profound effects on cellular physiology and pathological responses. 14 Within the last several years, there has been increasing interest within the pharmaceutical and biotechnology field to develop miRs into effective therapies either by reintroducing a missing miR or blocking the activity of an overexpressed miR. To date, most studies have been conducted in vitro, with overexpression achieved by transducing HSCs in culture. 13,34,35 Herein, targeting miR19b to HSC activity in vivo has been addressed. Successful use of miRs as therapeutics requires that they be delivered to a targeted cell type or tissue, transit the cell membrane, and become functionally active without introducing supra-physiological effects. Achieving a therapeutic response requires that it remain stable, evading endonuclease activity, immune surveillance, and phagocytosis. Finally, toxicity must be minimized and off-target effects characterized. 36 The fact that a single miR has the potential to regulate expression of many genes highlights the need for specificity. AAVs are a diverse collection of non-pathogenic ssDNA parvoviruses, which represent an attractive option for delivery of transgenes due to their persistence in tissues and lack of pathogenicity, which are currently being evaluated in Phase I/II clinical trials for a variety of diseases. 37,38 This study demonstrates that targeted delivery for re-expression of miR19b to HSCs by use of an AAV2 vector successfully inhibits fibrosis in a rodent model of severe injury.
Footnotes
Acknowledgments
The authors wish to thank Ian Hines, East Carolina University, for kindly providing the murine collagen promotor used to create the AAV2 vectors. The authors also wish to extend their gratitude to Carolina College of Health Sciences Histotechnology Program, class of 2016, Catrina McMaster, Taylor Short, Paige Hornsby, and Christine Cunningham, with Education Coordinator Tracy Walling BS, HT (ASCP). This work was supported by NIH AA022702.
Author Disclosure
The authors have nothing to disclose.