
Abstract
SNORD44 is a C/D box small nucleolar RNA, and exhibits low expression in breast cancer and head and neck squamous cell carcinoma tissues. Its host gene is growth arrest specific transcript 5 (GAS5), which is a long noncoding RNA. GAS5 is downregulated in colorectal cancer (CRC), and overexpression of GAS5 suppresses cell proliferation. However, the function of SNORD44 in CRC remains largely unknown, and the application of SNORD44 combined with GAS5 in CRC treatment has not been reported. In this study, the expression levels of SNORD44 and GAS5 were measured in CRC tissues by quantitative RT-PCR. The correlation between SNORD44 and GAS5 was evaluated by Pearson correlation analysis. An oncolytic adenovirus expressing SNORD44 and GAS5 (SPDD-UG) was constructed. The biological effects of SPDD-UG were investigated in CRC cell line SW620 and LS174T in vitro and in xenografts. The synergistic effect of rapamycin and SPDD-UG was explored in SW620 and LS174T cells and tumors. We demonstrated that SNORD44 expression level was markedly decreased in CRC tissues and positively correlated with GAS5 expression. SPDD-UG significantly inhibited SW620 and LS174T cell growth and induced cell apoptosis. Intratumoral injection of SPDD-UG significantly suppressed xenografts growth in nude mice. Moreover, the mechanistic target of rapamycin (mTOR) inhibitor, rapamycin, enhanced the antitumor effect through antagonizing the PI3K/Akt pathway activated by SPDD-UG. These results suggest that overexpression of SNORD44 and GAS5 by oncolytic adenovirus provides a promising method for CRC therapy.
Introduction
C
Small nucleolar RNAs (snoRNAs) are metabolically stable RNAs with 60 to 300 nucleotides located in the nucleus 4 and are mainly classified into two families, C/D box snoRNAs and H/ACA box snoRNAs, based on short conserved sequence elements. 5 The 2′-O-methylation guide C/D box snoRNAs contain conserved box C (RUGAUGA) and D (CUGA) motifs. The pseudouridylation guide H/ACA box snoRNAs carry box H (ANANNA) and ACA elements. 6,7 SNORD44 is a C/D box snoRNA, and is usually used as a reference gene in microRNA quantitative real-time PCR (qRT-PCR) detection. 8,9 Recent studies have shown that SNORD44 is downregulated in breast cancer and head and neck squamous cell carcinoma (HNSCC) tissues, and correlated with poor prognosis. 10,11 Additionally, the expression of SNORD44 positively correlated with the expression of its host gene, growth arrest specific transcript 5 gene (GAS5), in HNSCC. 11
GAS5 locates at chromosome 1q25 and is upregulated during cell growth arrest induced by serum starvation or the lack of growth factors. 12,13 Many studies have shown that GAS5 is downregulated in breast cancer, prostate cancer and lung cancer, and correlated with poor prognosis. 14 –16 Yin et al. verified that the lower expression of GAS5 was significantly correlated with larger CRC tumor volume, and overexpression of GAS5 inhibited CRC cell proliferation both in vitro and in vivo. 17
Based on the correlation between the expression of SNORD44 and GAS5 in HNSCC and the tumor suppressor role of GAS5 in CRC, we interrogate whether the expression of SNORD44 and GAS5 are also correlated in CRC samples and whether the combination of them could achieve anti-CRC effect.
Oncolytic adenoviral vector has been widely used in cancer therapy. 18,19 SPDD (survivin promoter-E1A 24-bp deletion-E1B deletion) is an oncolytic adenoviral vector. Its E1A gene is controlled by the promoter of survivin, which is a member of the inhibitor of apoptosis family and is transcriptionally upregulated in CRC. 20,21 The Rb binding domains (924–947 bp) in the E1A region and the entire E1B gene are deleted (Fig. 1A). SPDD displayed potent replication capacity and cytotoxicity in cancer cells, along with low toxicity against normal cells. 22
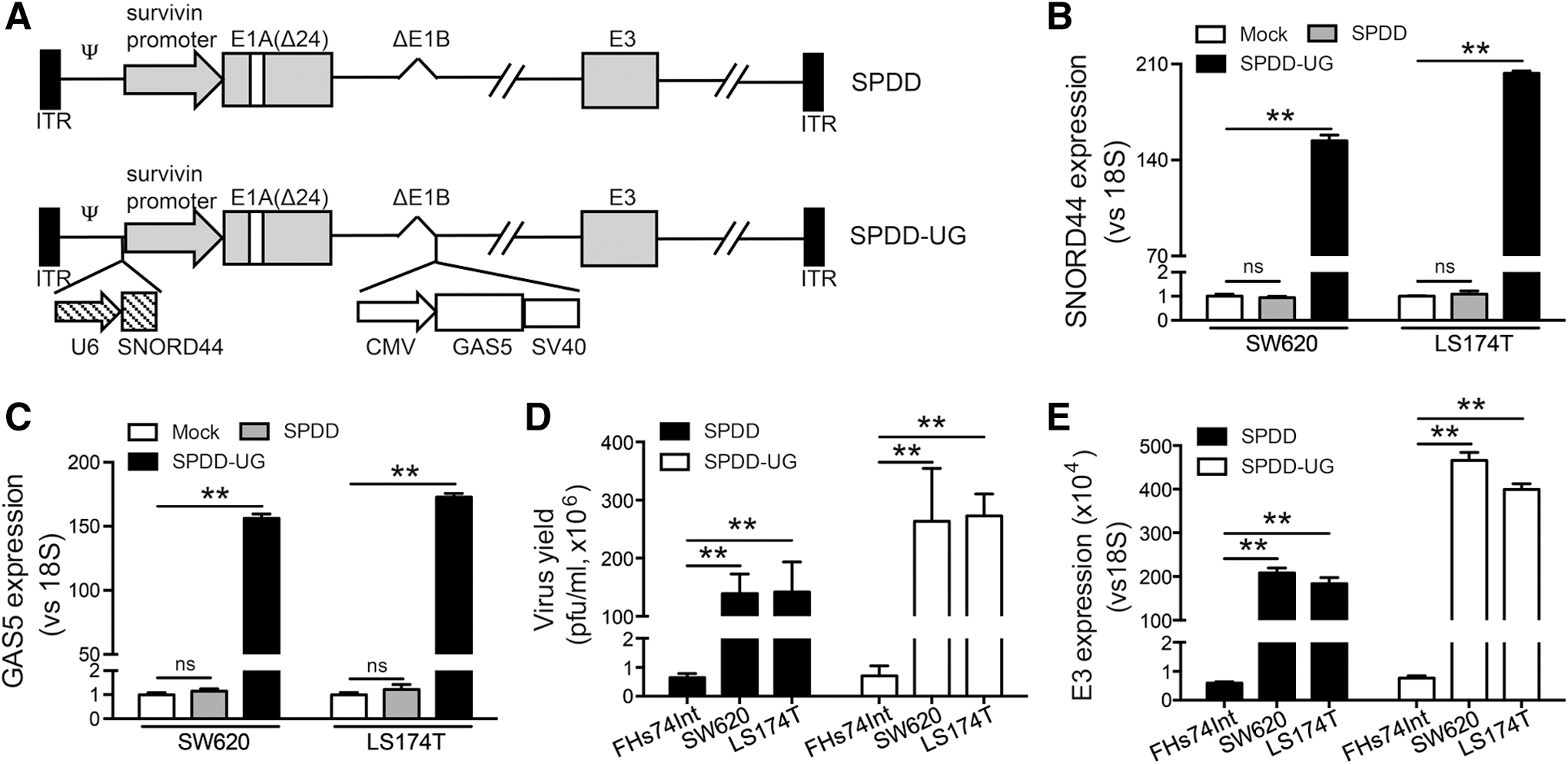
C/D box small nucleolar RNA SNORD44 and GAS5 overexpression mediated by oncolytic adenoviruses.
In this study, we found that the expression of SNORD44 and GAS5 is lower in CRC tissues than in the adjacent non-cancerous tissues. Overexpression of SNORD44 and GAS5 by SPDD (SPDD-UG) significantly inhibited CRC cell proliferation and induced caspase-dependent cell apoptosis both in cell lines and in tumor-bearing nude mice. Moreover, the mechanistic target of rapamycin (mTOR) inhibitor, rapamycin, suppressed the activation of the phosphatidylinositol 3-kinase/protein kinase B (PI3K/Akt) pathway arose by adenovirus infection, and enhanced the antitumor effect of SPDD-UG.
Materials and Methods
Patient samples
The frozen CRC patient samples and normal tissues were obtained anonymously from Hepatic Surgery Center, Tongji Hospital (Wuhan, China) with approval of the local ethics committees and written consent of all patients. Studies upon these samples were approved and handled in accordance with the institutional review board of Tongji Medical College.
Cell culture
Human colon carcinoma cell lines SW620 and LS174T, human small intestine epithelial cell line FHs74Int, and human embryonic kidney cell line HEK-293T were obtained from the Cell Bank of the Type Culture Collection of the Chinese Academy of Sciences (Shanghai, China). HEK-293 cell line was obtained from Microbix Biosystems Inc. (Toronto, Canada). All cells were cultured according to the instructions of the corresponding providers.
Lentivirus construction
Full-length SNORD44 and GAS5 cDNA were amplified by PCR and cloned into the lentiviral vector pLVX (Addgene, Cambridge, MA), respectively. The lentiviral stock was produced in 293T cells by transfecting pLVX-SNORD44/GAS5 and the lentivirus backbone plasmids pMD2.G and psPAX2 (Addgene). Cells were infected with the lentivirus and selected with puromycin (10 μg/mL).
Adenovirus construction and identification
Oncolytic adenovirus was packaged and amplified in HEK-293 cells and purified by gradient CsCl solution centrifugation. Virus titer was measured by Quick Titer Adenovirus Titer Immunoassay Kit (Cell Biolabs, San Diego, CA) according to the manufacturer's instructions. Adenovirus genomic DNA was extracted and applied for PCR using the following primers: survivin promoter, forward 5′-cgcgttctttgaaagcagtcg-3′ and reverse 5′-tgccgccgccgccacctctgc-3′; E1A region, forward 5′-ttacctgccacgaggctgctttc-3′ and reverse 5′-ttatggcctggggcgtttac-3′; E1B region, forward 5′-gtaaacgccccaggccataa-3′ and reverse 5′-tcaatctgtatcttcatcg-3′.
Quantitative RT-PCR assay
Total RNA was isolated from cells using TRIZOL reagent (Invitrogen, Carlsbad, CA). Single-strand cDNA for PCR template was synthesized using ReverTra Ace qPCR RT Kit (Toyobo, Osaka, Japan), and SuperReal Premix plus (Vazyme Biotech, Nanjing, China) was used for amplification according to the corresponding protocols. The primers sequences are: SNORD44, forward 5′-cctggatgatgataagcaaatg-3′ and reverse 5′-gtcagttagagctaattaagacc-3′; GAS5, forward 5′-cttctgggctcaagtgatcct-3′ and reverse 5′-ttgtgccatgagactccatcag-3′; E3, forward 5′-gctaccggacttacatctacc-3′ and reverse 5′-agggatatatagctagggctaaag-3′; 18S forward, 5′-aactttcgatggtagtcgcc-3′ and reverse 5′-ccctggatgtggtagccgttt-3′.
Western blot
Protein concentrations were estimated using the Lowry assay (BioRad, Hercules, CA). Western blot was performed using standard protocols. Primary antibodies against pro-caspase 3, pro-caspase 8, pro-caspase 9, poly (ADP-ribose) polymerase (PARP) (Santa Cruz biotechnology, Santa Cruz, CA), Akt, phospho-Akt, mTOR, phospho-mTOR, p70S6K, phospho-p70S6K (Cell Signaling Technology, Danvers, MA), and glyceraldehyde 3-phosphate dehydrogenase (CWBIO, Beijing, China) were used. All the horseradish peroxidase-conjugated secondary antibodies were purchased from Santa Cruz biotechnology.
Cell proliferation assay
Cell proliferation was measured using the CCK-8 kit (Vazyme Biotech). Cells were incubated with 10 μL CCK-8 regent at 37°C for 4 h. Absorbance was read at a wavelength of 450 nm using Biotek Eon Microplate Reader (Biotek Instruments, Winooski, VT). The coefficient of drug interaction (CDI) was used to analyze the synergistically inhibitory effect of SPDD-UG and rapamycin combinations. CDI is calculated as follows: CDI = AB/(AXB). According to the absorbance of each group, AB is the ratio of the combination groups to the control group, and A or B is the ratio of the single agent groups to the control group. Thus a CDI value less than, equal to, or greater than 1 indicates that the drugs are synergistic, additive, or antagonistic, respectively. CDI less than 0.7 indicate that the drugs are significantly synergistic. 23
Colony formation assay
Two thousand cells per well were seeded in six-well plates. Ten days later, cells were stained using crystal violet, photographed, and applied to statistical analysis.
Flow cytometry analysis
Apoptosis was analyzed using the Annexin V-FITC/PI Apoptosis Detection kit (BD Biosciences, SanJose, CA). Detection of cell proliferation was performed using Ki-67 staining kit (BD Biosciences) according to the instructions of the corresponding providers.
Animal experiments
All in vivo experiments were performed according to approved protocols from the U.S. Public Health Service Policy on Humane Care and the Use of Laboratory. One million SW620 or LS174T cells were subcutaneously injected into the right flank of each nude mouse. When tumors reached 80–120 mm3, either oncolytic adenoviruses (5 × 108 PFU per mouse) or PBS was injected into tumors every other day for twice. For rapamycin treatment, mice were treated daily with rapamycin (10 mg/kg) by intraperitoneal injection for 10 days. The tumor volume was measured by a vernier caliper every other day and calculated as (length × width 2 )/2. At the end of the experiment, tumors were resected from the sacrificed mice for further studies. Aspartate transaminase and alanine transaminase levels in serum were measured.
Immunohistochemistry and TUNEL Assay
Tumor tissues were fixed in 4% formaldehyde overnight and embedded in paraffin. Five-micrometer-thick sections were cut and subjected to immunohistochemistry and TUNEL assay. To detect the Ki-67 and hexon expression, sections were incubated with Ki-67 antibody (Cell Signaling Technology), hexon antibody (Abcam, Cambridge, MA) or p-Akt antibody (Cell Signaling Technology) with 1:100 dilution. The TUNEL assay was performed using the in situ cell death detection kit (Roche Systems, Basel, Switzerland) according to the manufacturer's instructions. Hematoxylin was used as counterstain.
Hemotoxylin and eosin staining
For hemotoxylin and eosin staining, tumor tissues were treated as above, and 5-μm-thick sections were stained by hematoxylin and eosin.
Statistical analysis
Data are presented as the mean ± standard deviation. Mean values were compared using an unpaired Student's two-tailed t-tests; p < 0.05 was defined as statistically significant, and p < 0.01 was defined as extremely statistically significant.
Results
The expression of SNORD44 is positively correlated with GAS5 in CRC samples
SNORD44 is a C/D box snoRNA, and is located within extron 5–6 of GAS5 gene (Supplementary Fig. S1; Supplementary Data are available online at

SNORD44 and GAS5 express low in colorectal cancer (CRC) tissues and inhibit the CRC cell growth and colony formation.
To investigate the function of SNORD44 and GAS5 in CRC cells, stable CRC cell lines SW620 and LS174T overexpressing SNORD44, GAS5, and the combination of SNORD44 and GAS5 were established (Fig. 2D, E). Individual overexpression of SNORD44 or GAS5 suppressed cell growth. The combination had stronger inhibit effect (Fig. 2F, G). Additionally, the SNORD44 and GAS5-coexpressed cells showed a significantly decreased ability of colony formation (Fig. 2H, I). We also noticed that upregulation of SNORD44 did not regulate the expression of GAS5 gene, and vice versa (Supplementary Fig. S2).
Oncolytic adenovirus efficiently overexpresses SNORD44 and GAS5 in CRC cells
Since the combination of SNORD44 and GAS5 displayed better antitumor effect, and oncolytic adenovirus could replicate in tumor cells and had potential clinical usage ability, we constructed an oncolytic adenovirus carrying SNORD44 and GAS5 gene (SPDD-UG) (Fig. 1A). SNORD44 and GAS5 expression was controlled by U6 and CMV promoter, respectively. The SNORD44 expression cassette was cloned into the region ahead of E1A. The GAS5 expression cassette was inserted into the deleted E1B region. PCR analysis confirmed that all the oncolytic adenoviruses were free of wild type adenovirus contamination (Supplementary Fig. S3). The overexpression of SNORD44 and GAS5 in SW620 and LS174T cells was confirmed by qRT-PCR (Fig. 1B, C). We next examined whether the insertion of transgenes altered the capacity of viral replication using the viral progeny assay and qRT-PCR assay with primers targeting the adenoviral E3 region. The SPDD-UG production was not impaired compared with SPDD control virus in SW620 and LS174T cells, indicating that transgenes did not interfere with adenoviral replication. Additionally, oncolytic adenoviruses presented higher production in CRC cells than in the normal intestinal epithelial FHs74Int cells (Fig. 1D, E).
SPDD-UG inhibits CRC cell growth and induces cell apoptosis
SPDD-UG inhibited SW620 and LS174T cell growth at a series of multiplicity of infections (5, 10, 20) 4 days after infection (Fig. 3A, B). SPDD-UG also suppressed cell growth at a multiplicity of infection of 10 in a time-dependent manner (Fig. 3D, E). Importantly, SPDD-UG did not affect the growth of FHs74Int cells (Fig. 3C, F), the same as the control oncolytic adenovirus SPDD. Ki-67 is associated with tumor aggressiveness or progression in numerous malignancies. 25 SPDD-UG treatment reduced the Ki-67 expression in SW620 and LS174T cells, indicating that the cell proliferation was decreased (Fig. 3G, H). Annexin V/PI staining disclosed that SPDD-UG treatment induced apoptosis in SW620 and LS174T cells (Fig. 3I, J). Moreover, decreased protein level of pro-caspase 3/8/9 and increased cleavage form of PARP protein were observed 2 days after SPDD-UG infection, suggesting that SPDD-UG induced CRC cell apoptosis via caspase-dependent pathway (Fig. 3K).
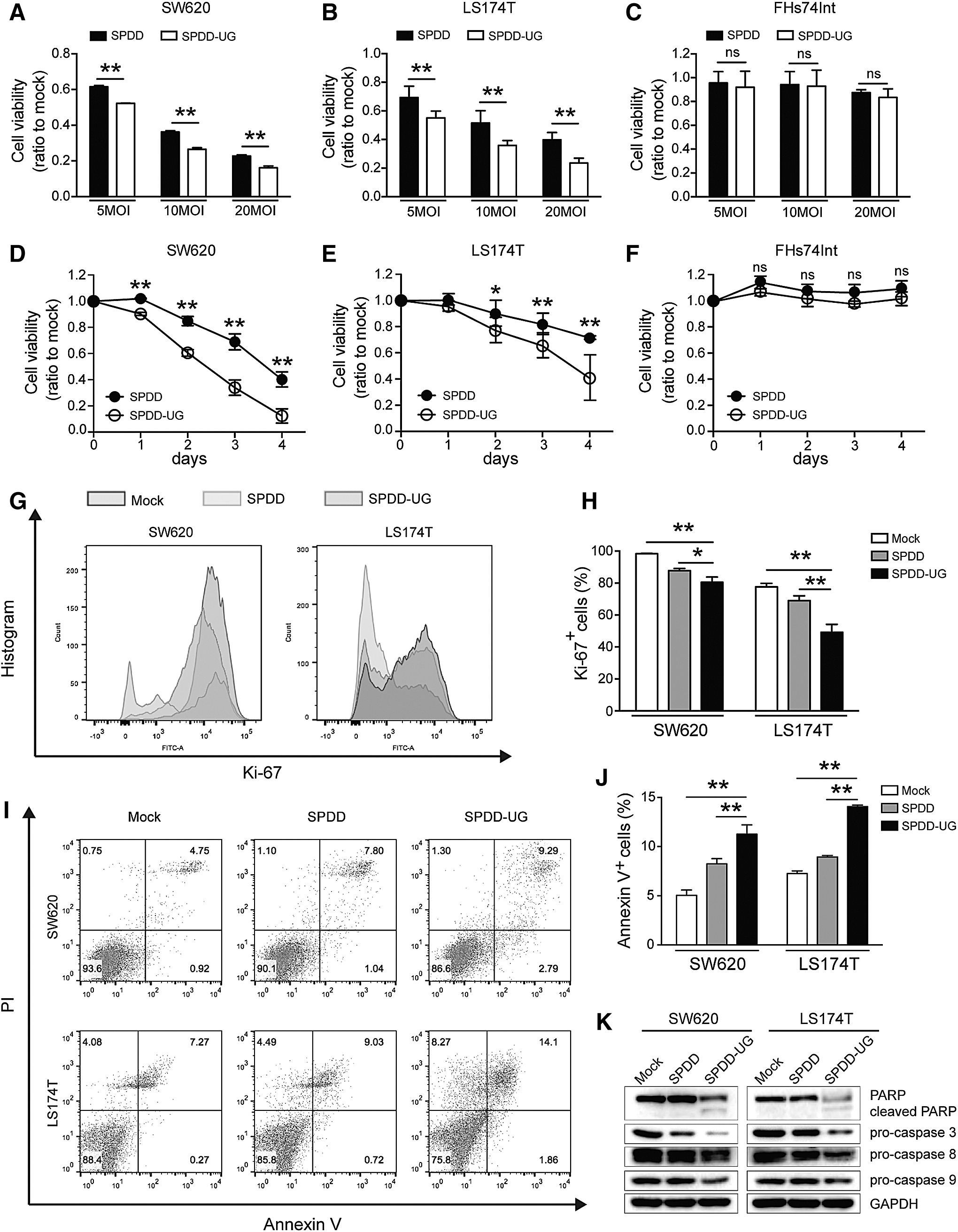
SPDD-UG inhibits CRC cell growth and induces apoptosis.
SPDD-UG reduces growth of CRC tumors in mice
Next, we sought to identify the antitumor effect of SPDD-UG in CRC xenograft models. Subcutaneous SW620 and LS174T xenografts in nude mice were established. SPDD-UG treated groups presented a remarkable reduction in tumor size and weight compared with SPDD or PBS-treated tumors (Fig. 4A–D). qRT-PCR analysis showed that SPDD-UG efficiently expressed SNORD44 and GAS5 in tumors 24 days after virus injection (Fig. 4E, F). The existence of oncolytic adenovirus in the tumor tissues was examined by immunohistochemical staining of adenovirus capsid protein hexon. SPDD and SPDD-UG treated tumors presented significant positive hexon staining (Fig. 4G–I). The SPDD-UG treated tumors displayed reduction of Ki-67 staining (Fig. 4G, H, J), indicating that SPDD-UG was competent to suppress SW620 and LS174T cell proliferation in vivo. TUNEL staining analysis revealed that SPDD-UG increased apoptosis in tumors (Fig. 4G, H, K).
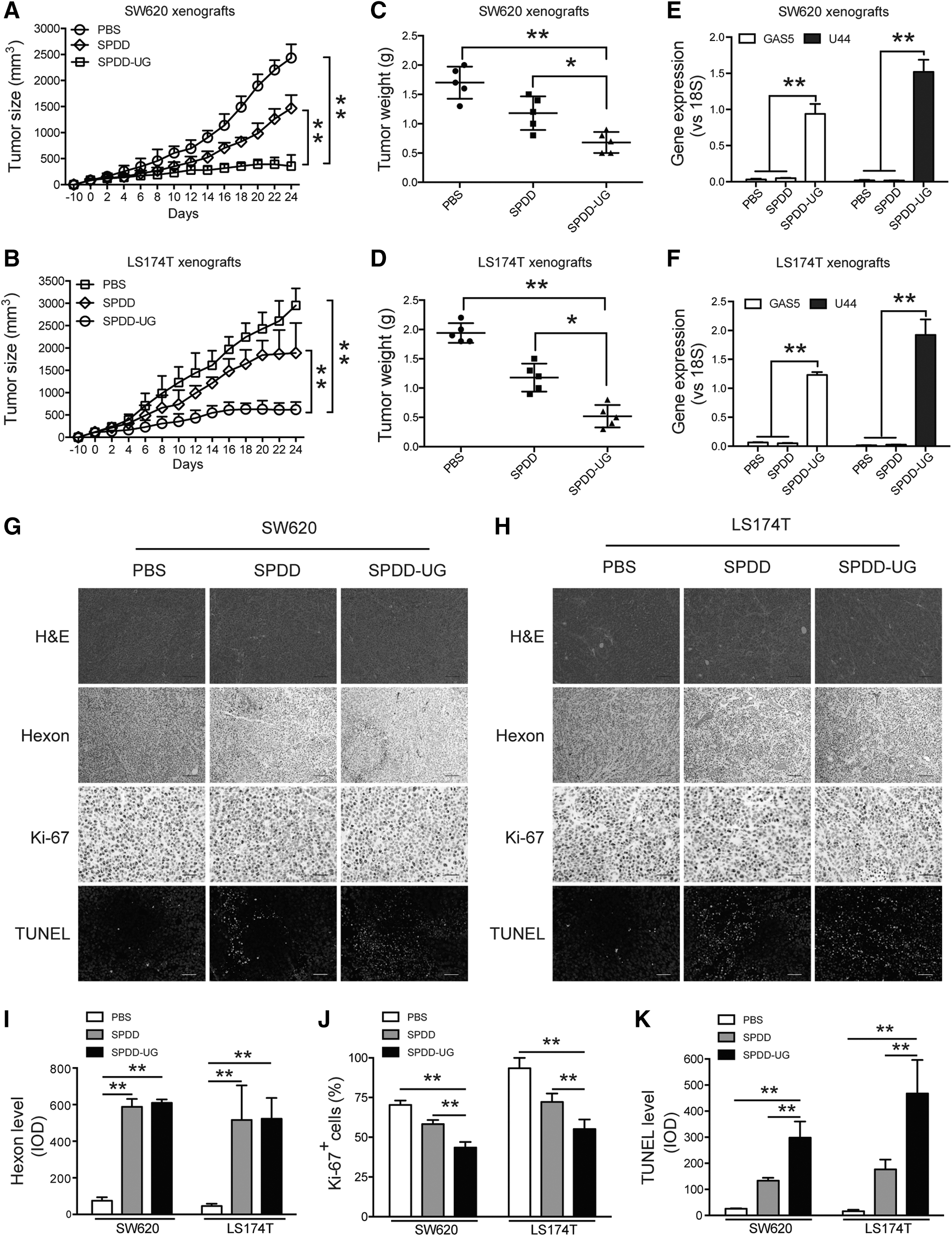
SPDD-UG inhibits CRC cell growth and induces apoptosis in vivo.
Rapamycin and SPDD-UG synergistically inhibit CRC cell proliferation
Although SPDD-UG effectively inhibited tumor growth, the size of those tumors still stayed around 100–500 mm3. We are wondering whether SPDD-UG induces some molecular factors that weaken its antitumor effect. Previous studies have shown that adenovirus infection could activate PI3K/Akt signaling pathway, which may benefit tumor cell survival. 26,27 We noticed that SPDD and SPDD-UG, especial SPDD-UG, distinctly increased the phosphorylation of Akt at Ser473 and mTOR at Ser2448. The phosphorylation of p70S6K at Thr389 was also obviously elevated by SPDD-UG (Fig. 5A, B). It was reported that GAS5 lncRNA sensitizes prostate cancer cells and breast cancer cells to mTOR inhibitors. 28,29 We found that the mTOR inhibitor rapamycin antagonized the activation of PI3K/Akt pathway arised by SPDD-UG (Fig. 5C). To test whether rapamycin could potentiate the SPDD-UG-mediated tumor inhibition, we combined SPDD-UG with rapamycin. SW620 and LS174T cells were treated with serious dose of SPDD-UG or/and rapamycin. Results showed that rapamycin further enhanced the efficacy of SPDD-UG in a synergistic manner (Fig. 5D–F).

Rapamycin inhibits the SPDD-UG-induced activation of PI3K/Akt pathway and enhances cytotoxicity of SPDD-UG.
Rapamycin enhances the tumor suppression effect of SPDD-UG in the CRC xenografts
Furthermore, we validated the therapeutic effect of SPDD-UG in combination with rapamycin in CRC xenograft models. Results showed that the growth of tumors was slower in the SPDD-UG and rapamycin co-treated mice than in the SPDD-UG-treated mice (Fig. 6A, B). No increase in aspartate transaminase and alanine transaminase levels after treatment was observed (Supplementary Fig. S4). Mice were sacrificed 24 days after treatment, and tumors were weighed. Tumors from the co-treated mice were lighter than those from the other groups (Fig. 6C, D). Additionally, the average body weight of the co-treated mice was heavier than the other mice (Fig. 6E, F). IHC analysis on tumor sections showed that rapamycin antagonized the phosphorylation of Akt at Ser473 (Fig. 6G–I). The co-treated tumors displayed enhanced reduction of Ki-67 staining (Fig. 6G, H, J). TUNEL analysis revealed that the co-treated tumors contained much more apoptotic cells than those from the other groups (Fig. 6G, H, K).
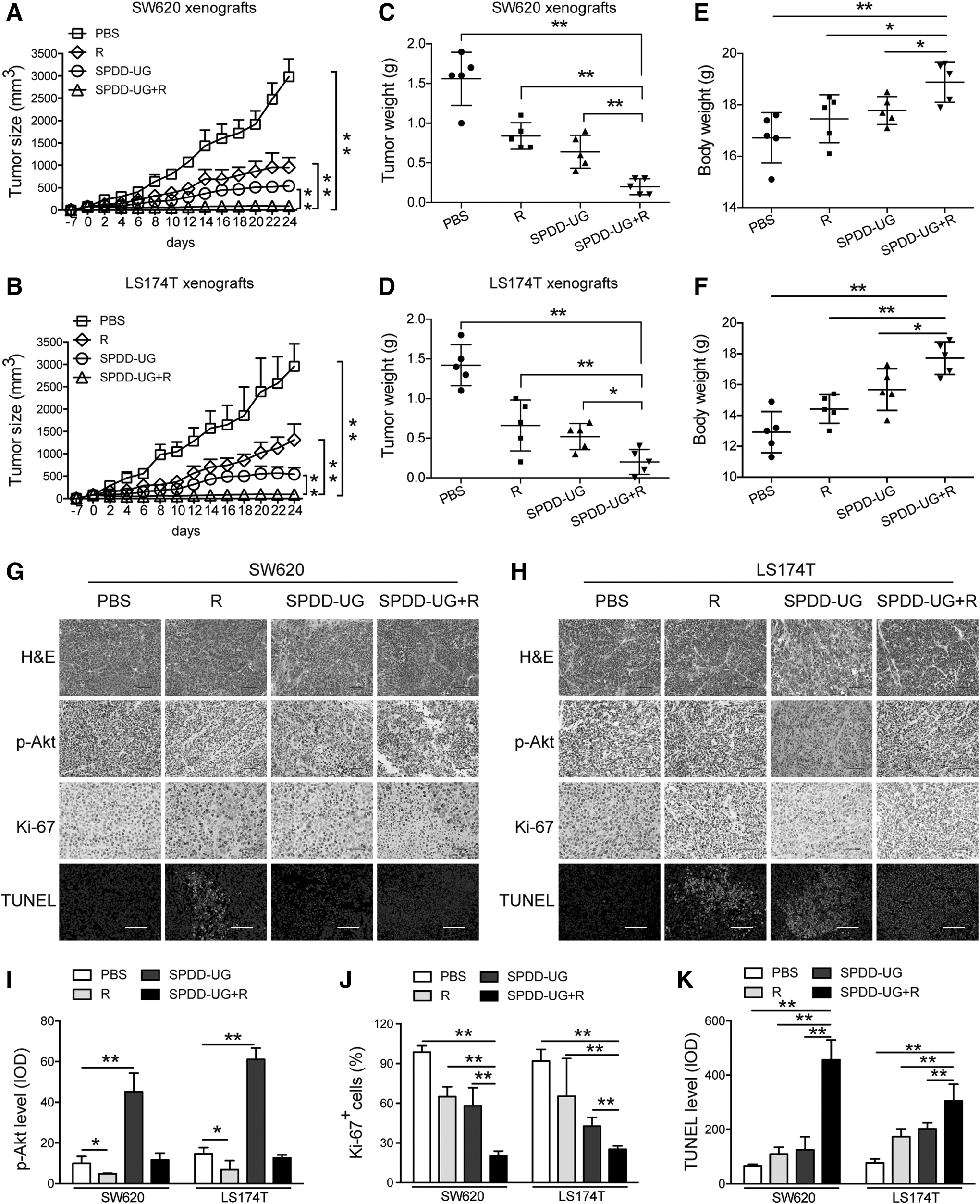
Combination of SPDD-UG and rapamycin significantly inhibits the growth of CRC cells in vivo.
Discussion
Oncolytic virus has the special property with selective infection of tumor cells, and thus lead to oncolysis of cancer cells and minimal toxicity to normal tissues. A great variety of oncolytic viruses have shown promise in preclinical models and clinical studies. 30 Human adenoviruses are responsible for respiratory, gastroenteric, and ocular infections and can serve as vectors for gene therapy. 31 Oncolytic adenovirus carrying two therapeutic genes, such as TRAIL gene and Smac gene or TRAIL gene and short hairpin RNA against COX-2, possessed better antitumor effect than one gene. 32,33
SNORD44 is usually used as a reference gene in qRT-PCR tests; however, recent studies have shown that SNORD44 is significantly associated with breast cancer and HNSCC prognosis. 10,11 In the present study, we found that SNORD44 was downregulated in CRC tissues (Fig. 2A), and overexpression of SNORD44 inhibited CRC cell growth (Fig. 1F–I), indicating that SNORD44 might function as a tumor suppressor in CRC. The long noncoding RNA GAS5 is the host gene of SNORD44. 24 Our studies are consistent with Yin et al.'s work that overexpression of GAS5 efficiently suppresses CRC cell proliferation (Fig. 2F–I). 17
Oncolytic adenoviral vector SPDD has selective replicating ability and potential cytotoxicity in cancer cells. 22 So far, few research using oncolytic adenoviral vector delivering snoRNA and its host gene for cancer therapy has been reported. We cloned SNORD44 and GAS5 gene into SPDD vector, and demonstrated that SPDD-UG presented significantly cytotoxicity to CRC cells in vitro and in vivo (Figs. 3 and 4).
The PI3K/Akt signaling pathway is one of the major dysregulated signaling pathways in cancer. 34,35 It is involved in diverse cellular processes such as cell growth and proliferation. 36,37 It is reported that the PI3K/Akt pathway can be stimulated by adenovirus infection. 26,27 We found that the SPDD-UG–infected SW620 and LS174T cells had an increased phosphorylation of Akt, mTOR, and p70S6K (Fig. 5A, B), indicating that SPDD-UG activated the PI3K/Akt signaling pathway. Rapamycin was first used to prevent organ rejection in kidney transplant patients. 38 Nowadays, it is reported that rapamycin could suppress the proliferation of colon cancer cells in culture, and promote apoptosis by inhibiting the phosphorylation of Akt and p70S6K. 39,40 Therefore, we explored the combined effect of SPDD-UG and rapamycin. Results revealed that rapamycin antagonized the phosphorylation of Akt, mTOR and p70S6K (Fig. 5C), and improved the antitumor efficiency of SPDD-UG both in cell lines and in animal models (Figs. 5D–F and 6).
In summary, this study revealed a synergistic antitumor effect of rapamycin and oncolytic adenovirus expressing SNORD44 and its host gene GAS5, providing a promising treatment strategy for CRC. To our knowledge, this is the first report demonstrating that rapamycin synergizes with oncolytic adenovirus carrying snoRNA and host gene to inhibit CRC cell growth.
Footnotes
Acknowledgments
The authors thank animal care facility for excellent technical assistance. This work was supported by National Natural Science Fund (31671348) and Zhejiang Provincial Top Key Discipline of Biology Open Foundation (2016D10).
Author Disclosure
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.