
Abstract
Infantile malignant osteopetrosis (IMO) is a rare, lethal, autosomal recessive disorder characterized by nonfunctional osteoclasts. More than 50% of the patients have mutations in the TCIRG1 gene, encoding for a subunit of the osteoclast proton pump. The aim of this study was to develop a clinically applicable lentiviral vector expressing TCIRG1 to correct osteoclast function in IMO. Two mammalian promoters were compared: elongation factor 1α short (EFS) promoter and chimeric myeloid promoter (ChimP). EFS promoter was chosen for continued experiments, as it performed better. IMO osteoclasts corrected in vitro by a TCIRG1-expressing lentiviral vector driven by EFS (EFS-T) restored Ca2+ release to 92% and the levels of the bone degradation product CTX-I to 95% in the media compared to control osteoclasts. IMO CD34+ cells from five patients transduced with EFS-T were transplanted into NSG mice. Bone marrow was harvested 9–19 weeks after transplantation, and human CD34+ cells were selected, expanded, and seeded on bone slices. Vector-corrected IMO osteoclasts had completely restored Ca2+ release. CTX-I levels in the media were 33% compared to normal osteoclasts. Thus, in summary, evidence is provided that transduction of IMO CD34+ cells with the clinically applicable EFS-T vector leads to full rescue of osteoclasts in vitro and partial rescue of osteoclasts generated from NSG mice engrafting hematopoietic cells. This supports the continued clinical development of gene therapy for IMO.
Introduction
I
Previous studies have rescued the murine oc/oc disease model of osteopetrosis with gene therapy, utilizing a gammaretroviral vector to target hematopoietic stem cells. 11 It has also been demonstrated that the resorptive function of human IMO osteoclasts was restored in vitro by lentiviral-mediated gene transfer of TCIRG1 cDNA into CD34+ cells obtained from the peripheral blood of IMO patients followed by expansion of the cells in culture and differentiation on bone slices to mature bone-resorbing osteoclasts. 4 However, in that study, TCIRG1 was driven by the spleen focus forming virus (SFFV) promoter that raises safety concerns in terms of insertional deregulation of proto-oncogenes. 12 Instead of employing strong viral promoter/enhancer sequences, having led to severe adverse events in past gene therapy trials, 13,14 the use of weaker cellular promoters can increase treatment safety.
The current work thus modified the lentiviral vector for possible use in a clinical setting. The SFFV promoter was replaced with one of two different mammalian promoters, and these were then tested for correction of IMO osteoclast function in vitro. The correction at the level of putative stem cells was also assessed by transplantation of gene corrected CD34+ cells from IMO patients to NSG mice.
Materials and Methods
CD34+ cell isolation, culture, and expansion
Samples of peripheral blood from IMO patients (University Medical Center Ulm; Supplementary Table S1; Supplementary Data are available online at
Vectors, viral production, and transduction of CD34+ cells
All the vectors used in this study are self-inactivating (SIN) lentiviral vectors with a pRRL backbone (Supplementary Fig. S1). For comparing promoter experiments, three rescue vectors were used (SFFV-TG, EFS-TG, ChimP-TG), which contain the cDNA of human TCIRG1 under the spleen focus-forming virus (SFFV) promoter, the elongation factor 1α short (EFS) promoter, or the chimeric myeloid promoter (ChimP), 15 respectively, upstream of an internal ribosomal entry site (IRES), which is followed by the gene for enhanced green fluorescent protein (GFP) used as a marker gene. For subsequent in vitro studies and for transplantations, a vector expressing TCIRG1 alone under the EFS promoter (EFS-T) was used, without any marker gene. Control vectors expressed GFP under the SFFV or EFS promoters (SFFV-G, EFS-G). Lentiviral vectors were produced by transient transfection of the vector plasmids into 293T cells, along with packaging plasmid (pCMV ΔR8.91), and envelope plasmid (VSV-G pMDG). Transductions were carried out in 24-well plates coated with RetroNectin (Takara Bio, Otsu, Japan). For the in vitro experiments, CD34+ cells were transduced with a first hit at a multiplicity of infection (MOI) of 30 for 6 h on day 3 and a second hit at a MOI of 30 for 6 h on day 7 followed by a week of culture with a myeloid cytokine cocktail and subsequent differentiation to osteoclasts, as described above. For the in vivo experiments, a shorter transduction protocol was developed to allow efficient transduction while maintaining the stem/progenitor nature of the CD34+ population. Mononuclear cells were thawed, and CD34+ cells were isolated and transduced with the first hit (MOI of 30 or 100) overnight followed by transduction on the following day with a second hit (MOI of 30 or 100) for 6 h, after which the cells were transplanted in the NSG mice. The total culture time of the cells prior to transplantation was <30 h.
Osteoclastogenesis
After 2 weeks, the expanded cells were reseeded into 96-well plates on plastic or on bovine cortical bone slices at a density of 1 × 105/well for cell assays and 1.0 × 106/well on plastic in a 12-well plate for Western blot. The cells were incubated at 37°C and 5% CO2 in alpha minimum essential medium containing 10% heat-inactivated fetal bovine serum (FBS), 100 units/mL of penicillin, 100 μg/mL of streptomycin, and 388 μg/L of thymidine. They were expanded for 3 days in the presence of 50 ng/mL of M-CSF and were differentiated for an additional 10 days in the presence of 50 ng/mL of M-CSF and 50 ng/mL of receptive activator of nuclear factor kappa-B ligand (RANKL), both from R&D Systems, with medium changes every 2–3 days. After 13 days, the cells were either fixed in 4% formaldehyde for further analyses or lysed for Western blot analysis. Resorption was assessed by CTX-I and Ca2+ release into the media and the formation of resorption pits. Osteoclastogenesis was assessed by tartrate-resistant acid phosphatase (TRAP) activity in the media.
Western blot
Cells were harvested into radioimmunoprecipitation assay (RIPA) buffer. Protein concentrations were measured using a Protein Assay Kit II (Bio-Rad, Hercules, CA). Total protein (15 μg) in SDS sample buffer was separated by gel electrophoresis in an SDS-PAGE 4–12% gradient gel followed by blotting onto a nitrocellulose membrane. Membranes were then blocked in Tris-buffered saline (TBS) with Tween 20 (TBST) with 5% skim milk powder for 1 h at room temperature followed by incubation with a primary antibody overnight at 4°C in TBST with 5% skim milk powder using the following antibody dilutions: mouse monoclonal anti-TCIRG1 (catalog # H00010312-M01A; Abnova, Taipei, Taiwan) 1:1000 and rabbit polyclonal anti-p38 MAPK (catalog # 9212; Cell Signaling Technology, Danvers, MA) 1:1000. The blots were then washed for 3 × 10 min and incubated with the corresponding horseradish peroxidase–conjugated secondary antibody for 1 h at room temperature followed by 3 × 10 min washes in TBS. Blots were developed using ECL Western Blotting Reagents (GE Healthcare, Waukesha, WI). To estimate the relative TCIRG1 protein expression levels between vectors, a semi-quantitative analysis of the developed Western blot films was carried out using Fiji. 16 The films were digitized as PDF files using an office scanner, and converted into 8-bit black-and-white tiff images in Fiji. The bands of interest were marked using the rectangle tool, and the band intensity peaks were plotted using the gel analysis tool. Background signals in the plots were removed by separating the peaks from the background intensity using the straight-line tool, and the area of each isolated peak was subsequently measured. The TCIRG1 peak areas were divided by the corresponding p38 peak areas, and the TCIRG1/p38 peak area ratios were normalized to that of the CB EFS-T condition, due to one experiment lacking a CB EFS-G condition, of the respective experiment (n = 3).
TRAP activity measurements
Between 1 and 20 μL of media from 96-well cell cultures on either bone or plastic was added to a 96-well plate and diluted with water to a volume of 20 μL. The diluted samples were incubated with 80 μL of freshly prepared reaction buffer (0.25 M acetic acid, 0.125% Triton X-100, 0.25 M NaCl, 2.5 mM EDTA, 1.1 mg/mL of ascorbic acid, 5.75 mg/mL of disodium tartrate, 2.25 mg/mL of 4-nitrophenylphosphate, pH 5.5) at 37°C for 1 h in the dark, and the reaction was then stopped by adding 100 μL of 0.3 M NaOH. Absorbance was measured at 405 nm, with 650 nm as a reference using a SpectraMax M5 (Molecular Devices, Sunnyvale, CA) plate reader.
Resorption biomarkers
The release of the c-terminal type I collagen fragments (CTX-I) from resorbed bone slices was determined using the CrossLaps for Culture kit (IDS, Boldon Colliery, United Kingdom), which was used according to the manufacturer's instructions.
The release of Ca2+ was analyzed by measuring the concentration of total calcium in media after resorption using a colorimetric calcium (CPC) assay and an ADVIA 1800 Clinical Chemistry System (both from Siemens Healthineers, Erlangen, Germany).
Resorption pit formation
Resorption pits on the fixed bone slices were visualized by washing them with water, removing the remaining cells by lysing them with RIPA buffer, and scrubbing with a cotton swab followed by staining with hematoxylin for 7 min. Excess dye was removed by scrubbing the bones with a cotton swab. Digital micrographs were obtained using a 10 × objective and an Olympus DP71 digital camera mounted on an Olympus IX-70 microscope using the Cell-A software (Olympus, Center Valley, PA).
NSG mice and transplantations
Breeding pairs of immunodeficient NOD-scid IL2rγnull (NSG) mice were obtained from Charles River Laboratories (Sulzfeld, Germany). The mice were maintained in the conventional animal facility at the Biomedical Centre, Lund University. All experiments were performed according to protocols approved by the local animal ethics committee. NSG mice (8–15 weeks old) were sub-lethally irradiated with 300 cGy and transplanted 6 h later with 1 × 105 untransduced CB CD34+ cells or IMO CD34+ cells transduced with either EFS-T or EFS-G by tail-vein injection. The mice were administered ciprofloxacin via their drinking water for 2 weeks to avoid post-transplantation infections. Peripheral blood was harvested at different time points, and bone-marrow cells were harvested by crushing the femora with a mortar after termination of the mice.
Vector copy number
Vector copy number (VCN) analysis was performed on whole bone-marrow genomic DNA from samples harvested from mice 9–19 weeks after transplantation. The mean VCN per cell was determined by quantitative reverse transcription polymerase chain reaction. Samples were measured in triplicates using 100 ng of genomic DNA. Primers for the WPRE element of the vector were used to determine the amount of viral sequences, which was further normalized to a genomic reference sequence of the Ptbp2 gene. 17 A serial dilution of a plasmid standard containing both sequences was measured in parallel to perform an absolute quantification. A cell line clone with predetermined VCN was used as an inter-plate calibrator.
In vitro immortalization assay
Three independent in vitro immortalization (IVIM) assays using the EFS-T vector were performed at Hannover Medical School according to previously published protocols. 18 –20 For determining the incidence of positive and negative assays, potentially immortalized clones were discriminated from rare cases of background proliferation by the first quartile (Q1) expectation level of the positive control. A mutagenic vector such as RV-SF, at VCN levels above three copies, is expected to show positive assays with re-plating frequencies (RF) >3.17 × 104 in 75% of the cases (experience from metadata available at Hannover Medical School). All plates with a RF between the limit of detection (LOD; 1.05 × 104) and the Q1 level cannot be distinguished from spontaneous cell proliferation.
Flow cytometric analysis of cells from transplanted NSG mice
Peripheral blood and bone marrow of transplanted NSG mice was analyzed for human reconstitution by determining the percentage of cells positive for huCD45-APC (BD Biosciences, San Jose, CA) and for transduction efficiency by determining the percentage of GFP+ cells in the control group. For lineage analysis, the cells were stained with antibodies directed against CD33-PeCy7, CD15-PeCy7, CD19-BV605, and CD3-PE (all from BD Biosciences).
Statistics
The resorptive function of osteoclasts generated from NSG-engrafting vector-corrected IMO hematopoietic cells was analyzed statistically by comparing the EFS-T condition with the EFS-G condition using a two-sided Mann–Whitney test, where * indicates p < 0.05, ** indicates p < 0.01, *** indicates p < 0.001, and **** indicates p < 0.0001. Results are shown as the means ± standard error of the mean (SEM). For the IVIM assay, the significance of the differences between RF was calculated with Fisher's exact test, where * indicates p < 0.05.
Results
Restored resorptive function of osteoclasts from IMO patients after lentiviral-mediated TCIRG1 gene transfer driven by mammalian promoters
It was previously shown that CD34+ IMO cells transduced with SFFV-TG can be differentiated into functional osteoclasts in vitro. 4 In this study, the efficacy of two different mammalian promoters—EFS and ChimP—were evaluated by comparing them to SFFV-TG (Supplementary Fig. S1). All three TCIRG1-expressing vectors transduced approximately 35% of IMO CD34+ cells after two hits with a MOI of 30 (Fig. 1A). Transduced IMO and CB CD34+ cells were differentiated into osteoclasts on plastic, and TCIRG1 protein expression was analyzed by Western blot. TCIRG1 protein was expressed in the mature rescued osteoclasts at day 13 of osteoclast culture, but TCIRG1 protein was not detected in untransduced IMO cells or cells transduced with SFFV-G (Fig. 1B). The TCIRG1 levels were highest in cells exposed to the SFFV-TG vector followed by the EFS-TG vector and lowest in cells exposed to the ChimP-TG vector. Osteoclast differentiation on bone slices was verified by assessing TRAP activity in the media, and the ability to resorb bone was evaluated by measuring the release of Ca2+ and CTX-I. The Ca2+ and CTX-I levels increased in CD34+-derived IMO osteoclasts transduced with the rescue vectors compared to those transduced with the SFFV-G vector and the untransduced IMO osteoclasts, indicating an increase in resorptive activity and at least partial restoration of function (Fig. 1C and D). Once again, the levels were highest for cells transduced with SFFV-TG (Ca2+: 80 ± 11%; CTX-I: 81 ± 6%, relative to CB-derived osteoclasts), followed by EFS-TG (Ca2+: 72 ± 7%; CTX-I: 54 ± 16%), and finally ChimP-TG (Ca2+: 62 ± 6%; CTX-I: 35 ± 9%). Ca2+ and CTX-I levels remained unchanged in IMO SFFV-G osteoclasts compared to untransduced IMO osteoclasts. Media from the TCIRG1-transduced IMO osteoclasts had a slightly lowered TRAP activity compared to both SFFV-G and IMO controls (Fig. 1E). To evaluate further the effect of lentiviral gene transfer of TCIRG1 cDNA into IMO osteoclasts, bone slices were stained with hematoxylin to visualize resorption pits. After 13 days of differentiation, the mature IMO osteoclasts generated from cells transduced with the rescue vectors had formed a high number of clearly visible pits, whereas resorption pits were almost absent on bones with untransduced IMO cells and IMO cells transduced with SFFV-G (Fig. 1F). Although there was no statistically significant difference between the biomarker values for EFS-TG and ChimP-TG, there was a consistent trend present in four parameters (Ca2+, CTX-I, TRAP, and WB expression levels) in all experiments, indicating that EFS-TG was the vector that generated the highest expression levels and the best functional outcome in vitro of the tested mammalian vectors.
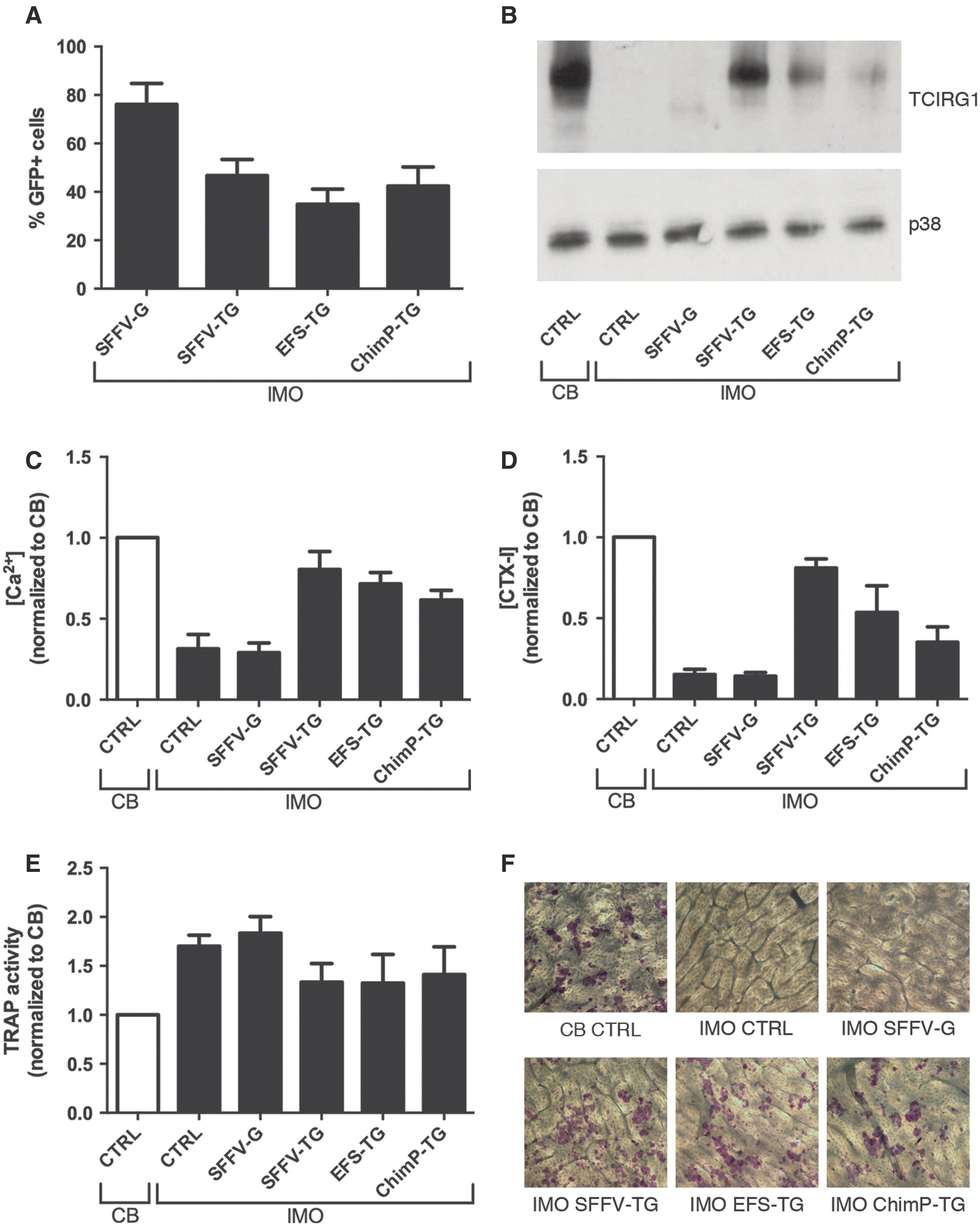
Lentiviral transduction of TCIRG1 driven by mammalian promoters can restore resorption in osteoclasts differentiated from infantile malignant osteopetrosis (IMO) CD34+ cells in vitro. CD34+ cells were transduced and expanded for 2 weeks, seeded on bone slices, and differentiated into osteoclasts for 13 days in the presence of macrophage colony-stimulating factor (M-CSF) and receptive activator of nuclear factor kappa-B ligand (RANKL).
Clinically relevant EFS-T vector can restore resorption in osteoclasts differentiated from IMO CD34+ cells in vitro
Having chosen the EFS promoter for continued experiments, the IRES and GFP were removed from the EFS-TG vector to obtain the clinically relevant EFS-T vector, which was then tested in vitro for efficacy and safety (Supplementary Fig. S1). Transduced IMO and CB CD34+ cells were differentiated into osteoclasts on plastic for analysis of TCIRG1 expression and on bone slices for resorption analysis. The expression of TCIRG1, evaluated by Western blot, was higher in the cells transduced with EFS-T than in those transduced with EFS-TG (Fig. 2A). Both the Ca2+ (Fig. 2B) and the CTX-I (Fig. 2C) levels in the media were higher when using EFS-T (Ca2+: 92 ± 5%; CTX-I: 95 ± 6%) than when using EFS-TG (Ca2+: 76 ± 14%; CTX-I: 67 ± 28%), and they were nearly comparable to those from CB-derived osteoclasts. TRAP activity was lower in the media from cells transduced with EFS-T than from those transduced with EFS-G, indicating a trend to normalization to the levels observed in the media of CB-derived osteoclasts (Fig. 2D). The osteoclasts derived from IMO cells transduced with EFS-T were capable of forming high numbers of resorption pits on bone slices (Fig. 2E), thus confirming the functional rescue of IMO cells in vitro with the EFS-T vector.
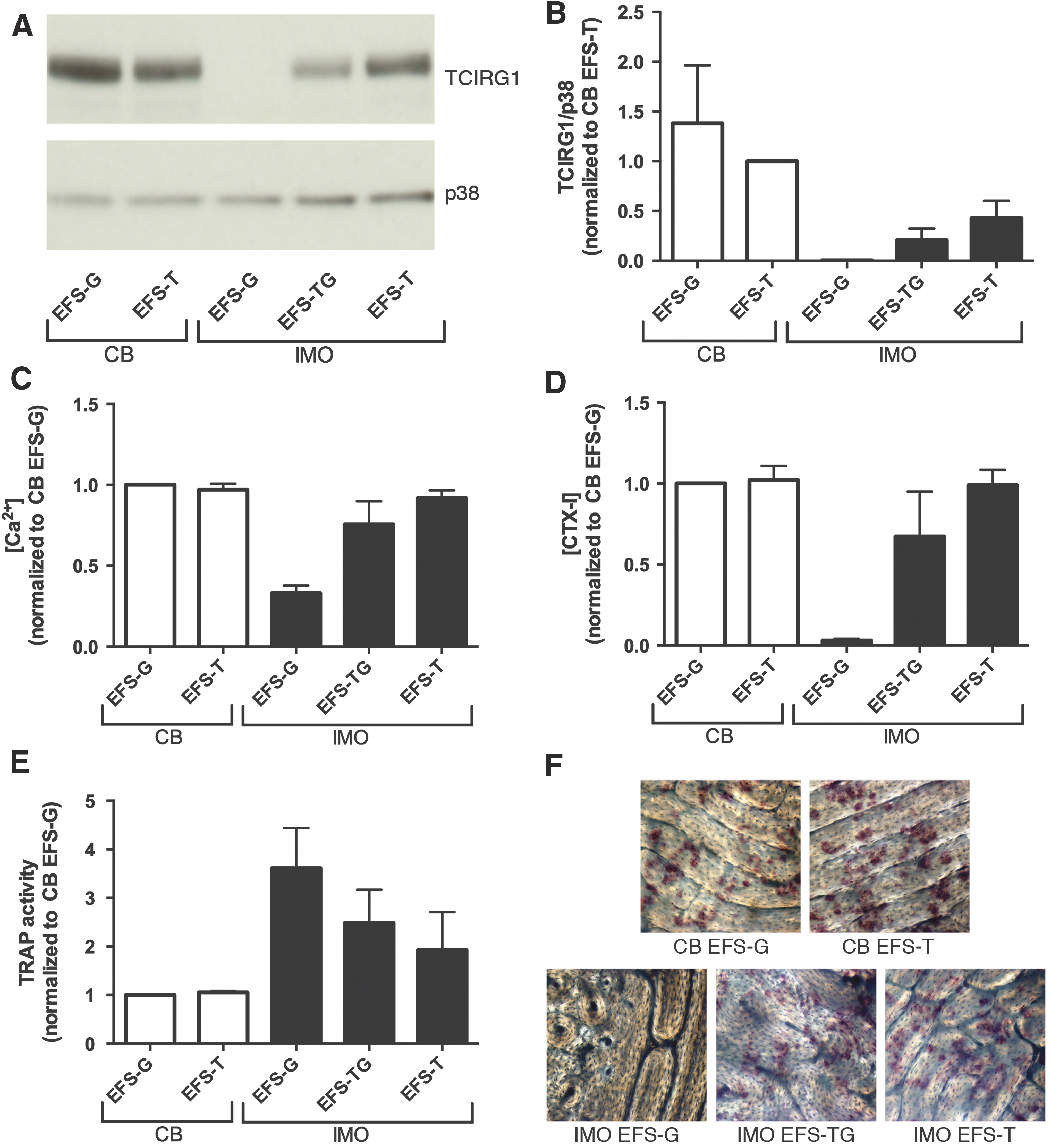
The clinically relevant EFS-T vector can restore resorption in osteoclasts differentiated from IMO CD34+ cells in vitro. CD34+ cells were transduced and expanded for 2 weeks, seeded on bone slices, and differentiated into osteoclasts for 13 days in the presence of M-CSF and RANKL.
EFS-T exhibits a low mutagenic potential compared to RV-SF and LV-SF
A safety concern regarding the clinical use of integrating viral vectors is the risk of insertional mutagenesis. The IVIM assay has demonstrated the capability to detect transformation of virally transduced cells under myeloid differentiation conditions. 18 In three independent IVIM assays, the EFS-T vector was compared to a gammaretroviral vector (RV-SF) and to a lentiviral vector, with the strong viral promoter SFFV (LV-SF) as positive controls. Cells were also subject to mock transduction in similar culture conditions, without viral vector, to monitor background activity. No re-plating clones were seen for the mock control or the EFS-T vector, whereas RV-SF-transduced samples induced clones in 5/8 cases, and the LV-SF-transduced samples induced clones in 2/5 cases (Fig. 3). The RV-SF vector had a mean RF of 8.34 × 10−3, the LV-SF vector had a mean RF of 1.24 × 10−3, and the EFS-T vector only had one positive well in 2/10 cases, corresponding to a mean RF of 2.63 × 10−5.

EFS-T exhibits a low mutagenic potential compared to RV-SF and LV-SF. Re-plating frequency (RF) of mock, RV-SF, LV-SF, and EFS-T transduced samples. Black bar indicates the mean RF. The filled circles for negative assays below the limit of detection (LOD) were manually inserted into the graph. The difference in incidence of positive (above the Q1 level) to negative assays for RV-FS to EFS-T was significant (p = 0.0256).
Long-term engraftment of transduced CB and IMO CD34+ cells in NSG mice
NSG mice (8–15 weeks old) were transplanted with untransduced CB CD34+ cells or IMO CD34+ cells transduced with either EFS-T or EFS-G (Table 1). The mice were sacrificed 9–19 weeks post transplantation, and bone marrow was analyzed for human reconstitution, the control animals transplanted with IMO cells transduced with EFS-G also for the level of GFP marking. In all groups of transplanted mice, human CD45+ cells were on average around 35% (Fig. 4A), showing that peripheral blood IMO CD34+ cells have the capacity to engraft in NSG mice similarly to CB CD34+ cells. In addition, GFP-marked cells were found in all mice transplanted with IMO cells transduced with EFS-G (1.1–12.6% of huCD45+ cells). No differences in lineage distribution of bone-marrow cells harvested from mice transplanted with human CB CD34+ cells or with IMO CD34+ cells transduced with EFS-G or EFS-T were observed, indicating that EFS-T does not skew the differentiation potential of transduced CD34+ cells in the NSG model (Fig. 4B).
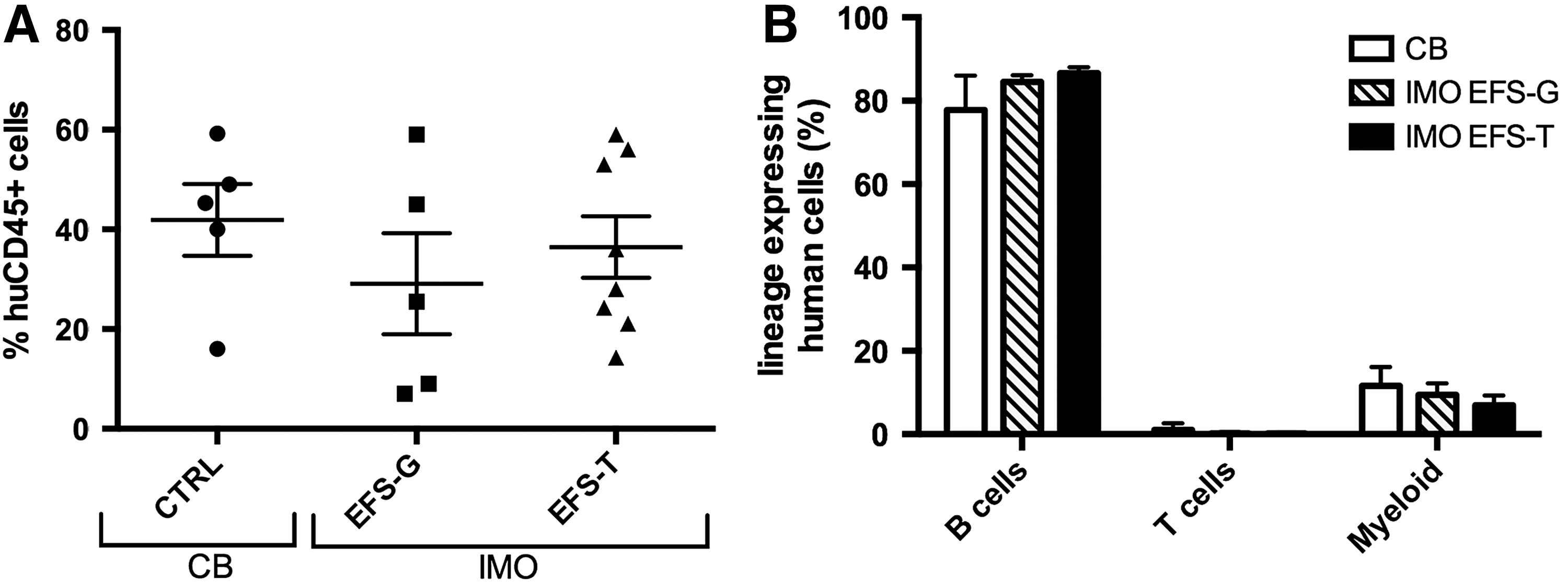
Long-term engraftment and lineage analysis of cord blood (CB) cells and transduced IMO CD34+ cells transplanted into NSG mice. Bone-marrow cells were harvested from NSG mice 9–19 weeks after transplantation with CB cells (n = 5) or IMO cells transduced with the EFS-G (n = 5) or EFS-T vector (n = 8) and were assessed for human engraftment in total bone marrow
Data from individual NSG mice transplanted with IMO or CB CD34+ cells
Rescue was evaluated as restoration of osteoclast resorption compared to positive and negative controls.
IMO, infantile malignant osteopetrosis; MOI, multiplicity of infection; VCN, vector copy number; P1–P7, IMO patients; CB, cord blood; UT, untransduced; n.d., not determined.
Restored resorptive function of osteoclasts generated from NSG-engrafting vector-corrected IMO hematopoietic cells
Bone-marrow cells were harvested from NSG mice 9–19 weeks after transplantation with CB cells (n = 5) or IMO cells transduced with EFS-G (n = 7) or EFS-T (n = 11). Human CD34+ cells were isolated (range of CD34+ cells per mouse 1.5–9.6%), expanded for 2 weeks, seeded on plastic or bone slices, and differentiated into osteoclasts for 13 days in the presence of M-CSF and RANKL. Western blot analysis was performed on lysates from mature osteoclasts differentiation and showed the presence of the a3 subunit after rescue in 4/8 of the mice transplanted with IMO EFS-T cells. Lysates from two different mice in this group are shown (Fig. 5A), one in which TCIRG1 could be detected and one in which it was not detected, suggesting that there was variability of rescue of NSG-engrafting cells between mice. Vector-corrected IMO osteoclasts exhibited completely restored Ca2+-release (97 ± 21% compared to CB-derived osteoclasts), with a 3.4-fold increase (Fig. 5B; p = 0.019) relative to the non-corrected IMO osteoclasts. CTX-I levels in the media were partially restored (33 ± 6% compared to CB-derived osteoclasts) and 14-fold (p < 0.0001) higher than those of the non-corrected IMO osteoclasts, which failed to resorb bone (Fig. 5C). There was no difference in TRAP activity of the osteoclasts derived from mice transplanted with IMO cells transduced with EFS-G or EFS-T (Fig. 5D). VCN was assessed in eight mice transplanted with EFS-T-transduced CD34+ cells and ranged from 0.10 to 0.46 VCN per human CD45+ cell for all mice, except one where VCN was 1.96 (Table 1). The VCN did not correlate directly with the MOI of the transduction. In transplantation experiment 4, the use of a MOI of 100 instead of a MOI of 30 did not result in a higher VCN or higher rescue (Table 1). Overall, these data show that the EFS-T vector can at least partially restore the resorptive function of osteoclasts differentiated ex vivo from NSG-engrafting vector-corrected CD34+ cells from IMO patients.

Vector-corrected IMO osteoclasts generated from NSG-engrafting hematopoietic cells show rescued resorption capacity. Bone-marrow cells were harvested from NSG mice 9–19 weeks after transplantation with CB cells (n = 5) or IMO cells transduced with the EFS-G (n = 7) or EFS-T vector (n = 11). Human CD34+ cells were expanded for 2 weeks, seeded on bone slices, and differentiated into osteoclasts for 13 days in the presence of M-CSF and RANKL.
Discussion
The aim of the present study was to use lentiviral-mediated gene transfer of TCIRG1 with a clinically applicable vector to rescue the phenotype of human IMO osteoclasts in vitro and after generation of osteoclasts from NSG-engrafting hematopoietic cells.
In a previous proof-of-principle work, sufficiently high levels of transgene expression were obtained in IMO osteoclasts in vitro by using the viral SFFV promoter. 4,21 In the current work, the EFS promoter that is being used in the X-SCID 22 –24 and ADA SCID 25 clinical trials and the chimeric myeloid promoter (ChimP), 15 planned to be used in the Chronic Granulomatous Disease (CGD) trial and with the advantage of being lineage specific, were tested. 26 Due to the limited patient material available, the aim was to choose the non-viral promoter that could induce the highest resorption rescue in IMO osteoclasts. Therefore, the EFS promoter and the ChimP were compared to the SFFV promoter. The EFS-TG vector mediated a higher level of protein expression and higher rescue of resorptive function of the IMO osteoclasts in vitro than the ChimP-TG vector, as evaluated by measuring release of calcium and the resorption marker CTX-I into the media. From a safety point of view, it has previously been shown that lentiviral-mediated expression of TCIRG1 is regulated in the same manner as the endogenous gene product, despite being expressed by a lentiviral vector with a generic promoter. 21 Thus, the myeloid specificity of the ChimP is not strictly necessary.
Based on the results from the comparison of the three promoters in vitro, the clinically applicable EFS-T vector was developed, with a mammalian promoter and without GFP, for continued experiments. EFS-T-corrected IMO osteoclasts in vitro restored release of calcium and CTX-I into the media corresponding to 92 ± 5% and 95 ± 6% of those obtained with CB-derived osteoclasts. The overall higher rescue level observed with the EFS-T vector compared to the EFS-TG vector is most likely due to the removal of the IRES-GFP sequence allowing for a higher expression of TCIRG1.
For the in vivo experiments, CD34+ cells from peripheral blood of five patients with different TCIRG1 mutations were obtained without the need for mobilization due to the high percentage of circulating CD34+ cells characteristic of IMO patients. 27 The cells were transduced with EFS-T or with EFS-G and transplanted into sub-lethally irradiated NSG mice; CB CD34+ cells were transplanted as positive controls. It was not possible to analyze the correction of osteoclasts in vivo, as these cells do not develop in this xenotransplant model, probably due to the species specificity of M-CSF. 28 Therefore, the bone marrow of the mice was harvested 9–19 weeks after transplantation for analysis and osteoclast differentiation ex vivo. IMO CD34+ cells from peripheral blood engrafted in NSG mice to the same degree as CB CD34+ cells, in line with the observation that they can be used as a backup in clinical transplantations should a graft failure occur. 27 Isolated human CD34+ cells were differentiated to mature osteoclasts, and resorption was assessed. The positive and negative control osteoclasts performed as expected: osteoclasts derived from mice transplanted with CB cells were capable of resorbing bone effectively ex vivo, while osteoclasts derived from mice transplanted with IMO EFS-G cells exhibited strongly impaired resorption. 3,29 The main objective was to assess the functional restoration of the osteoclasts derived from the mice transplanted with IMO EFS-T cells. On average, based on calcium release, resorption was completely restored compared to that of osteoclasts derived from animals transplanted with CB cells, and based on CTX-I levels, it was restored to 33 ± 6%, as well as being significantly higher compared to IMO EFS-G osteoclasts. The rescue levels in individual mice were variable and can be divided into three categories: in osteoclasts derived from CD34+ cells isolated from two mice, no rescue was observed; in cells derived from eight mice, partial rescue was detected; and in cells derived from one mouse, a complete rescue was observed, as assessed by an increase in calcium and CTX-I levels, a decrease in TRAP activity, and comparison to the results of osteoclasts derived from mice transplanted with CB cells. This variability is also seen in the VCN analysis that ranged from 0.10 to 0.46 per cell positive for human CD45+ cells in the mice with no rescue or partial rescue, whereas it was 1.96 in the mouse whose cells conferred complete rescue. This indicates that the variability in resorption is probably due to differences in transduction efficiency of the more primitive cells. This does not seem to be directly correlated to the MOI, as an increase in VCN was not seen when using a MOI of 100 instead of a MOI of 30 for the in vivo experiments, but could instead be strongly influenced by the use of different patient samples and vector batches. A future aim is to optimize and standardize the vector production and transduction protocol in order to obtain a more consistent VCN ranging between 1 and 2, in the NSG-engrafting putative stem cells, which should allow a higher level of rescue to be observed in cells harvested from mice. It is possible to aim safely for a higher VCN, as the results from the IVIM assay that was performed conclude that the EFS-T vector has a strongly reduced mutagenic potential when compared to the gammaretroviral positive control vector or a SIN-lentiviral vector with a strong viral promoter/enhancer element, even at mean VCN levels above three copies per cell.
For future development and application of a clinical gene therapy protocol for treatment of IMO, a crucial question is what level of correction of osteoclast function is needed in vivo to reverse the disease phenotype. It was previously shown that transplantation of gene therapy–corrected cells in oc/oc mice completely reversed the disease, even though the in vitro bone resorption capacity of these cells was only 10% of wild-type cells. 30 Furthermore, it was possible to show that transplantation of wild-type cells in oc/oc mice in a non-myeloablative setting, resulting in an engraftment level of only 4–5%, was sufficient to correct the disease. 31 In terms of the human disease form, the addition of 30% umbilical CB CD34+ cells to IMO CD34+ cells in vitro, followed by osteoclast differentiation, was sufficient to restore resorptive function of these cells completely. 21 Furthermore, in the same study, levels as low as 5% CB CD34+ cells or gene-corrected IMO CD34+ cells mixed into non-manipulated IMO CD34+ cells resulted in significant resorption, possibly due to fusion of preosteoclasts harboring normal/gene corrected TCIRG1 with TCIRG1 deficient counterparts to form osteoclasts. 4,21,31 Thus, gene correction of only a fraction of cells and partial rescue of osteoclast function may be sufficient for clinical benefit when treating patients with IMO.
In summary, this study provides evidence for almost complete rescue of IMO osteoclasts in vitro by a clinically applicable lentiviral vector expressing TCIRG1 under the mammalian promoter EFS and lacking a marker gene. Furthermore, it shows partial rescue of IMO osteoclasts generated from vector-corrected, NSG mice-engrafting hematopoietic cells. These findings support further development of hematopoietic stem cell targeted gene therapy, not only of IMO, but also of other diseases affecting osteoclasts.
Footnotes
Acknowledgments
We thank Adrian Thrasher and Manuel Grez for the ChimP construct, and Christopher Baum and Axel Schambach (Hannover Medical School, Germany) for providing us with the lentiviral vectors. K.H. is supported by the Danish Research Foundation (Den Danske Forskningsfond). J.R. is supported by grants from The Swedish Childhood Cancer Foundation and a Clinical Research Award from Lund University Hospital, The Foundations of Lund University Hospital. A.S. is partially supported by grants from the EU (ERARE initiative, project OSTEOPETR). A.S. and M.R. were supported by the EU (FP7-Health-2010-CELL-PID: #261387). H.L. is supported by Marie Curie Initial Training Networks (Euroclast, FP7-People-2013-ITN: # 607446). The Lund Stem Cell Center is supported by a Center of Excellence grant in life sciences from the Swedish Foundation for Strategic Research. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Author Disclosure
J.R. has subsequent to acquisition of data presented here entered into a consultancy agreement with Rocket Pharmaceutical (New York, NY) for the development of clinical gene therapy of IMO. No competing financial interests exist for the remaining authors.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.