
Abstract
Cystic fibrosis (CF) lung disease is an ideal candidate for a genetic therapy. It has been shown previously that preconditioning with lysophosphatidylcholine (LPC) prior to lentiviral (LV) vector delivery results in long-term in vivo gene expression in the airway epithelium of CF mice. It was hypothesized that this outcome is largely due to transduction of airway basal cells that in turn pass the transgene onto their progeny. The aim of these studies was to confirm if the in vivo delivery of a human immunodeficiency virus type 1 (HIV-1) vesicular stomatitis virus envelope glycoprotein (VSV-G) pseudotyped LV vector following LPC airway conditioning results in transduction of mouse airway basal cells in situ and if the transgene is passed onto their progeny. Additionally, the study sought to determine the efficiency of in vitro transduction of human airway basal cells. First, normal mouse nasal airways were pretreated with LPC prior to delivery of a HIV-1 VSV-G pseudotyped LV vector carrying a LacZ marker gene (LV-LacZ). An epithelial ablation model utilizing polidocanol was then used to demonstrate that clonal outgrowth of linear and spotted clusters of transgene expressing ciliated, basal, and goblet cells occurs following transduction of basal cells. Second, human basal cells were cultured from primary bronchial epithelial cells, with identity confirmed by keratin 5 staining. High levels of transgene expression were found following LV-LacZ transduction. This study demonstrates the ability of the vector delivery protocol to transduce mouse airway basal cells, the LV vector to transduce human basal cells, and the likely role of these cells in maintaining long-term gene expression. These findings support and further develop the potential of LV gene transfer for persistent correction of CF airway disease.
Introduction
A gene-addition therapy, where a fully functioning copy of the CFTR gene is added into defective cells, has the potential to correct the underlying gene and subsequent protein defects associated with all CF mutation classes. Both non-viral and viral vectors have been explored for airway gene therapy, with encouraging results from studies using integrating viral vectors such as lentiviruses. 4,5 Lentiviral (LV) vectors have the unique advantage of transducing both dividing and nondividing cells, raising the possibility of producing lifelong benefit following a single treatment, via the integration of a CFTR transgene into resident airway progenitor cells. 6 –8
While there is still much to be learnt about the organization and regulation of airway epithelial stem and progenitor cell compartments, precise cell lineage tracing studies in mice have confirmed that basal cells are the progenitor cell precursor of the secretory and ciliated epithelial cell lineages of the proximal airways. 9,10 Basal cells are also considered the dominant progenitor cell type responsible for maintenance and repair of human airways. 11 Although basal cells extend to the terminal bronchioles in humans, they are confined to the upper airways in mice, so the mouse proximal airways are a highly relevant model when exploring airway basal cells as a potential target for in situ long-term gene therapy correction. 12
It has been shown that LV vectors provide effective gene transduction in live mouse airways. In some cases, a single dose of a LV-LacZ vector, delivered 1 h after conditioning the airway with lysophosphatidylcholine (LPC), was sufficient to induce LacZ reporter gene expression for the lifetime of the mouse. 5 In separate studies, significant therapeutic CFTR gene expression lasting for at least 12 months has been shown in a CF mouse model. 13 These long expression periods suggest that the transgenes were inserted into basal cells of the respiratory epithelium. Interestingly, the observation that some LacZ-expressing cell types present at day 92 were not detected shortly after transduction also suggests that transgenes inserted into airway basal cells were passed on to their progeny through normal ongoing mitotic division and differentiation. 5
The advantages of transducing airway basal cells in situ are threefold. First, a single airway basal cell has the potential to generate ∼8 × 103 daughter cells, thus allowing extensive multiplication of benefit. 14 Second, although LV vectors are less immunogenic upon re-administration than other viral vectors, the use of a single-dose protocol that produces transduction of airway basal cells may avoid the need for re-dosing. 15 Third, the delivery of a gene therapy that corrects airway basal cells in situ is a more practical and less invasive approach than methods that reseed the airways with expanded cells corrected in vitro.
The first aim of this study was to use a well-characterized polidocanol (PDOC) model of epithelial injury and repair 4,16 to verify the potential of the LPC pre-treatment and LV vector to transduce airway basal cells in situ, and to demonstrate that they pass the transgene onto their progeny. Although these two compounds are similar in structure and function, in this study, they are used at different concentrations and for different functions. In this setting, LPC is used at a low concentration for airway conditioning prior to LV vector delivery to open the tight junctions between the epithelial cells transiently, allowing access to the vesicular stomatitis virus envelope glycoprotein (VSV-G) receptors on the basolateral cell surface, as well as the basal cell niche on the basement membrane. PDOC is used at a high concentration as an experimental compound (and is not designed to be used as part of a clinical protocol) to injure the airway epithelium and remove the terminally differentiated cells, thereby forcing regeneration of the epithelium from the basal progenitor cells that are left intact through the forced injury process.
The second aim of the study was to assess the efficiency of the LV vector to transduce human airway basal cells. This was investigated by treating isolated and cultured human airway basal progenitor cells with the same LV-LacZ vector at different multiplicities of infection (MOIs) in two-dimensional submerged cultures.
Methods
All animal studies were approved by both the University of Adelaide and the Women's and Children's Hospital animal ethics committees.
Mouse models
C57BL/6 mice, 6–8 weeks of age, were used to assess LV transduction of airway basal cells. To assess the proliferation index of the CF airway epithelium, CF mice (cftrtm1unc )-Tg (FABp CFTR) were compared to CF heterozygous control mice (cftrtm1unc ). Female mice were used in all studies.
For all recovery studies, mice were anesthetized with a mixture of 10 μL/g body weight of medetomidine (Domitor; 0.1 mg/mL; Orion Corporation, Espoo, Finland) and ketamine (7.6 mg/mL; Parnell Laboratories, Alexandria, Australia), delivered by an intraperitoneal (i.p.) injection. After procedures, anesthesia was reversed with an i.p. injection of atipamezole hydrochloride (Antisedan; 1 μg/g; Orion Corporation). Mice were kept in a humidified incubator (32°C) and monitored until recovery from the anesthetic was complete. At experiment completion, the animals were humanely killed via 100% CO2 inhalation.
LV vector production
A human immunodeficiency virus type 1 (HIV-1) VSV-G pseudotyped LV vector expressing nuclear localized LacZ (LacZnlsco) under transcriptional control of the myeloproliferative sarcoma virus (MPSV) promoter, designated here as LV-LacZ, was produced according to previously published methods. 17 The virus titer was 2 × 109 TU/mL for the in vivo studies and 1.4 × 109 TU/mL for the in vitro studies, as assayed by quantitative polymerase chain reaction. 18
Induced regeneration of the airway epithelium via PDOC ablation
To determine if transduced basal cells in the nasal respiratory epithelium pass the transgene onto their progeny upon differentiation, regeneration of the respiratory epithelium was induced using PDOC (Polyoxyethylene 9 Lauryl Ether; Sigma–Aldrich, St Louis, MO), following LPC and LV-LacZ delivery.
In a pilot study prior to the gene delivery studies, the nasal epithelial layer of C57BL/6 mice (n = 9) was treated with a 10 μL volume of 2% (w/v) PDOC (delivered via inhalation-driven instillation, as previously described) into one nostril to verify the effect and duration of the transient epithelial ablation and regeneration. 4,16 The untreated nostril was used as a control. Following PDOC treatment, mice were treated with buprenorphine (2 μg/g; Reckitt Benckiser, North Chesterfield, VA) for pain relief, a prophylactic measure for potential pain as advised by the animal ethics committee. Relevant nasal tissue samples were collected 24 h (n = 3), 4 days (n = 3), or 7 days (n = 3) post PDOC treatment and examined histologically.
In the primary study, one nostril of C57BL/6 mice (n = 44) was treated with LPC and LV-LacZ vector, and the second nostril was used as an untreated control, as previously described.
18
Prior to vector instillation the airways were conditioned with 4 μL of 0.3% L-α-lysophosphatidylcholine (cat. #L4129; Sigma–Aldrich) in phosphate-buffered saline (PBS). The LV vector (20 μL, as used in previous gene transfer studies) was administered 1 h later into the same nostril (2 × 10 μL aliquots over 2–3 min).
19
Following vector delivery, the mice were randomly separated into four groups (n = 11 per group): (1) Short-term control: the mice were humanely killed at 7 days following vector instillation to determine the short-term gene expression profile. (2) 1 × PDOC ablation: 4 weeks following vector instillation, the mice were treated with PDOC (same manner as described above for the pilot study); at 8 weeks they were treated with PBS, and then they were humanely killed at 14 weeks. (3) 2 × PDOC ablation: 4 and 8 weeks following vector instillation, the mice were treated with PDOC, and then they were humanely killed at 14 weeks. (4) Long-term control: 4 and 8 weeks following vector instillation, the mice were sham treated with PBS and humanely killed at 14 weeks.
Identification of marker gene expressing cell types
After the animals were humanely killed, LacZ gene expression in the nasal airways was revealed using the standard X-gal processing method. 5 The distribution and pattern of transduced cells was first confirmed by en face photography. To identify the cell types expressing the LacZ marker gene, 5 μm wax sections were mounted on glass slides, processed, and histochemically stained using hematoxylin and eosin or Safranin-O. Alcian blue/periodic acid-Schiff staining was employed to determine if mucin secretory cells not directly transduced by the VSV-G pseudotype expressed the LacZ marker following forced regeneration. 20
Assessment of airway basal cell proliferation
Tracheal tissue (n = 6 mice) was dissected from mice immediately following humane killing and placed into 10% neutral buffered formalin overnight. Ki-67 staining was performed, as previously described. 21 The proportion of proliferating airway basal cells in the CF mice and heterozygous control mice was determined by analysis of Ki-67 staining. 21
Assessment of in vitro human airway basal cell transduction
To assess the potential of the LV-LacZ vector to transduce human airway basal cells, it was tested in a human airway basal cell culture system. Human primary airway cells (cc-2540s; Lonza, Mount Waverley, Australia) were seeded and basal cells expanded by passaging twice in 25 cm2 collagen-coated flasks containing Bronchial Epithelial Cell Growth Medium (BEGM; cc-3170; Lonza) and were incubated at 37°C and 5% CO2.
Cells were harvested at 90% confluency for reseeding, and samples were taken to confirm cell type. Cells were transferred to chamber slides (n = 3) coated with type 1 rat tail collagen and incubated overnight at 37°C and 5% CO2 in BEGM media. The slides were then processed and stained using anti-Cytokeratin 5 antibody (1:350; Abcam 52635; Abcam, Cambridge, United Kingdom) and goat anti-rabbit IgG H&L (1:500; Abcam 97051; Abcam) to verify that the expanded cells were airway basal cells.
Harvested cells were also reseeded into Nunc™ six-well plates coated with type 1 rat-tail collagen (152034; Thermo Fisher Scientific, Scoresby, Australia) to assess transduction efficiency. Cells were seeded at a density of 2.5 × 105 cells per well to be 75% confluent once adhered (approximately 2 h later). The seeded wells were divided into four groups (n = 3 per group) and treated with LV vector (no LPC pretreatment) at MOIs of 0, 1, 10, and 100. The cells were then incubated at 37°C and 5% CO2 overnight in a humidified incubator. The following day, the media was removed by aspiration, cells were washed with PBS, fresh media was added, and cells were returned to the incubator for a further 3 days.
To analyze LacZ gene expression, cells were fixed with 0.1% glutaraldehyde for 15 min in a shaking incubator at 35 rpm, and then the fixative was removed by aspiration. Cells were washed three times (5–15 min) with 1 mL of 1 mM MgCl2 per well, with the second and third washes placed in a shaking incubator at 35 rpm. One milliliter of X-gal solution was added to each well and incubated at 37°C and 5% CO2 for 3 h. 5 Quantification was performed by capturing images of five random fields in each well at 100 × magnification and counting the proportion of LacZ-positive and LacZ-negative cells to allow a percentage of positive cells to total cells to be obtained.
Statistics
Analysis was overseen by the institutional biostatistician. Results are represented as the mean and standard error. Data were tested for normality and analyzed using GraphPad Prism v6 (GraphPad Software, La Jolla, CA). Statistical significance was set at p = 0.05.
Results
Epithelial regeneration following PDOC-induced epithelial ablation
A pilot study was first performed to confirm that an appropriate level of nasal epithelial stripping was induced followed PDOC delivery. Histological analysis of mouse nasal airway tissue collected 24 h post PDOC instillation (n = 3) confirmed that ablation of nasal airway epithelium to the basement membrane had occurred (Fig. 1a). Four days following the PDOC instillation (n = 3), the epithelial layer had partially regenerated (Fig. 1b). By seven days post-PDOC instillation (n = 3), the airway epithelium had regenerated to the normal histological appearance of ciliated respiratory epithelium (Fig. 1c). The untreated nostrils of all mice (within-animal control) showed no signs of airway cell layer ablation (data not shown). These results confirm those previously reported for the PDOC ablation model. 4,16
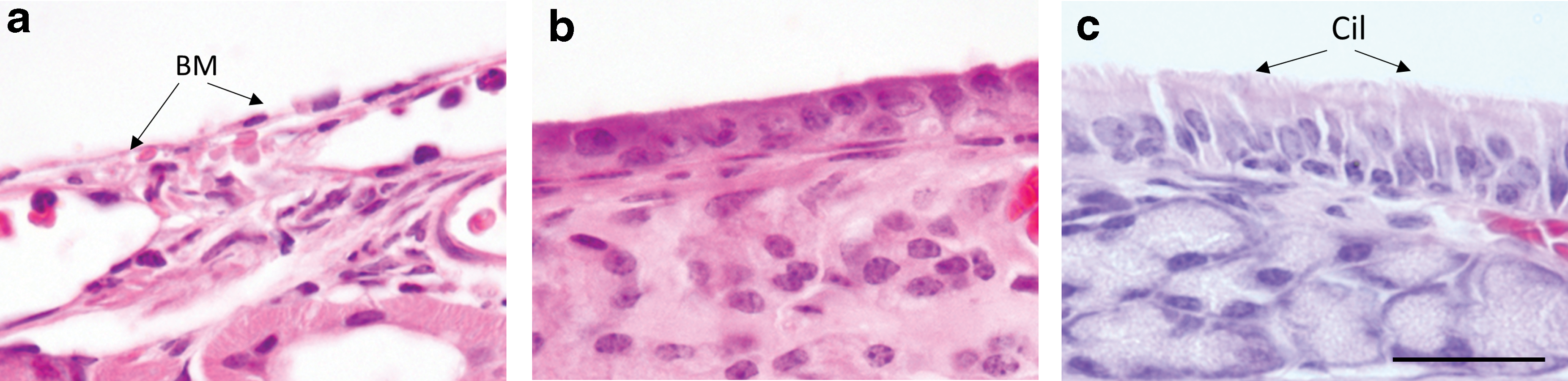
Assessment of the effects of polidocanol (PDOC) on the nasal septa of three animals showing:
Induced regeneration of the respiratory epithelium produces clonal clusters of LacZ positive cells
Three mice died during the study (one from the 1 × PDOC group, and one from the 2 × PDOC group). The short-term controls (7-day, no-ablation) displayed a typical finely speckled pattern of LacZ expression in the nasal airway epithelium (Fig. 2a and Supplementary Fig. S1). The long-term (14-week, no-ablation) controls displayed a similar speckled pattern, but with some larger clusters also present (Fig. 2b and Supplementary Fig. S2). In contrast, after PDOC treatment, only clusters of LacZ-expressing cells were observed, with no speckling present (Fig. 2c and d and Supplementary Figs. 3 and 4). Both PDOC ablation groups displayed numerous cluster shapes, ranging from round (Fig. 2c and d) to linear (Fig. 3a and b).
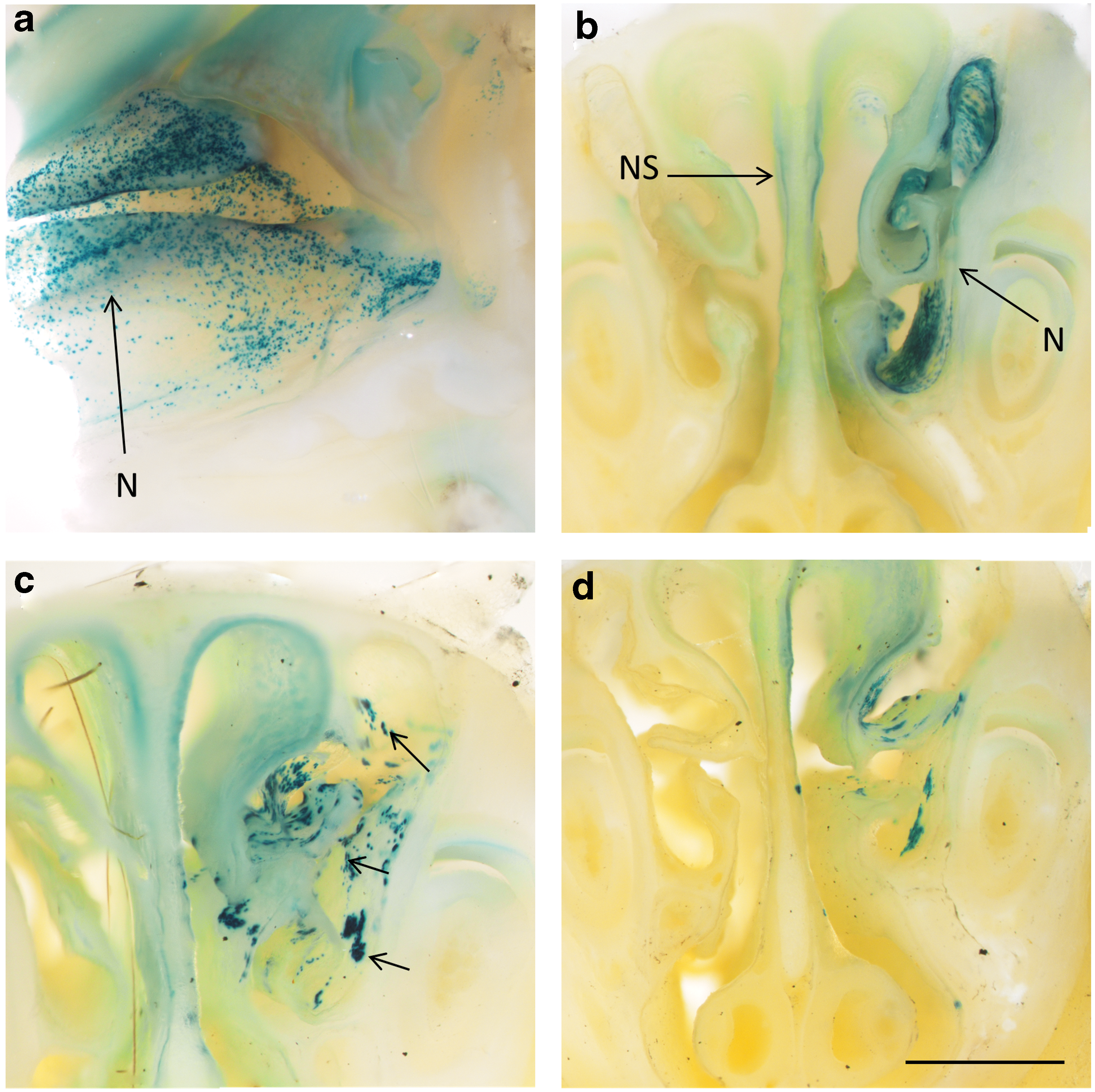
LacZ expression patterns in the treated nasal airway show evidence of clonal expansion patterns after PDOC treatment.
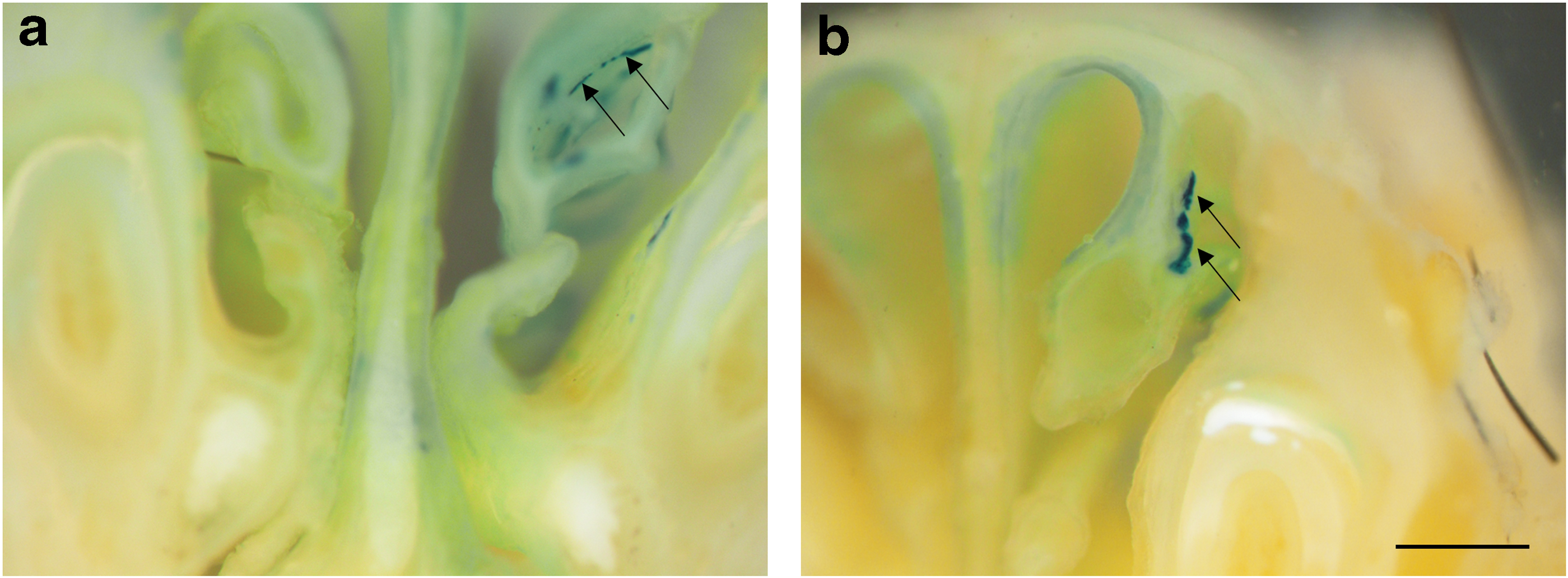
Examples of linear clusters
Identification of marker gene expressing cells types
Histological analysis of LacZ-expressing cell types revealed transduced ciliated cells and basal cells in all short-term (7-day) control animals. These same cell types were also present in all long-term control and the PDOC ablation groups. LacZ-expressing goblet cells were also present in these groups (Fig. 4).

Serial sections of nasal tissue were stained using
Proliferation of tracheal airway basal cells is greater in CF mice
Figure 5c shows a 2.1-fold increase in actively cycling cells in the tracheal epithelium of CF mice compared to control mice (p < 0.05, n = 5). Most of the actively cycling cells were found at or close to the inter-tracheal cartilage zone of the control mice (Fig. 5a). In CF mice, these actively cycling cells extended further into the tracheal cartilage zone (Fig. 5b).
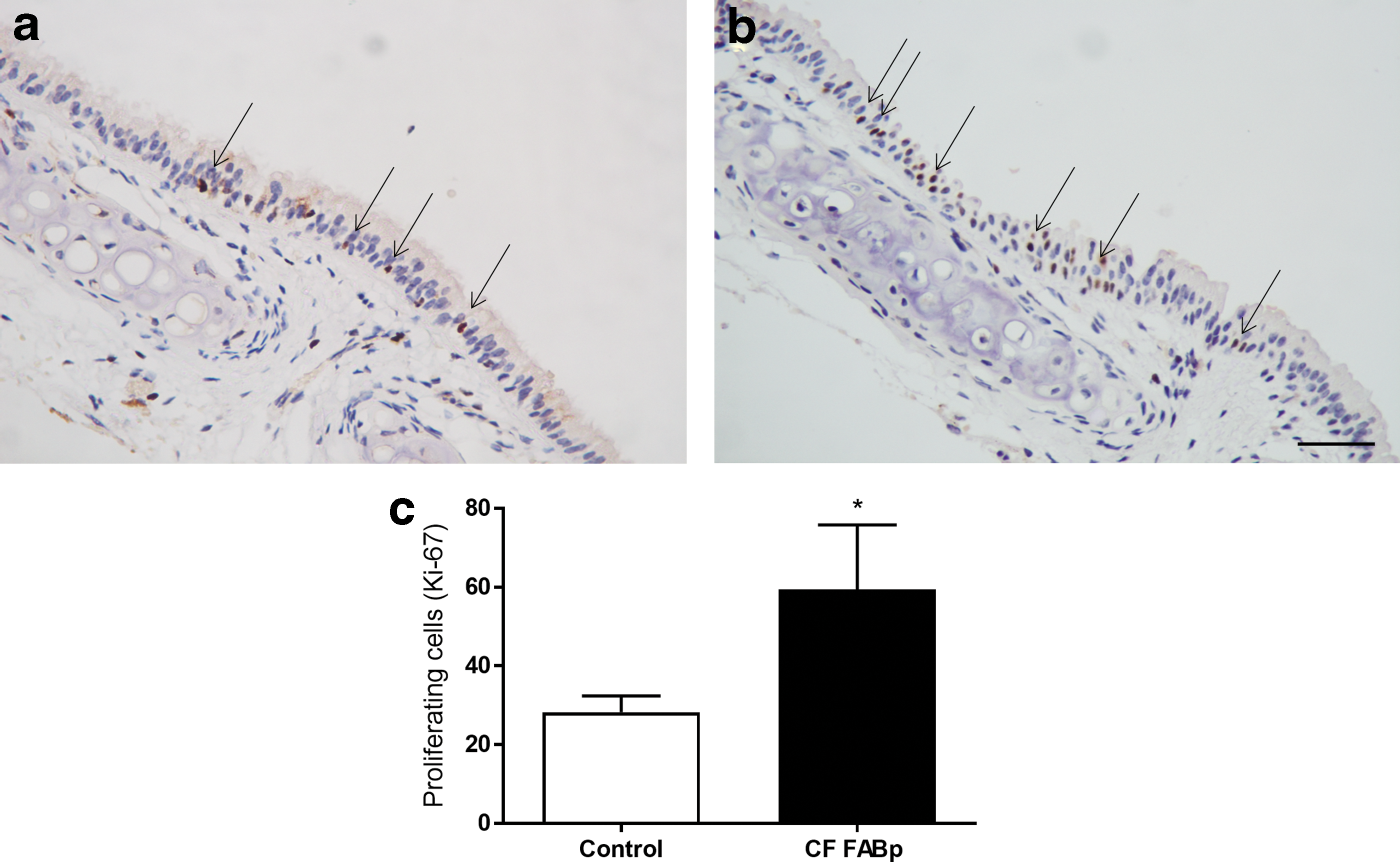
Ki-67 immunohistochemistry of the tracheal airway epithelium from
Assessing in vitro human airway basal cell transduction
To determine the efficiency of the LV vector to transduce human airway basal cells, wells were treated at MOIs of 0, 1 (Fig. 6a), 10 (Fig. 6b), and 100. As expected, LacZ expression was absent in the no-vector control group (not shown). The treated groups showed average transduction efficiencies of 38.3% for a MOI of 1, 94.5% for a MOI of 10, and 99.1% for a MOI of 100 (Fig. 6c). The cells expanded and passaged from human primary bronchial epithelium in basal cell media were validated as being cytokeratin 5 positive airway basal cells (Fig. 6d).

LacZ transduction of human airway basal cell cultures at
Discussion
Ideally, a gene-addition therapy for CF-related airway disease will produce lifelong expression of an introduced functioning copy of the CFTR transgene. It is proposed that a single dose of a chromosomally integrating CFTR viral vector, capable of transducing the airway basal cells that are responsible for lifelong repair and regeneration of the respiratory epithelium, may provide an effective treatment or cure for CF airway disease. Previous results show that the ability to enable vector access to the basolateral cell surface following LPC conditioning substantially boosts initial levels of gene expression, but also provides the opportunity to transduce the basal cells that reside on the epithelial basement membrane. This study sought to verify that the vector delivery protocol facilitates in situ transduction of airway basal cells, and that transduced basal cells subsequently transmit the transgene onto their progeny. Additionally, the study aimed to investigate the potential for translating this system into human airways by utilizing in vitro cultures to ascertain the transduction efficiency in human airway basal cells.
Denuding the airway epithelium using PDOC creates an environment where basal cells, which are not removed by the PDOC treatment, are stimulated to proliferate and differentiate to regenerate the injured epithelium. If a proportion of basal cells are transduced following the LPC conditioning and LV vector delivery, and the transgene is passed on to the progeny, then this will be revealed as marker gene positive clusters of terminally differentiated cells once the epithelium is fully regenerated. Analysis of the distribution and size of clusters of transduced cells provides valuable information about the incidence and replicative potential of transduced cells. Figure 1 demonstrates that PDOC treatment in non-gene-treated transduction control mice resulted in epithelial stripping and subsequent regeneration over 7 days, validating the previously described model. 4,16 Following pretreatment and gene vector delivery, cells expressing LacZ were present in the nasal airway after both single and double PDOC ablations. The presence of clusters of LacZ-expressing cells following two PDOC treatments suggests that the clusters observed from the single ablation were unlikely to be a consequence of incomplete stripping of transduced surface epithelial cells during the ablation treatment.
En face comparison of the patterns of expression provides clues to the underlying mechanisms of sustained gene expression. The speckled pattern of mostly individual isolated LacZ-expressing cells seen in the 7-day and 14-week control groups that did not receive PDOC was clearly different to the clustered pattern of LacZ-expressing cells in both PDOC groups (Fig. 2 and Supplementary Figs. S1–S4), likely due to the presence of initially transduced terminally differentiated cells. In contrast, the clustered appearance of LacZ-expressing cells in the long-term control and PDOC groups is consistent with a pattern of clonal expansion where transduced airway basal cells have passed the transgene onto their progeny during epithelial regeneration, whether as part of the normal cellular proliferation or in response to PDOC. In the PDOC-treated groups, there were few individually transduced cells present, confirming that transduced terminally differentiated cells located above the basement membrane had been removed by the ablation process. These observations validate the role of transduced basal cells in long-term reporter and therapeutic CFTR transgene expression demonstrated in previous studies by Limberis et al and Stocker et al. 5,13 While the number of clonal clusters of transgene positive cells shown in Fig. 2 and Supplementary Figs. S3 and S4 are modest, and may not provide a level of expression needed for the desired sustained therapeutic outcome, these results are an important step that can be built upon through strategies developed to maximize the targeting of airway basal cells in situ.
Clonal expansion from airway basal cells transduced with the LacZ marker gene has been predicted to produce round-shaped LacZ positive clusters of cells upon epithelial repair and regeneration, as the newly differentiated cells expand out from the transduced airway basal cell. 4 In the single and double ablation groups in the nasal airways, symmetrical clustering of LacZ positive cells was observed (Fig. 2c). Unexpectedly, in conjunction with the observation of rounded LacZ-positive cell clusters, linear cell clusters were also present (Fig. 3). The linear cell cluster pattern may have been a result of the fluid dynamics of the introduced doses during the instillation of the LPC and vector that caused the transduction of airway basal cells along a linear track. Alternatively, there may be some mechanism that causes basal cells to proliferate or differentiate in linear patterns. To the authors' knowledge, these distinctively different clonal patterns have not been reported in airway basal cell literature, and further studies are warranted to elucidate the mechanisms behind the differences in order to gain further insight into the nature of airway repair and regeneration.
It has been shown previously that after treatment with the same LV vector used in this study, the cell types expressing the transgene changed over time, with the emergence of transgene-expressing goblet cells after 92 days. 5 In that study, the emergence of LacZ-expressing goblet cells was speculated to be due to transduced basal cells passing on the transgene during the natural cell turnover process. In the present study, histological analysis showed that in the short-term nasal control group, only LacZ-expressing airway basal cells and ciliated cells were present. In the long-term control group and PDOC groups, there were also LacZ-expressing goblet cells present, in accordance with the previous study. 5 These mucin secretory cells are not normally transduced by VSV-G pseudotyped vectors, suggesting that transduced airway basal cells pass the LacZ reporter gene to all their progeny upon differentiation, including to those cell types not initially targeted by the VSV-G pseudotype. The finding that transduction of airway basal cells results in transfer of the transgene to their progeny has important ramifications for provision of a single CFTR gene transfer that could provide lifelong therapeutic benefit to CF lungs. Whether this level of basal cell transduction is sufficient to provide lasting clinical benefit in a CF patient remains unclear.
KI-67 staining revealed there was a 2.1-fold increase in the number of actively cycling basal cells in the airways of CF mice compared to normal. Whether this is due to an increase in the division rate of a normal basal cell population or to a hyperplasia of basal cells dividing at a normal rate remains unknown. In either case, this finding provides further support for the premise that targeting these cells in the CF airways will elicit a sustained therapeutic response, as there are twice as many active cells to transduce than in normal airways. A question for future studies is whether a therapeutic correction of CF also returns the number of cycling basal cells to normal levels.
To ascertain if the findings observed in mice could be translated to human airway basal cells, primary human airway cells were obtained, airway basal cells were proliferated, and the cultures then treated with the LV-LacZ vector. Figure 6 shows that when treated with a MOI of 1:1, an average of 38.3% of cells were transduced, and nearly all cells expressed the marker gene at an MOI of either 10 or 100. A previous study reported that a F/HN pseudotyped SIV LV vector can transduce human airway basal cells at an efficiency of 47% when treated with a MOI of 100. 22 In another study using a LV-LacZ vector, treatment of basal cell cultures at MOIs of 100 and 500 showed a transduction efficiency of 54% and 82%, respectively. 23 The high basal cell transduction efficiency of the vector should reduce the amount of clinical grade vector that needs to be produced.
It is acknowledged that there is potential in isolating the basal cells from a patient, expanding and correcting them in vitro via gene therapy, and then re-administering them back into the patient. Recently, it was demonstrated that human tracheal stem/progenitor cells can engraft into the trachea of mice and contribute to restoring the airway epithelium after injury. 24 While the human stem/progenitor cells used in the study were transduced with LV particles, neither the efficiency of transduction nor the multiplicity of infection were clearly stated. The high efficiency of transduction observed with the vector at a low MOI makes it an ideal candidate to be used for the in vitro correction of isolated cells prior to engraftment back into the airways.
This study has some limitations. Firstly, it was not designed to analyze quantitatively the number of progenitor cells that were transduced. The LacZ marker gene was chosen, since it allows for the clear and immediate en face observational assessment of the patterns of cluster formation that resulted from transduction of basal cells and clonal outgrowth. In the future, a separate study could be designed to use fluorescence-activated cell sorting to determine the number of basal cells transduced with a fluorescent reporter, but this would not allow the en face visualization of outgrowth, which was the key outcome for this study. Second, the decalcification and Xgal processing of the tissue for visualization of LacZ expression in mouse nasal airways prevented subsequent immunohistochemical analyses to identify basal cells using the markers K5 and/or NGFR. So, only histological methods could be employed to identify the transduced cell types. However, despite these limitations, an epithelial injury model has been used to show for the first time that transduced airway basal cells pass the transgene onto their progeny, that there are more actively cycling basal cells in the CF mouse nasal airway, and that the vector can efficiently transduce human airway basal cells.
Conclusion
This study shows that airway basal cells can be transduced with a LV-LacZ vector, and that the transgene is passed onto their progeny. It has been shown previously that airway conditioning with LPC prior to LV delivery can produce long-term gene expression, and this study suggests that the mechanism of persistence may in part rely on clonal expansion of transduced basal cells. A linear pattern of clonal expansion has also been described for the first time, and different rates of basal cell cycling in CF and normal mice have been demonstrated. When applied to human airway basal cells in vitro, this vector has a very high efficiency of transduction, making it a suitable candidate for cell or gene therapy clinical development.
Footnotes
Acknowledgments
Studies supported by the U.S. CF Foundation, Robinson Research Institute (Centre for Stem Cell Research grant), NHMRC (project grant APP1098127), and philanthropic donors via the Cure 4 Cystic Fibrosis Foundation (
Author Disclosure
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.