
Abstract
A previous study described the important regulatory roles of microRNAs (miRNAs) in ischemic stroke. However, the functional significance of long non-coding RNA (lncRNAs) in ischemic stroke was largely unknown. This study aimed to identify lncRNA profiling and elucidate the regulatory mechanisms in the pathophysiology of stroke. RNA sequencing was performed on the blood of three ischemic stroke patients and three normal controls. Differential expression analysis was used to identify differentially expressed lncRNAs (DElncRNAs) and mRNAs (DEmRNAs). After further correlation and co-expression analysis, the corresponding co-expression networks and miRN–lncRNA–mRNA interaction network were then constructed. The expression of DElncRNAs and DEmRNAs was verified in Gene Expression Omnibus. RNA sequencing and subsequent bioinformatics analysis produced a total of 61 DElncRNAs (14 upregulated and 47 downregulated) and 673 DEmRNAs (432 upregulated and 241 downregulated). LOC105372881 and LOC101929707 were the most highly increased and decreased lncRNAs in ischemic stroke. LncRNA–mRNA co-expression networks were constructed according to 3,008 positively co-expressed and 607 negatively co-expressed lncRNA–mRNA pairs. The DElncRNAs may play roles in the pathways of glycolysis/gluconeogenesis, arrhythmogenic right ventricular cardiomyopathy, adherens junction, lysosome, and hematopoietic cell lineage by regulating their co-expressed mRNAs. Combined with previous data, a miRNA–lncRNA–mRNA interaction network for ischemic stroke was constructed. Based on GSE22255, the expression of six DElncRNAs (CEBPA-AS1, LINC00884, HCG27, MATN1-AS1, HCG26, and LINC01184) and 11 DEmRNAs (TREML4, AHSP, PI3, TESC, ANXA3, OAS1, OAS2, IFI6, ISG15, IFI44L, and LY6E) was similar to the current sequencing data. This study is the first to identify blood lncRNAs in human ischemic stroke using RNA sequencing. The findings may be the foundation for understanding the potential role of lncRNAs in ischemic stroke.
Introduction
I
Following an ischemic stroke, global mRNA expression profiles are altered extensively in the blood or brain. 5 Moreover, the expression profiles of non-coding RNAs (ncRNAs) are altered rapidly after ischemic stroke, including microRNAs (miRNAs), long non-coding RNA (lncRNAs), and piwi-interacting RNAs (piRNAs). 5 The different classes of ncRNAs have distinct regulatory functions, which regulate the processes of transcription and translation. 6 Mature miRNAs, approximately 21–23 nucleotides in length, play a role in the regulation of degradation and/or translational repression of the transcript by direct effects target mRNAs. 7 A vast number of lncRNAs are transcribed in eukaryotic cells. The lncRNAs are a class of ncRNAs >200 nucleotides in length, which are widely involved in gene transcription regulation and which act as epigenetic modifiers. 8 Recent studies have also demonstrated that dysregulation of ncRNAs can modulate the pathophysiology of central nervous system disorders, including focal ischemia. 9 –11 However, up to now, the functional significance of ncRNAs has not been completely elucidated, and recent studies have focused on the role of ncRNAs in the regulation of post-transcriptional gene expression and of responses to ischemia and neuroprotection lncRNAs. 5
Several studies have demonstrated that stroke alters the miRNA expression profiles in the brain and blood of human subjects and rodent models. 6,12 –14 Moreover, focal ischemia extensively alters the lncRNA expression in the rat brain following transient middle cerebral artery occlusion (MCAO). It has been suggested that stroke-induced lncRNAs may interact with chromatin-modifying proteins (CMPs), and two CMPs, Sin3A and coREST, have been examined. 11,15 Recently, Dykstra-Aiello et al. compared the lncRNA expression in the whole blood of ischemic stroke and normal controls by lncRNA microarray. They found that linc-SLC22A2 and linc-OBP2B-1 may relate to male ischemic stroke patients, and OTTHUMT00000079682 may play a role in female ischemic stroke patients. 16 In addition, common differentially expressed lncRNAs (DElncRNAs) between both sexes were also identified.
A previous study documented the important regulatory roles of miRNAs in ischemic stroke by integrated analysis of the published miRNAs and mRNAs data deposited in the Gene Expression Omnibus (GEO) database. 17 To identify the functional significance of lncRNAs (another class of ncRNAs) in the regulation of pathophysiology of ischemic stroke, the present study used RNA sequencing technology, which is superior to microarray, to profile lncRNA and mRNA expressional signatures in three pairs of ischemic stroke patients and three healthy controls. Due to the restriction of the sample size, it was difficult to identify the differentially expressed mRNAs (DEmRNAs) and DElncRNAs in male ischemic stroke and female ischemic stroke, respectively. Hence, the study focused on those DElncRNAs and DEmRNAs of ischemic stroke, irrespective of sex differences. Based on previous results and the identified differentially expressed lncRNAs/mRNAs in the current study, the lncRNA–mRNA co-expression network and miRNA–lncRNA–mRNA interaction network were constructed to elucidate the underlying regulatory mechanism of ncRNAs in ischemic stroke development.
Materials and Methods
Patient sample preparation
The current study included three ischemic stroke patients and three healthy controls of Asian ancestry between 52 and 73 years of age (Table 1). The etiology of stroke was classified according to the TOAST classification criteria. The three cases were patients with pathologically confirmed ischemic stroke diagnosed at the First Affiliated Hospital of Shantou University Medical College. The patient demographics, clinical features, and risk factors were extracted from medical records (Table 1). The three controls were randomly selected from healthy individuals who donated bloods samples and completed the questionnaire on medical histories. Peripheral venous blood was collected from each of the participants and used for RNA extraction. This study was approved by the Institutional Review Board of the First Affiliated Hospital of Shantou University Medical College. Signed informed consent was obtained from all of the study subjects.
Subject characteristics
NIHSS, National Institutes of Health Stroke Scale; TG, triglycerides; HDL-C, high-density lipoprotein cholesterol; LDL-C, low-density lipoprotein cholesterol.
RNA sequencing
Total RNA was isolated and purified using standard TRIzol Reagent (Invitrogen, Carlsbad, CA) according to the manual. The RNA concentration and purity were assessed using the Nanodrop2000 spectrophotometer (Thermo Fisher Scientific, Wilmington, DE). RNA integrity was further assessed using an Agilent 2100 Bioanalyzer (Agilent Technologies, Santa Clara, CA). After removing the ribosomal RNA, the cDNA library was constructed using the Truseq RNA sample Prep Kit (Illumina, Inc., San Diego, CA). Subsequent sequencing (2 × 150 bp) was conducted on the Illumina HiSeq 4000 sequencing system (Illumina, Inc.) according to the manufacturer's instructions. Library construction and RNA sequencing were performed by BGI (Beijing, China).
Bioinformatics analysis
About 36 million raw RNA sequencing data were obtained for each sample. First, quality-control filtering was performed by removing the adapter sequences and the low-quality bases using the FastQC program, and high-quality clean reads were generated. The clean reads were then mapped to human reference genome GRCh38 by tophat v2.1.1. The lncRNA and mRNA annotation files were downloaded from NCBI (
Differential expression analysis
To identify differentially expressed lncRNAs or mRNAs between ischemic stroke patients and the controls, differential expression analyses were performed using cuffdiff. A p-value of <0.05 was selected as the criteria for significantly differentially expressed lncRNA (DElncRNA) or mRNAs (DEmRNA). In order to obtain an overview of the characteristics of lncRNA and mRNA expression profiles, heat maps were further generated by hierarchical clustering analysis based on the normalized values of all DElncRNAs and the top 100 DEmRNAs using R package.
Correlation and co-expression analysis
The functions of most lncRNAs were unknown. Construction of a lncRNA–mRNA co-expression network was a better way to understand the functional forecast of lncRNAs. First, Pearson's correlation coefficient (PCC) was calculated between the expression levels of each DElncRNA–DEmRNA pair in ischemic stroke patients and the controls. Then, the co-expression DElncRNA–DEmRNA pairs were selected, with an absolute value of PCC ≥0.90 and a p-value <0.05 for further network construction. Specifically, this study generated two generalized linear models: (1) positive co-expression of DElncRNA–DEmRNA pairs in which DElncRNA and DEmRNA expression were similar in ischemic stroke and the controls; and (2) negative co-expression of DElncRNA–DEmRNA pairs in which DElncRNA and DEmRNA expression were different in ischemic stroke patients and the controls. Accordingly, LncRNA–mRNA co-expression networks were constructed based on the DElncRNA–DEmRNA co-expression associations, which were further visualized by Cytoscape 3.1 (
Delineation of functional annotation
Considering that lncRNAs execute functions by regulating the expression of the neighboring or overlapping coding genes, the biological function of the DEmRNAs needs to be identified, which were co-expressed with DElncRNA in ischemic stroke and the controls. Therefore, gene ontology (GO) and KEGG pathway analysis were conducted using the online tool GeneCoDis3 (
Construction of miRNA–lncRNA–mRNA interaction network
In a previous study, a total of 445 miRNA–mRNA target pairs were identified by integrated analysis of two mRNA and one miRNA microarray expression profiles from the GEO database. 17 To understand potential regulation relationships systematically among miRNAs, lncRNAs, and mRNAs in ischemic stroke, a miRNA–lncRNA–mRNA interaction network was constructed in the current study using the previous 445 miRNA–mRNA target pairs and the co-expression DElncRNA–DEmRNA pairs. The network was visualized using Cytoscape 3.1.
Cross-validation
GSE22255, a previously published GEO data set including 20 ischemic stroke cases and 20 normal controls, was downloaded from the GEO database. All 40 participants were Portuguese. Data for GSE22255 were obtained using GPL570 (HG-U133_Plus_2) Affymetrix Human Genome U133 Plus 2.0 Array. The background correction, normalization, and summarization were performed using the robust multi-chip average algorithm. Partek software was used to derive the adjusted data. 18 The log2 transformation to the intensities of lncRNAs and mRNAs was extracted from the GSE22255 data. Based on these data, differential expression analyses of mRNAs and lncRNAs in GSE22255 were performed using a t-test. Individual p-values and log2 values (fold change) were obtained. Then, the expression changes of selected DEmRNAs and DElncRNAs between ischemic stroke patients and normal controls in the RNA sequencing results was validated by GSE22255.
Results
Overview of the transcriptome profiling
To determine the lncRNA expression profiles in human ischemic stroke, three patients with ischemic stroke and three healthy controls were recruited. Their blood RNAs were prepared for sequencing using RNA sequencing technology. After quality-control filtering of the raw sequencing reads, 73099660, 71705392, and 73137640 clean reads were generated for three ischemic stroke patients, and 72009940, 73026418, and 73035222 clean reads were generated for three controls. Clean reads (>89%) could be mapped to the human reference genome GRCh38. In total, 11,477 lncRNAs and 19,576 mRNAs were identified. Principal components analysis (PCA) of the RNA sequencing data demonstrated global separation of ischemic stroke patients from normal controls (Supplementary Fig. S1; Supplementary Data are available online at
Next, the transcript expression level (FPKM) was calculated, and differential expression analysis was performed to identify the DElncRNAs and DEmRNAs between ischemic stroke patients and the controls. With the level of significance set at p < 0.05, 61 lncRNAs were differentially expressed in ischemic stroke, including 14 upregulated and 47 downregulated DElncRNAs. Among these 61 DElncRNAs, only 24 DElncRNAs had descriptions; the other 37 DElncRNAs were uncharacterized. Hence, the biotype of these 24 DElncRNAs was analyzed. Five DElncRNAs were antisense lncRNA, two DElncRNAs were sense-overlapping lncRNAs, and 17 DElncRNAs were intergenic lncRNAs. The full list of DElncRNAs, their biotype, and the loci with genes are displayed in Supplementary Table S1. With the level of significance set at p < 0.05, a total of 673 DEmRNAs were identified in ischemic stroke, including 432 upregulated DEmRNAs and 241 downregulated DEmRNAs. Unsupervised hierarchical clustered heat maps of the expression of DElncRNAs and DEmRNAs in ischemic stroke are displayed in Fig. 1. Based on Fig. 1, the expression of these 61 DElncRNAs clustered all samples into the ischemic stroke group and normal control group, respectively (Fig. 1A); the expression of the top 100 DEmRNAs clustered all samples into the ischemic stroke group and normal control group as well (Fig. 1B). Despite there being little heterogeneity in each group, it was found that the DElncRNAs and DEmRNAs obtained in this study could distinguish the ischemic stroke group from the normal control group. Among the top 10 DElncRNAs, LOC105372881 and LOC101929707 were the most significantly up- and downregulated DElncRNA in ischemic stroke, respectively (Table 2). Moreover, TREML4 and OAS1 were the most significantly up- and downregulated mRNAs in ischemic stroke, respectively (Table 3). Furthermore, these DElncRNAs were distributed in all chromosomes, except for chromosomes 8, 10, and 13, and the sex chromosomes X and Y, while the DEmRNAs were widely distributed in all chromosomes, except for the sex chromosome Y (Fig. 2).
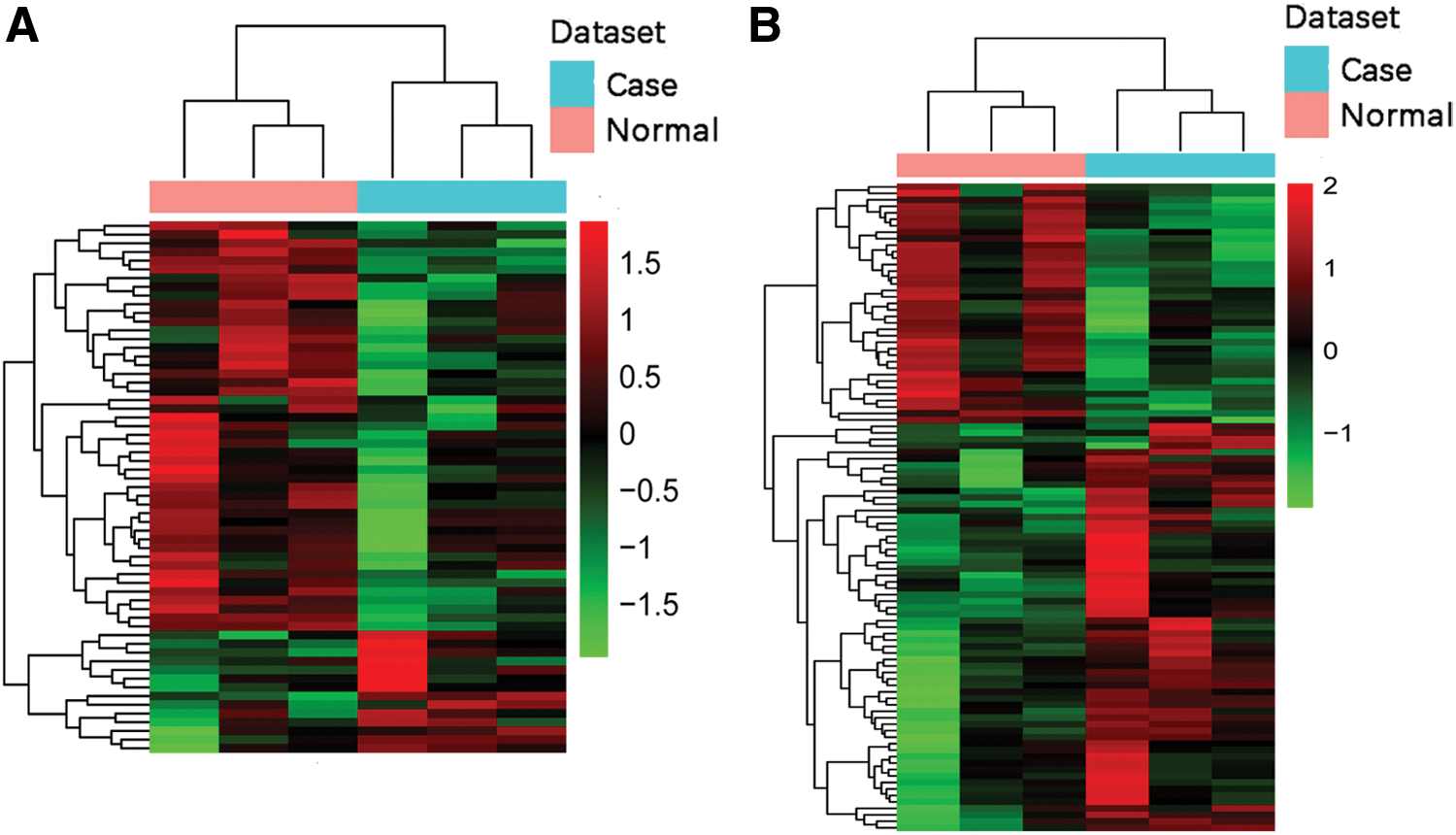
Unsupervised hierarchical clustering analysis showing expression profiles of differentially expressed long non-coding RNAs (DElncRNAs)
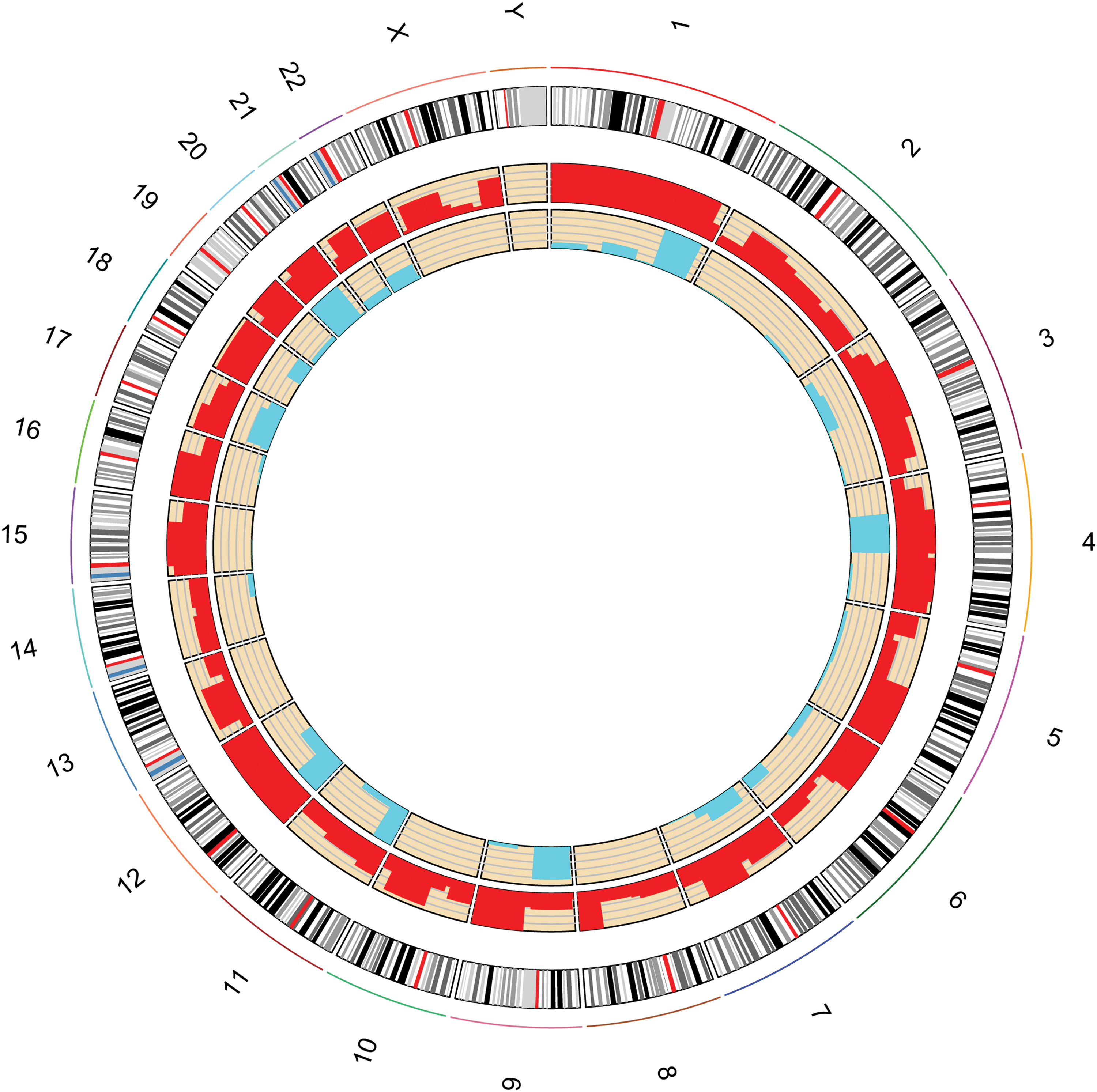
Circus plots representing the distribution of DElncRNAs and DEmRNAs on chromosomes. The outer circle is the chromosome map of the human genome hg19. The red inner circle and blue inner circle represent the distribution of DEmRNAs and DElncRNAs on different chromosome, respectively. Color images available online at
Top 10 up- and downregulated lncRNAs in ischemic stroke
Top 10 up- and downregulated mRNAs in ischemic stroke
Co-expression of lncRNAs and mRNAs in ischemic stroke
Considering that the functions of most lncRNAs have been unknown, lncRNA–mRNA co-expression analysis was performed to understand the roles of lncRNAs according to their co-expressed mRNAs. By calculating the PCC of 61 DElncRNAs and 673 DEmRNAs, a total of 3,008 lncRNA–mRNA pairs that were positively co-expressed and 607 lncRNA–mRNA pairs that were negatively co-expressed were identified. Based on the two distinct co-expression patterns of lncRNA–mRNA associated with ischemic stroke, corresponding lncRNA–mRNA co-expression networks were then constructed (Fig. 3). In the positive co-expression network, those 3,008 lncRNA–mRNA pairs were represented by 61 unique lncRNAs and 561 unique mRNAs. The top five lncRNAs with the largest numbers of associations were LINC00482, LINC00884, LOC105377826, LOC105369319, and LOC101928007. Moreover, the negative co-expression network contained 3,008 lncRNA–mRNA pairs, involving 54 lncRNAs and 286 unique mRNAs. The top five lncRNAs with the largest numbers of associations were LINC01215, LOC100996741, LOC105377809, LOC102724850, and LOC105376244.

Positive co-expression network

Functional characteristics of the identified lncRNAs
To predict the functions of DElncRNAs identified in this study better, functional annotation of their co-expressed mRNAs was performed. It is intriguing to note that the mRNAs positively co-expressed and negatively co-expressed with lncRNAs were both enriched in the pathways of glycolysis/gluconeogenesis, arrhythmogenic right ventricular cardiomyopathy, adherens junction, lysosome, and hematopoietic cell lineage.
Moreover, the 561 mRNAs positively co-expressed with lncRNAs were specially implicated in functions including type II diabetes mellitus, NOD-like receptor signaling pathway, hepatitis C, insulin signaling pathway, measles, and osteoclast differentiation (Fig. 4A). The 286 unique mRNAs that were negatively co-expressed with lncRNAs were specially enriched in malaria, phagosome, asthma, cytokine–cytokine receptor interaction, systemic lupus erythematous, Staphylococcus aureus infection, and tuberculosis (Fig. 4B).
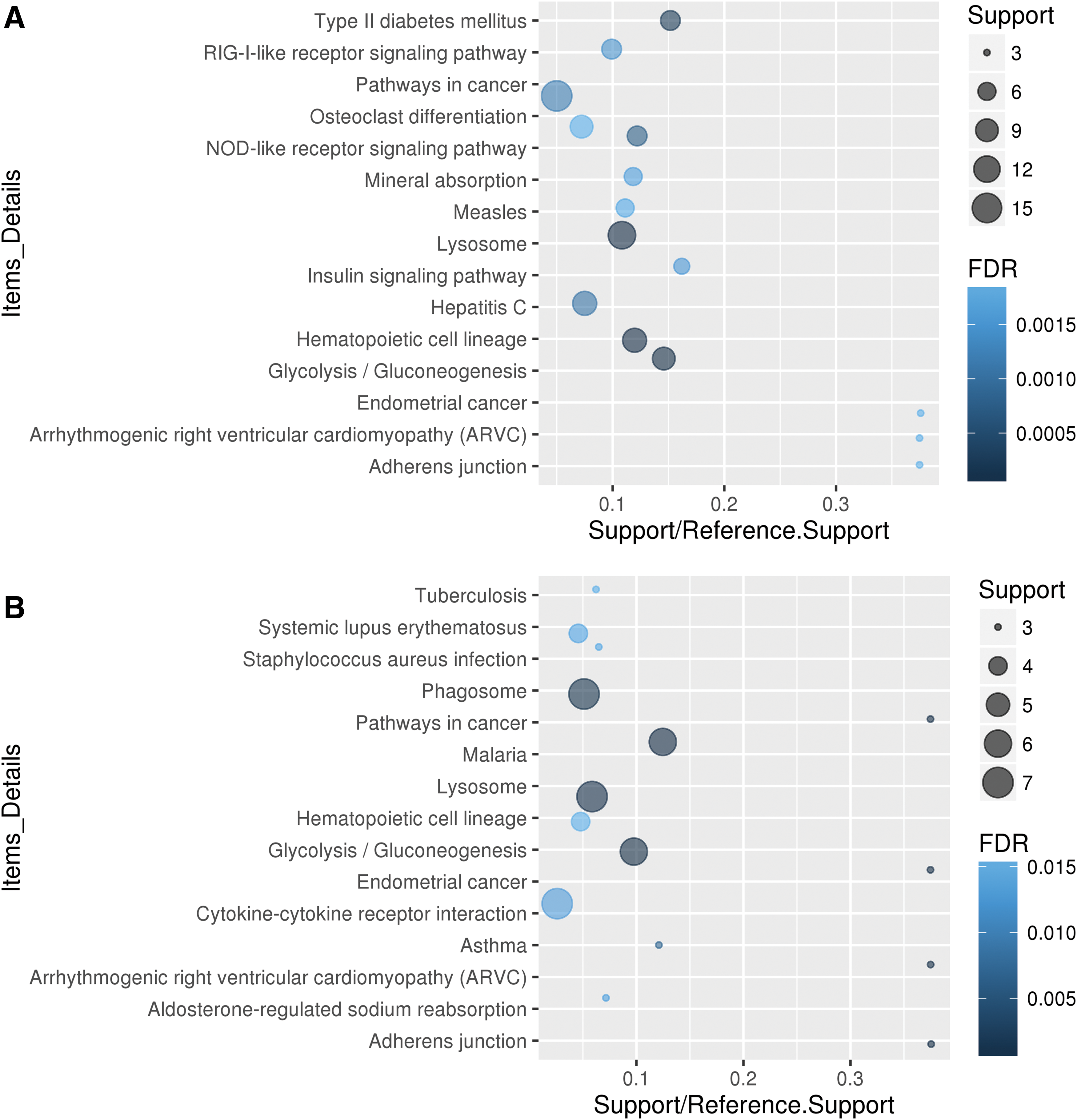
Top 15 enriched KEGG pathways for DEmRNAs showing positively co-expressed
MiRNA–lncRNA–mRNA interaction network
Using the 445 miRNA–mRNA target pairs from a previous study and 3,615 lncRNA–mRNA co-expression pairs, a miRNA–lncRNA–mRNA interaction network was constructed (Fig. 5). In the network, there were 12 miRNAs (9 upregulated and 3 downregulated), 32 lncRNAs (10 upregulated and 22 downregulated), and 13 mRNAs (7 upregulated and 6 downregulated). LINC00884, LOC105377826, LOC105376781, and LINC00482 were the hub lncRNAs with the largest numbers of associations in the miRNA–lncRNA–mRNA interaction network. The relation of 32 DElncRNAs and ischemic stroke has not been reported previously, which is consistent with the fact that little is currently known about the function of lncRNAs in ischemic stroke. Hence, the functions of DElncRNAs were predicted according to their co-expressed mRNAs.

The miRNA–lncRNA–mRNA interaction network in human ischemic stroke. Rectangles represent miRNAs, ellipses represent lncRNAs, inverted triangles represent mRNAs. Red and blue indicate upregulation and downregulation. Yellow and green edges indicate positive and negative co-expression of lncRNA and mRNA, respectively. Color images available online at
Among the upregulated mRNAs, a variant of rs864745 within JAZF1 was responsible for arteriolosclerosis, 19 and JAZF1 was co-expressed with LINC01094, CEBPA-AS1, LOC102724850, and LINC00672. IRAK3 was involved in the key pathway of transforming growth factor beta signaling in hemorrhagic transformation of human ischemic stroke, 20 and IRAK3 was co-expressed with LOC105377826, LOC105376781, LOC105369319, LOC101928007, LINC00884, and LINC00482. GADD45A was altered in the hippocampus of ischemic mice, 21 and GADD45A was co-expressed with LOC105377826, LOC105376781, LOC105369319, LOC101928007, LINC00884, and LINC00482.
Moreover, among the downregulated mRNAs, LEF1 was required for sprouting angiogenesis, reconstruction of the basal lamina, and the resolution phase in the post-stroke angiogenesis in the aged brain. 22 LEF1 was co-expressed with LOC105377809, LOC105370660, LOC102723446, LINC01215, LINC01184, LINC01128, LOC105377826, LOC105376781, LOC105376244, and LINC00884. Mutations in KLHL3 cause pseudohypoaldosteronism type II PHAIID, which is characterized by hypertension—an important risk factor for stroke. 23 KLHL3 was co-expressed with LOC105371655, LOC102723446, LINC01336, LINC01184, LINC01128, and LOC105376781. EphA4 receptors were necessary for the Ephrin-A3 reverse signaling, which regulated hippocampal neuronal damage and astrocytic glutamate transport after transient global ischemia. 24 EPHA4 was co-expressed with LOC105370660, LINC01128, and LOC105376244.
Validation in the GEO data set
The expression pattern of 61 DElncRNAs and 11 selected DEmRNAs was verified using the GSE22255 data set. RNA sequencing is superior to microarray for characterizing transcriptomes. The DEmRNAs and DElncRNAs of ischemic stroke in this study were obtained by RNA sequencing performed on an Illumina platform. The expression profiles of DEmRNAs and DElncRNAs were based on counts of reads mapped to human reference genome GRCh38. However, data for GSE22255 were obtained by GPL570 (HG-U133_Plus_2) Affymetrix Human Genome U133 Plus 2.0 Array. 18 The expression profiles of DEmRNAs and DElncRNAs were based on probe intensities. The majority of probes in this array were probes for mRNA; few probes for lncRNA can be found in this array. Hence, the probes for lncRNA in this array were not sufficient to detect all the targets in this study. Only 15 DElncRNAs were detected in GSE22255. The p-value and log2 (fold change) of selected DEmRNAs and DElncRNAs in GSE22255 are displayed in Supplementary Table S2. According to Supplementary Table S2 and Fig. 6, two DElncRNAs (CEBPA-AS1 and LINC00884) and five DEmRNAs (TREML4, AHSP, PI3, TESC, and ANXA3) were upregulated, while four DElncRNAs (HCG27, MATN1-AS1, HCG26, and LINC01184) and six DEmRNAs (OAS1, OAS2, IFI6, ISG15, IFI44L, and LY6E) were downregulated in ischemic stroke with no significant difference. Although these 11 DEmRNAs and 6 DElncRNAs were not significantly expressed between ischemic stroke and normal control in GSE22255, the trends of expression change of these DEmRNAs and DElncRNAs between ischemic stroke and normal control in GSE22255 were similar in the RNA sequencing results.

Validation of the DElncRNAs
Discussion
ncRNAs have various biological functions and act as important regulators of gene expression at the levels of transcription, RNA processing, and translation. 25 Recent studies have begun to focus on the significant functions of ncRNAs in regulating transcription and translation in maintaining normal cellular physiology. 6,26 The dysregulation of ncRNAs causes them to act as regulators of the pathophysiology of cerebral vascular endothelium and responses to the brain's ischemia and neuroprotection. 5,27 lncRNAs are a class of ncRNAs with >200 nucleotides, 28 and the dysregulation of spatiotemporal expression profiles can contribute to pathologies of the central nervous system. 9 Dharap examined the effect of ischemia on lncRNA expression in experimental rat models after MCAO. 11 Indeed, a few dysregulated lncRNAs, such as CDKN2B-AS1, 29 CDKN2B-AS10, 30 and CDKN2B-AS11, 31 have been linked to stroke. However, the alteration of lncRNA expression in human patients suffering ischemic stroke and their functional significance in ischemic stroke remain largely unknown. By using microarray analyses, a previous study obtained the lncRNA expression profile of ischemic stroke patients who were mostly white. This study used the RNA sequencing approach to obtain lncRNA expression profiles for Asian patients with ischemic stroke and normal controls. Compared to normal controls, a total of 61 ischemic stroke-associated DElncRNAs and 673 DEmRNAs were identified. Maybe due to the racial difference and different approaches for identifying the DElncRNAs in these two studies, the DElncRNAs in ischemic stroke obtained in this were quite different from the previous study. The majority of DElncRNAs in ischemic stroke obtained in this study were found to be dysregulated in ischemic stroke. Besides, four common DElncRNAs (LINC00174, LOC101927156, LOC102723446, and LOC105376244) in ischemic stroke in both studies were obtained according to their start–end locations on chromosomes. It was speculated that these four DElncRNAs may be key lncRNAs in ischemic stroke, despite racial differences.
In addition, the expression validation of 11 DEmRNAs and 6 DElncRNAs in ischemic stroke obtained in the RNA sequencing results was conducted by a microarray in GSE22255. Although these DEmRNAs and DElncRNAs were not significantly expressed between ischemic stroke and normal control in GSE22255, the trends of expression change of these DEmRNAs and DElncRNAs between ischemic stroke and normal control in GSE22255 were similar to the RNA sequencing results, which provided evidence for this study. Since the RNA sequencing results were based on Chinese patients, while the GSE22255 data were derived from Portuguese patients, it was speculated that the difference in expression between GSE22255 and the RNA sequencing results may be due to racial differences.
Up to now, the functions of lncRNA in the brain were mostly studied using an experimental rat model after MCAO. 11,15 The emerging evidence for function of most lncRNAs was lacking understand of the regulatory mechanism of ischemic stroke. Consistent with this, the roles of DElncRNAs identified in the current study were almost unknown. First, the strategy of constructing a co-expression network of lncRNAs and mRNAs 32 was used to research the biological functions of DElncRNAs in ischemic stroke in more depth. Two different co-expression patterns—positive and negative co-expression networks—related to ischemic stroke were found, which suggested distinct underlying regulation mechanisms. The positive co-expression network contained 3,008 DElncRNA–DEmRNA pairs involving 61 DElncRNAs and 561 DEmRNAs, while the negative co-expression network contained 3,008 lncRNA–mRNA pairs involving 54 DElncRNAs and 286 DEmRNAs.
Further functional annotation of the DEmRNAs co-expressed with the DElncRNAs indicated that several common pathways were enriched, including glycolysis/gluconeogenesis, arrhythmogenic right ventricular cardiomyopathy, adherens junction, lysosome, and hematopoietic cell lineage. Previous studies have indicated that cardiomyopathy is the predominant manifestation of a neuromuscular disorder, and ischemic stroke is the most frequent neurological disease induced by cardiomyopathies. 33 Ischemic stroke was reported to be a neurological complication of arrhythmogenic right ventricular cardiomyopathy. 34 –37 Hence, arrhythmogenic right ventricular cardiomyopathy was speculated to be associated with ischemic stroke, while the precise role of arrhythmogenic right ventricular cardiomyopathy in ischemic stroke needed further research.
A previous study suggested that cerebral ischemia may enhance hepatic gluconeogenesis, contributing to post-stroke hyperglycemia, 38 whereas the adherens junction was involved in long-term stroke protection, 39,40 and lysosome was involved in the neuronal death and survival after ischemic stroke. 41 It is noteworthy that hematopoietic lineage cells possess therapeutic potential against ischemic stroke. 42 Another constructed network in the current study was the miRNA–lncRNA–mRNA interaction network, which was used to survey further the significant dysregulated lncRNAs and mRNAs. In the miRNA–lncRNA–mRNA interaction network, LINC00884, LOC105377826, LOC105376781, and LINC00482 had the most associations. Three lncRNAs, LOC105370660, LINC01128, and LOC105376244, were co-expressed with EPHA4. EPHA4 regulated by miRNA145 was reported to regulate the damage of hippocampal neuronal and glutamate transport in astrocytes in transient cerebral ischemia 43 and motor function after ischemic stroke by interactions with downstream target genes. 44 Previous studies have reported that dysfunction of miRNA145 was associated with the pathologies of ischemic stroke by regulating angiogenesis, hyperlipidemia, nerve injury, and inflammatory reaction. 1,6,45 –52 The mechanism may include multiple molecular signaling pathways and interactions with lncRNAs and mRNAs. Hence, it is speculated that the miR145–lncRNAs (LOC105370660, LINC01128, and LOC105376244)–EPHA4 interaction may play an important role in ischemic stroke. Moreover, LOC105376244 was a commonly identified DElncRNA in ischemic stroke in both this present study and a previous ischemic stroke-related microarray study, which emphasized its importance in ischemic stroke. Besides EPHA4, another stroke-related gene, LEF1, was co-expressed with LOC105376244 as well. These findings provide clues for exploring the functions of LOC105376244 in ischemic stroke.
In additional, other major connected mRNAs have been known to be associated with ischemic stroke, such as JAZF1, IRAK3, GADD45A, and KLHL3. 19 –24 As another common DElncRNA in both this study and a previous ischemic stroke-related microarray study, LOC102723446 was co-expressed with LEF1 and KLHL3. It is speculated that LOC102723446 may be involved in ischemic stroke by regulating the expression of LEF1 and KLHL3. Besides LOC102723446, it was deduced that other lncRNAs co-expressed with the above mRNAs may play roles in the pathology of ischemic stroke by regulating co-expression genes. To the authors' knowledge, the miRNA–lncRNA–mRNA interaction network presented in this study is the first of its kind for human ischemic stroke, which provides clues for exploring the function of DElncRNAs in ischemic stroke.
Taken together, this study identified the lncRNA and mRNA expression profiling in human ischemic stroke by high-throughput RNA sequencing technology. The data show that an abundance of novel DElncRNAs may be associated with the risk of ischemic stroke and post-stroke outcome. Moreover, the construction of a DElncRNA–DEmRNA co-expression network and DEmiRNA–DElncRNA–DEmRNA interaction network provided clues for exploring the biological functions of DElncRNAs in ischemic stroke. The precise role of these novel DElncRNAs in the pathogenesis of ischemic stroke needs to be clarified in future work. The present findings might provide the foundation for future research on the potential role of lncRNAs in ischemic stroke.
However, the sample size for RNA sequencing was small, which is a limitation of this study. Research with a larger sample size and further experiments need to be conducted to confirm this conclusion and to identify the key lncRNAs and genes in male and female ischemic stroke, respectively.
Footnotes
Author Disclosure
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.