
Abstract
In this study we propose a novel approach based on the use of mesenchymal stromal cells (MSCs), aiming at limiting risks of graft failure in gene therapy protocols associated with low conditioning regimens. Because the engraftment of corrected hematopoietic stem cells (HSCs) is particularly challenging in Fanconi anemia (FA), we have investigated the relevance of MSCs in an experimental model of FA gene therapy. Our results showed, first, that risks of graft failure in recipients conditioned with a moderate dose of 5 Gy and infused with limited numbers of wild-type HSCs are significantly higher in Fanca−/− recipients as compared with wild-type recipients. However, when wild-type HSC numbers inducing 30–50% of graft failures in Fanca−/− recipients were coinfused with MSCs, no graft failures were observed. Moreover, graft failures associated with the infusion of low numbers of gene-corrected Fanca−/− HSCs were also significantly overcome by MSC coinfusion. Our study shows for the first time that MSC coinfusion constitutes a simple and nontoxic approach to minimize risks of graft failure in gene therapy applications associated with low conditioning regimens and infusion of limited numbers of corrected HSCs.
Introduction
I
MSCs are multipotent cells 3 –5 characterized by important immunoregulatory properties. 6 –8 On the basis of these properties, MSCs have been used to facilitate HSC engraftment in xenogeneic 9 –13 and allogeneic transplants. 14 –16 More recently we have demonstrated that MSCs also improve the engraftment of healthy donor HSCs in a mouse model of autologous HSC transplantation. 17 In those experiments we demonstrated that the simultaneous coinfusion of HSCs and MSCs was an essential requirement for improving the engraftment of donor HSCs, suggesting that MSCs could be acting as carriers of transplanted autologous HSCs to recipient bone marrow (BM) niches.
On the basis of our previous studies, here we have investigated the potential beneficial effect of MSCs in a clinically relevant model of autologous HSC gene therapy. In this respect, because of the limited number of HSCs present in the hematopoietic tissues of patients with Fanconi anemia (FA), 18 –20 and because none of the clinical trials so far conducted in patients with FA have shown engraftment of gene-corrected HSCs, 18,19 we have considered FA an ideal model to demonstrate the potential beneficial effect of MSCs in HSC gene therapy. In addition, because mutations in FANCA account for more than 60% of patients with FA, 21,22 a Fanca−/− mouse model was chosen for our studies.
Although myeloablative conditioning and infusion of high doses of corrected HSCs constitute the preferential options to guarantee the stable engraftment of gene-corrected HSCs, current conditioning protocols are associated with significant genotoxicity, particularly in DNA repair syndromes, such as FA. 23,24
Our data demonstrate for the first time that MSCs minimize risks of graft failure associated with moderate conditioning and infusion of limiting numbers of gene-corrected Fanca−/− HSCs. These results suggest that MSCs could offer similar benefits in clinical hematopoietic gene therapy.
Materials and Methods
Mice
FVB/NJ mice, aged 10–12 weeks, were housed and bred at the Centro de Investigaciones Energéticas, Medioambientales, y Tecnológicas (CIEMAT, Madrid, Spain) Animal Facility (Registration No. ES280790000183). Mice with a disruption in Fanca (Fanca−/− mice) 25 were back-crossed to obtain an FVB-Fanca −/− strain. Mice were routinely screened for pathogens in accordance with FELASA (Federation of European Laboratory Animal Science Associations) procedures, and received water (filtered [pore size, 50 μm]/ultraviolet-irradiated) and food (SAFE R04 25 kGy γ-irradiated) ad libitum. All experimental procedures were carried out according to Spanish and European regulations (Spanish RD 53/2013 and Law 6/2013 that transpose and fulfill the European Directive 2010/63/UE about the use and protection of vertebrate mammals used for experimentation and other scientific purposes). Procedures were approved by the CIEMAT Animal Experimentation Ethical Committee according to approved biosafety and bioethics guidelines.
Generation and characterization of mouse MSCs
To obtain MSCs, FVB mice were sacrificed, and adipose tissue from the epiploon was cut into small pieces, digested with collagenase A (1 mg/ml; Roche Diagnostics, Mannheim, Germany) in Dulbecco's modified Eagle's medium (DMEM) for 2 hr, and filtered (pore size, 0.40 μm). Samples were seeded in flasks (Corning, Corning, NY) at 1.6 × 105 cells/cm2 and cultured in supplemented MesenCult medium for mouse cells (STEMCELL Technologies, Vancouver, BC, Canada) at 37°C and 5% CO2. After 24 hr, nonadherent cells were discarded and fresh medium was added and replaced twice per week. When cells reached 80% confluence, adherent cells were trypsinized, washed, and seeded at a concentration of 4 × 103 cells/cm2. In all the experiments MSCs were used at passages 5–8.
Osteogenic and adipogenic MSC differentiation capacity was ensured by using StemXVivo osteogenic/adipogenic base media supplemented with StemXVivo osteogenic supplement or StemXVivo adipogenic supplement (R&D Systems, Minneapolis, MN), respectively, according to the manufacturer's instructions.
For immunophenotype analysis, MSCs were harvested with 0.05% trypsin–EDTA, washed, and resuspended in 1% bovine serum albumin (Sigma-Aldrich, St. Louis, MO)-supplemented phosphate-buffered saline (PBS). MSCs were incubated with monoclonal antibodies for 30 min at 4°C. Flow cytometric analyses included CD34, CD45.1, CD80, Sca-1, CD29, CD44, CD73, CD90, CD105, CD106, CD144, and CD166 expression (BD Biosciences Pharmingen, San Diego, CA). Results were analyzed with FlowJo (FlowJo, Ashland, OR).
Hematopoietic stem cell purification
Bone marrow cells were collected from tibias and femurs of wild-type and Fanca−/− male mice by flushing with DMEM. After red blood cell lysis, cells were resuspended in PBE (PBS containing 0.5% bovine serum albumin and 5 mM EDTA). Lineage−, Sca-1+, c-Kit+ (LSK) cells were obtained with lineage-specific antibodies (anti-Gr1, anti-B220, anti-CD3e, anti-CD11b, anti-Ter119) and anti-Sca-1 and anti-c-Kit antibodies (BD Biosciences, San Jose, CA), and sorted with a BD Influx cell sorter (BD Biosciences). After cell sorting, mean purities of 90.27 ± 12.20% LSK cells were obtained.
HSC lentiviral transduction
Fanca−/− LSK cells were transduced with a pre-GMP PGK-FANCA-wPRE* lentiviral vector (FANCA-LV) 26 produced by Généthon (Evry, France) at a multiplicity of infection (MOI) of 200 IU/ml, for 16 hr, at 37°C in StemSpan medium (STEMCELL Technologies), supplemented with mouse stem cell factor (mSCF; 50 ng/ml), human thrombopoietin (hTPO; 20 ng/ml), human Flt-3 ligand (hFlt-3L; 25 ng/ml), and human interleukin-3 (hIL-3; 10 ng/ml) and hIL-6 (25 ng/ml) (EuroBioSciences, Friesoythe, Germany) in RetroNectin (2 μg/cm2, Takara Bio, Otsu, Shiga, Japan)-treated plates for 4 hr.
Clonogenic assays
Colony-forming cell (CFC) assays were performed by plating 150 Fanca−/− or wild-type LSK cells in 35-mm plates with 1 ml of MethoCult GF M3534 (STEMCELL Technologies) containing increasing concentrations (from 0 to 100 nM) of mitomycin C (MMC). Samples were cultured for 7 days at 37°C, 5% CO2, and colonies of at least 50 cells were scored.
To determine the proportion of transduced Fanca−/− CFCs, individual colonies were picked after 14 days of culture and vector copy numbers (VCNs) per cell were analyzed by qPCR in the Rotor Gene RG-3000 (Corbett/Qiagen, Germantown, MD), using specific primers for the Psi sequence: Psi-F, 5′-CAGGACTCGGCTTGCTGAAG-3′ and Psi-R, 5′-TCCCCCGCTTAATACTGACG-3′, and detected with the TaqMan probe Psi-P, CGCACGGCAAGAGGCGAGG-FAM. For mouse genomic quantification, specific primers for the Titin gene were used: mTitin-F, 5′-AAAACGAGCAGTGACGTGAGC-3′ and mTitin-R, 5′-TTCAGTCATGCTGCTAGCGC-3′, and detected with the TaqMan probe mTitin-P, TGCACGGAAGCGTCTCGTCTCAGTC-Texas Red.
Hematopoietic stem cell transplants
Various doses of BM LSK cells were infused through the tail vein of female recipients irradiated with a sublethal dose of 5 Gy, using MG324 X-ray equipment (300 kV, 12.8 mA; Philips, Hamburg, Germany). In experiments corresponding to Figs. 2 and 4, purified LSK cells were infused as a sole population or mixed with MSCs (400,000–600,000 MSCs/mouse), as previously described. 17 To prevent differences between control and experimental groups associated with the circadian rhythm, irradiations and transplantations in mice corresponding to experimental and control groups were always done in parallel.
The engraftment of donor cells was periodically analyzed in samples from peripheral blood (PB) and hematopoietic tissues of transplanted female recipients by qPCR, using primers for the male-specific SRY sequence and the mouse β-actin in the Rotor Gene RG-3000 device (Corbett/Qiagen).
The frequency of repopulating cells in wild-type LSK cells determined after infusion into wild-type and FA recipients irradiated with 5 Gy was quantified with extreme limiting dilution analysis (ELDA) software. 27
Cell cycle analysis
Lineage-negative BM cells were selected by magnetic-activated cell sorting (MACS; Miltenyi Biotec, Bergisch Gladbach, Germany), resuspended in Hoechst binding buffer (HBB, Hanks' balanced salt solution containing 20 mM HEPES, 5.5 mM glucose, and 10% fetal bovine serum [FBS]), and stained with Hoechst 33342 (Sigma-Aldrich) for DNA, and pyronin Y (Sigma-Aldrich) for RNA, for 45 min at 37°C. Thereafter, cells were stained for LSK phenotype and analyzed with the BD Influx cell sorter (BD Biosciences).
Hematopoietic progenitor analysis
To study hematopoietic progenitor subpopulations, BM cells were stained for LSK phenotype with CD34 and CD135 (BD Biosciences) antibodies. Long-term HSCs (LT-HSCs) were defined as CD34−CD135− cells, short-term HSCs (ST-HSCs) as CD34+CD135− cells, and multipotent progenitors (MPPs) as CD34+CD135+ cells. Cells were analyzed with a BD Fortessa cell cytometer (BD Biosciences).
Statistical analysis
Statistical analysis was performed by one-way analysis of variance with a Bonferroni post-hoc test and by a nonparametric Mann–Whitney test. Results are expressed as means ± the standard error of the mean (SEM), and significant p values were ≤0.05. Statistical analysis was performed with GraphPad Prism 5.00 for Windows (GraphPad Software, San Diego, CA).
Results
The transplantation of limited numbers of wild-type HSCs in Fanca−/− recipients is associated with increased risks of graft failure
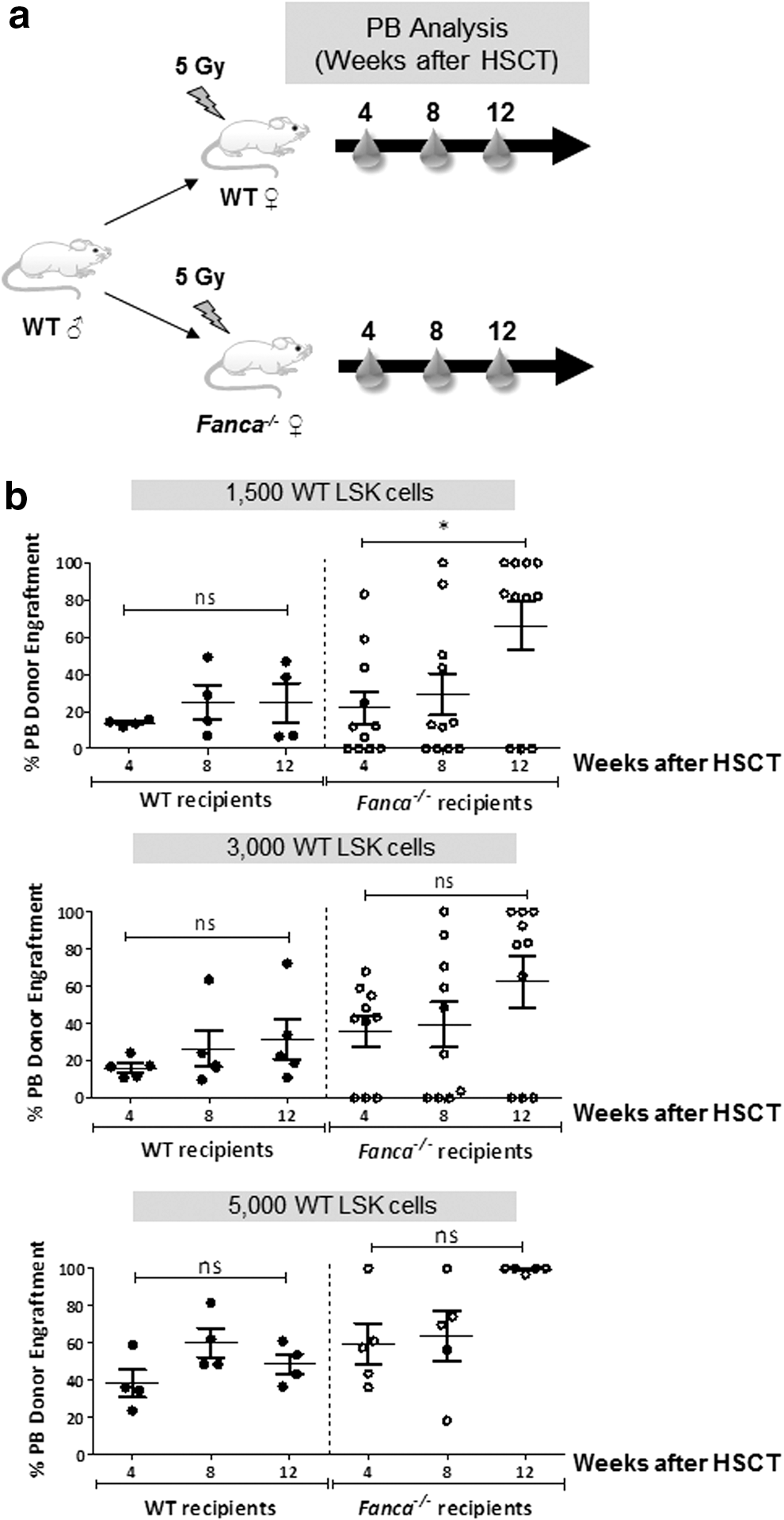
In a first set of experiments we investigated whether the use of Fanca −/− recipients had an impact in the engraftment of donor wild-type HSCs. With this aim, limited numbers of wild-type LSK cells were transplanted in parallel into wild-type and Fanca −/− mice previously irradiated with a submyeloablative dose of 5 Gy (Fig. 1a).
Comparative analysis of the engraftment ability of wild-type HSCs in Fanca
−/− and wild-type recipient mice.
Although doses as low as 1500 wild-type LSK cells were sufficient to confer stable engraftments in wild-type recipients, the infusion of either 1500 or 3000 wild-type LSK cells resulted in 30% graft failures in Fanca−/−
recipients (Fig. 1b). To quantify whether numbers of wild-type repopulating cells required to engraft 5 Gy-irradiated wild-type and FA recipients were statistically significantly different, an extreme limiting dilution analysis was conducted in recipients transplanted with 100–5000 LSK cells (see description of ELDA assays in Materials and Methods). Interestingly, the frequency of repopulating cells determined in FA recipients was three times lower as compared with frequencies determined in wild-type recipients (1/1826 LSK cells versus 1/595 LSK cells, respectively; p < 0.05; see Supplementary Table S1) (supplementary data are available online at
Strikingly, although the number of wild-type LSK cells required to prevent graft failures was significantly higher in FA than in wild-type recipients, a progressive increase in the proportion of donor cells was observed in FA recipients, but not in wild-type recipients (Fig. 1b).
Taken together, the results obtained in these experiments show that higher risks of graft failure are associated with the infusion of low numbers of HSCs into recipients with the Fanca −/− genotype. On the other hand, our data also demonstrate that when FA recipients are used, an evident proliferation advantage of donor-engrafted HSCs takes place, facilitating in the long term the predominant exogenous reconstitution of these recipients.
MSCs limit the risks of graft failure in Fanca−/− recipients transplanted with wild-type HSCs
To investigate whether graft failures observed in Fanca −/− recipients could be minimized by MSC coinfusion, 1500 wild-type LSK cells, either with or without 400,000–600,000 wild-type MSCs, were infused into Fanca −/− recipients irradiated with 5 Gy (see Fig. 2a). MSCs showed the expected fibroblast-like morphology, adipogenic and osteogenic differentiation capacity, and expressed the characteristic markers of these cells (positive for CD29, CD44, CD73, CD90.2, CD105, CD106, CD144, and CD166, and negative for the hematopoietic CD34, CD45.1, CD80 markers) (Supplementary Fig. S1).

Effect of MSCs on the engraftment of wild-type HSCs in Fanca
−/− recipients.
Consistent with data from Fig. 1, 30% of Fanca−/− recipients infused with 1500 wild-type LSK cells showed graft failure, whereas the remaining recipients showed progressive increases in donor repopulation as the posttransplantation time was extended (Fig. 2b). Although no statistically significant difference in the level of donor engraftment was observed in recipients transplanted with wild-type LSK cells with respect to those cotransplanted with wild-type LSK cells and MSCs, the coinfusion of MSCs with 1500 LSK cells prevented graft failures in transplanted recipient mice. These results are consistent with those obtained previously by our group in an autologous bone marrow transplantation (BMT) model, 17 and suggested the potential beneficial effect of MSCs in experimental models of FA gene therapy.
Gene correction improves the repopulating properties of Fanca −/− HSCs
Before evaluating the efficacy of MSCs in an HSC gene therapy model of FA, we determined differences in the phenotypic and repopulating properties of uncorrected Fanca −/− HSCs, as compared with wild-type HSCs. Decreased numbers of freshly harvested HSCs were observed in the BM of Fanca −/− mice, as compared with wild-type mice (Supplementary Fig. S2a-c). In addition, a higher proportion of LSK cells in the G1 phase of the cell cycle was observed in Fanca −/− mice compared with wild-type mice (Supplementary Fig. S2d and e), consistent with observations made in other FA mouse models. 28,29 Infused doses as high as 5000 Fanca −/− LSK cells were incapable of engrafting wild-type recipients (Supplementary Fig. S3a and b), contrasting our data with wild-type LSK cells, where doses as low as 1500 LSK cells engrafted 100% of wild-type recipients (Fig. 1b). These results indicate that Fanca −/− mice are characterized by the reduced presence of HSCs in BM, whose quiescent state is also reduced as compared with wild-type HSCs.
Finally, the results of Supplementary Fig. S3c show that the engraftment of donor Fanca −/− LSK cells was modestly enhanced when Fanca −/− recipients instead of wild-type recipients were used, probably due to the deficient repopulating properties of endogenous Fanca −/− HSCs.
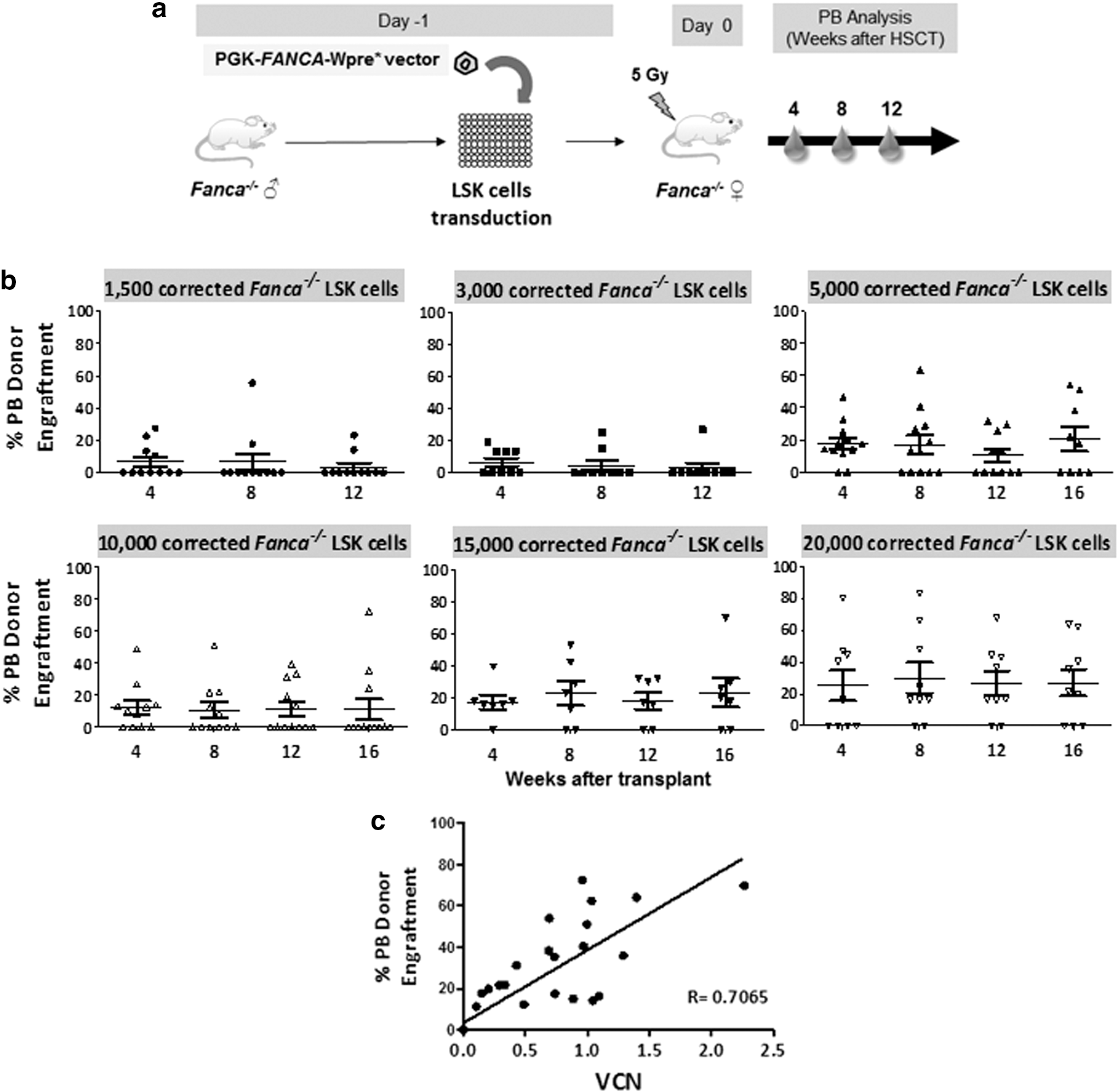
In subsequent experiments we moved to investigate risks of graft failure and evidence of repopulation advantage, using gene-corrected Fanca −/− LSK cells previously transduced with the PGK-FANCA-wPRE* lentiviral vector (Fig. 3a). qPCR analyses of individual colonies generated by gene-corrected Fanca −/− LSK cells showed transduction efficacies of 76.01 ± 0.17%, and mean VCN of 1.08 ± 0.28 per cell (data not shown). Clonogenic assays confirmed that transduction of Fanca −/− LSK cells with the therapeutic vector reversed the characteristic MMC hypersensitivity of Fanca −/− progenitor cells (Supplementary Fig. S4a), without inducing any significant toxicity in the CFCs (Supplementary Fig. S4b).
Analysis of the engraftment ability of gene-corrected Fanca
−/− HSCs in syngeneic Fanca
−/− recipients.
As shown in Fig. 3b, a high incidence of graft failure was observed when doses lower than 15,000 corrected Fanca −/− LSK cells were infused. Even after the infusion of 20,000 gene-corrected Fanca −/− LSK cells, 30% graft failures were observed. As shown in Fig. 3b, stable engraftments were noted along the posttransplantation period, although no evidence of in vivo proliferation advantage of gene-corrected Fanca −/− HSCs was observed. Analyses of the lentiviral VCN in PB cells of gene therapy-treated mice showed that VCNs correlated with donor engraftment levels (Fig. 3c), indicating that conclusions related to donor cell engraftment were applicable to the engraftment of gene-corrected cells.
Data corresponding to this set of experiments show that corrected Fanca −/− LSK cells repopulate Fanca −/− recipient mice in a dose-dependent manner, although they do not reproduce the efficient engraftment ability and proliferation advantage characteristic of wild-type LSK cells.
MSCs improve the engraftment and promote the repopulation advantage of gene-corrected Fanca −/− HSCs in Fanca −/− recipients
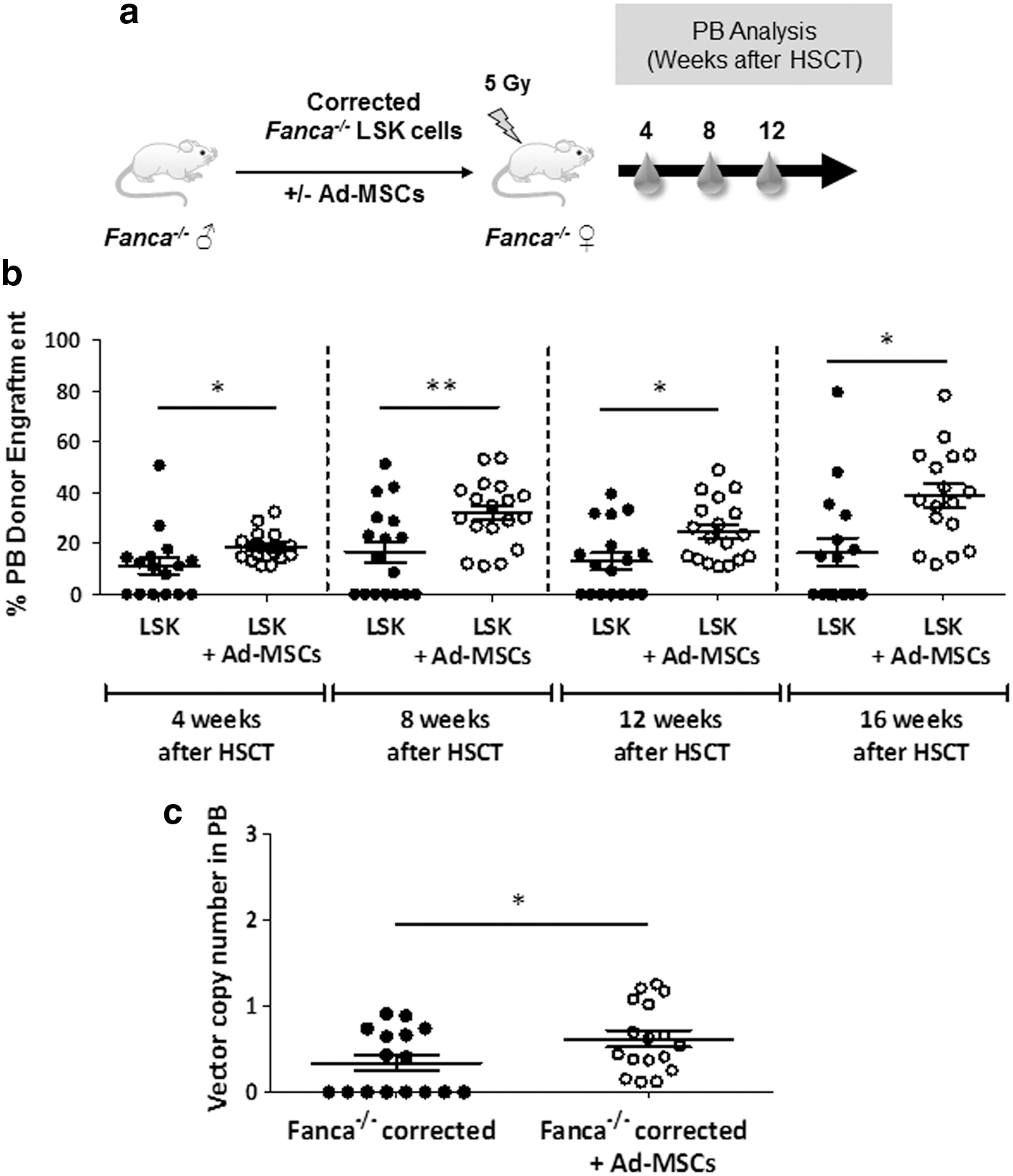
In a final set of experiments we investigated whether MSCs reduced graft failure risks associated with the infusion of limited numbers of corrected Fanca −/− HSCs in Fanca −/− recipients conditioned with 5 Gy. In these experiments, mice were transplanted with 10,000 gene-corrected Fanca −/− LSK cells, either as a unique population or coinfused with 400,000–600,000 MSCs (Fig. 4a).
Effect of MSCs on the engraftment of gene-corrected Fanca
−/− HSCs transplanted into Fanca
−/− recipients.
Consistent with data in Fig. 3b, significant risks of graft failure were detected in Fanca −/− recipients infused with reduced numbers of gene-corrected Fanca −/− LSK cells. Strikingly, whereas 47% of these animals showed graft failure at 16 weeks postinfusion, no graft failures were observed when these cells were coinfused with MSCs (see analyses of PB, shown in Fig. 4b). Similar results were observed in the hematopoietic organs of recipient mice at the end of follow-up, 16 weeks after gene therapy (Supplementary Fig. S5a–c). Moreover, the comparison of donor engraftments in PB between weeks 4 and 16 posttransplantation showed a significant increase in the group coinfused with corrected Fanca −/− LSK cells and MSCs (p < 0.05), but not in recipients infused only with corrected Fanca −/− LSK cells (p > 0.05), indicating that MSCs facilitated the progressive engraftment of gene-corrected Fanca −/− HSCs in recipient Fanca−/− mice.
VCN analyses determined in euthanized mice (16 weeks postinfusion) showed that, as deduced from analyses of donor engraftment, a significant number of recipients infused only with corrected Fanca −/− LSK cells (8 OF 15) did not show detectable levels of gene marking. However, every recipient that was coinfused with corrected Fanca −/− LSK cells plus MSCs showed the presence of the therapeutic vector in PB cells (Fig. 4c). Once again, conclusions regarding the efficacy of MSCs to improve the engraftment of donor HSCs were reproduced when the VCN/cell was determined in the hematopoietic tissues of gene therapy-treated mice (Supplementary Fig. S5a–c).
Taken together, our results indicate that graft failures associated with the infusion of limited numbers of corrected Fanca −/− HSCs into syngeneic recipients conditioned with a moderate conditioning regimen can be significantly overcome by the coinfusion of these cells with MSCs.
Discussion
Various experimental and clinical studies have shown that MSCs can significantly improve donor hematopoietic engraftment either in xenogeneic or allogeneic transplants, probably due to the immunoregulatory properties of these cells. 6 –8 Using an autologous HSCT model, we have demonstrated that MSC coinfusion also improves the engraftment of donor HSCs in a mouse model of autologous HSCT, an effect that was correlated with the increased homing of donor HSCs in recipients' BM. 17
Because HSC gene therapy constitutes an extraordinarily relevant application of autologous HSCT, we have now explored whether MSCs could also facilitate the engraftment of gene-corrected HSCs in a clinically relevant gene therapy model, such as FA. Although HSC gene therapy is considered a good therapeutic alternative for patients with FA, lessons learned from previous FA gene therapy trials have shown the difficulty in engrafting gene-corrected autologous HSCs in these patients. 18,19 In several HSC gene therapy trials, the engraftment of gene-corrected HSCs has been facilitated by the conditioning of the patient with high doses of genotoxic drugs, 2 something that is particularly harmful in the case of patients with FA because of their defective ability to repair DNA damage. 23,24 Consequently, the development of nongenotoxic conditioning, 30,31 and/or approaches capable of facilitating the homing of gene-corrected cells in minimally or even unconditioned patients, would improve the efficacy and minimize the toxicity of HSC gene therapy in FA, as well as in several other diseases.
In our current studies we have used a FA model based on the knockout of the Fanca gene. 32 As is the case in many other FA models, 32,33 Fanca −/− mice do not reproduce the bone marrow failure (BMF) that takes place in virtually all patients with FA. Therefore, in all our experiments Fanca −/− recipient mice were exposed to a moderate conditioning of 5 Gy, aiming to facilitate low levels of donor HSC engraftment in recipient hematopoietic tissues.
Our first experiments, in which low numbers of wild-type HSCs were transplanted into wild-type and Fanca −/− recipients (Fig. 1), clearly showed higher incidences of graft failure in Fanca −/− recipients. This observation implied differences in the frequency of wild-type repopulating cells depending on their infusion into wild-type or FA recipients (Supplementary Table S1). In fact, the frequency of repopulating cells determined in FA recipients was 3-fold lower as compared with frequencies determined in wild-type recipients. This observation is consistent with previous studies showing that loss of FA genes impairs the ability of stromal cells to support the engraftment of syngeneic HSCs. 34
Interestingly, despite the defective ability of Fanca −/− recipients to facilitate the engraftment of wild-type HSCs, progressive increases in the level of donor engraftment were observed in Fanca −/−, but not in wild-type, recipients. These results demonstrate that, although the engraftment of healthy HSCs is impaired in Fanca −/− recipients, a concomitant proliferation advantage of engrafted wild-type HSCs takes place in these animals, mimicking the situation described in mosaic patients with FA. 35
Strikingly, our data in Fig. 2 showed that when wild-type HSCs were coinfused with MSCs no graft failures were generated in any transplanted Fanca −/− recipient. In addition, a similar proliferation advantage of donor wild-type HSCs was noted in Fanca −/− recipients, regardless of the coinfusion of MSCs, showing the evident beneficial effect of MSCs when these cells are coinfused with limited numbers of wild-type HSCs in a mouse model of FA.
In our subsequent experiments we aimed at corroborating our observations in a clinically relevant HSC gene therapy model in which gene-corrected Fanca −/− HSCs were infused into Fanca −/− recipients. The results presented in Fig. 3 are consistent with our previous studies showing the long-term repopulating properties of PGK-FANCA-wPRE* LV-transduced Fanca −/− HSCs. 36 In our current study we demonstrate that numbers of either uncorrected or corrected Fanca −/− LSK cells required to repopulate the hematopoiesis of sublethally irradiated Fanca −/− recipients are significantly higher as compared with wild-type LSK numbers. In addition, our current studies show that no proliferation advantage of corrected Fanca −/− cells takes place when these cells are transplanted into Fanca −/− recipients (Fig. 3b), indicating that at least in the Fanca −/− mouse model, and the repopulating ability of gene-corrected grafts, is lower as compared with their wild-type counterparts. Although we cannot rule out the possibility that the ectopic expression of the human FANCA might induce an immune response in 5 Gy-irradiated Fanca −/− recipients, one should also consider that gene correction is not able to restore the repopulating properties characteristic of healthy HSCs.
Our data in Fig. 4 showing that coinfusion of MSCs with gene-corrected Fanca −/− HSCs prevented graft failures in Fanca −/− recipients is evidence of the efficacy of MSCs in this model of FA gene therapy. Interestingly, the progressive increase in donor engraftment in recipients coinfused with MSCs and corrected Fanca −/− LSK cells indicates that MSCs not only prevented graft failures, but also facilitated the development of a proliferation advantage of corrected Fanca −/− HSCs in FA recipients. This observation was confirmed by VCN analyses in the hematopoietic tissues of FA recipient mice, demonstrating that MSCs improved the repopulation of recipient mice with gene-corrected Fanca −/− HSCs.
In previous studies using an autologous HSCT mouse model we proposed that MSCs enhanced donor reconstitution by facilitating the homing of infused HSCs into the BM niche. 17 Whether the same mechanism is also applied to our current HSC gene therapy model would require confirmation, because in this model infused wild-type MSCs might provide restored hematopoietic signals to the BM of FA recipients. In addition, the known immunosuppressive properties of MSCs 6 –8 could protect against cytotoxic cytokines expressed in FA recipients, 37 –39 or against the recipient immune system rejecting gene-corrected Fanca −/− HSCs ectopically expressing hFANCA.
The results presented in this study demonstrate for the first time the efficacy of MSCs to prevent graft failures in a clinically relevant model of HSC gene therapy. Similar applications could be considered for the gene therapy of other monogenic diseases associated with risks of graft failure due to low conditioning regimens or infusion of reduced numbers of gene-corrected HSCs.
Footnotes
Acknowledgments
The authors thank Edilia Almeida and Miguel Angel Martín for assistance in the animal care unit, and Omaira Alberquilla for technical support in flow cytometry. The authors are also grateful to Dr. Paula Rio for discussions on Fanconi anemia.
This work was supported by the following grants: European Commission (EUROFANCOLEN; Innovation, ES2012-305421); Ministerio de Economía y Competitividad (SAF2012-39834); Dirección General de Investigación de la Comunidad de Madrid (CellCAM; Ref S2010/BMD-2420); and Fondo de Investigaciones Sanitarias, Instituto de Salud Carlos III (RETICS-RD12/0019/0023 and PI13/01954).
Author Disclosure
J. Bueren is a consultant of Rocket Pharma.
None of the authors of this article has been (past 5 years), is (currently), or will be employed by any company or organization that may gain or lose financially from publication of the article. The authors declare that they do not have any other competing interests.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.