
Abstract
The development of inhibitory autoantibodies to the infused clotting factor VIII (FVIII) is a major complication for severe hemophilia A management. Novel therapy options for hemophilia have significantly progressed in the last decade, and a gene therapy cure for hemophilia is becoming a reality. However, mechanistic studies of FVIII autoantibodies (FVIII inhibitors) have lagged behind and remain a challenge for both protein replacement and gene therapy. FVIII inhibitor formation is assumed to be a classical T cell–dependent immune response in which cytokines/chemokines play an important role. The study of cytokine profile changes during FVIII inhibitor development may be helpful to understand the mechanism of inhibitor development and to explore potential novel approaches that will minimize the risk. After FVIII–/– mice were treated with intravenous administration of an adeno-associated virus 8 vector encoding human FVIII, FVIII expression peaked at week 2 (W2), and FVIII inhibitor was thoroughly developed at week 8 (W8). W8 plasma that showed positive FVIII inhibitor, and W2 samples with negative FVIII inhibitor (anti-FVIII[+]), were subjected to multiplex cytokines measurement. W8 and W2 samples were both negative for FVIII inhibitor (anti-FVIII[–]) as the control. In comparison to mice in the anti-FVIII(–) group, mice in the anti-FVIII(+) group exhibited significantly elevated pro-inflammatory cytokines of interleukin (IL)-1, IL-6, IL-12p40, monocyte chemoattractant protein-1, macrophage inflammatory protein (MIP)-1, MIP-2, and tumor necrosis factor alpha (TNF-α), especially at higher titers. The anti-inflammatory cytokine of transforming growth factor beta (TGF-β) was decreased at W2 in both groups. Multivariate analysis of the risk factors for FVIII inhibitor development showed peak FVIII activity at W2. IL-6 and TNF-α at W8 were positively correlated with inhibitor formation, and negatively correlated with the age starting gene therapy. Collectively, the elevated monocyte derived pro-inflammatory cytokines/chemokines, together with the decreased anti-inflammatory cytokine of TGF-β at an early time point, may contribute to the persistent inflammatory environment in favor of an immune response toward FVIII inhibitor development.
Introduction
H
Management of hemophilia has evolved significantly in the past decade, including the half-life extended coagulation factor concentrates, and the ongoing clinical trials with successful results from adeno-associated virus (AAV)-based gene therapy for patients with both hemophilia A and B. Patients with detectible FVIII inhibitors are ineligible for gene therapy. In preclinical studies, approximately 40% of mice developed FVIII inhibitors after the systemic administration of AAV vector encoding human FVIII. The mechanisms leading to FVIII inhibitor development after gene therapy remain poorly understood. This study investigated the cytokine profile shift before FVIII inhibitor development (at week 2) and after inhibitor formation (at week 8) following AAV-mediated gene therapy in the FVIII-deficient mouse model. It was found that a decrease of the immunomodulatory cytokine TGF-β at the early time point, together with increased pro-inflammatory cytokines and chemokines, especially IL-1, IL-6, IL-12p40, monocyte chemotactic protein 1 (MCP-1), macrophage inflammatory protein (MIP)-1 and -2, and TNF-α, may be involved in the development of antibodies against FVIII.
Materials and Methods
FVIII expression constructs and AAV vector
Human FVIII expression cDNA, kindly provided by St. Jude Children's Research Hospital, 12 was driven by the liver-specific transthyretin (TTR) promoter, as described previously, 13 and packaged in AAV8 serotype. The vector was manufactured based on Good Manufacturing Practice criteria using a triple transfection protocol and titrated at the Virus Vector Core Facility at the University of North Carolina at Chapel Hill, as described previously. 14
Mice
FVIII-deficient mice (FVIII−/−) with a targeted deletion of exon 16 of the FVIIII gene (B6;129S-F8tm1Kaz/J) 15 were originally supplied by H.H. Kazazian Jr.'s laboratory (University of Pennsylvania, Philadelphia, PA), and were bred in-house. Only male mice were used in the study, given that the majority of hemophilia patients are males.
At 8–20 weeks of age, the FVIII−/− mice received a single dose injection of AAV8.hFVIII at a dose of 4 × 1012 vector genomes (vg)/kg of body weight via the tail vein at a volume of around 200 μL. The vectors were diluted with sterile normal saline. This strain of mouse is susceptible to FVIII inhibitor development, and the majority of the mice developed FVIII inhibitors after three doses of human FVIII protein infusion. 16 Blood samples post injection were harvested from the retro-orbital plexus into 3.2% sodium citrate under general anesthesia by continuous inhalation of 2.5% isoflurane. After each phlebotomy sample collection, neomycin cream was applied on the cornea to avoid any potential inflammation. All mouse studies were approved by the Institutional Animal Care and Use Committee of the University of North Carolina at Chapel Hill. Plasma samples were stored at −80°C before analysis.
Measurement of FVIII activity and inhibitory antibodies
Human FVIII activity was measured based on the one-stage FVIII activity assay (FVIII-specific aPTT). Recombinant human FVIII (ADVATE®; Baxter, Irvine, CA) that had been diluted with pooled FVIII-deficient mouse plasma was used to prepare the standard curve. The titer of anti-human FVIII inhibitory antibody was measured by the Bethesda assay, as previously described, 13,17 using a Start 4 Coagulation Analyzer (Diagnostica Stago, Asnières, France).
Multiplex cytokine measurement
The week 8 (W8) sample without FVIII inhibitor that also had FVIII activity remained stable in comparison to its corresponding week 2 (W2) sample, and was classified as a negative anti-FVIII(–) control, while the W8 sample with >1 BU/mL of anti-FVIII inhibitor and FVIII activity dropped sharply in comparison to the corresponding W2 sample, and was classified as a positive anti-FVIII(+) control.
Eighteen cytokines were measured from samples of plasma at W2 and W8: TGF-β, IL-1α, IL-1β, IL-2, IL-4, IL-5, IL-6, IL-10, IL-12p40, IL-12p70, IL-17, IFN-γ, MCP-1, MIP-1α, MIP-1β, MIP-2, RANTES, and TNF-α. Cytokines were measured on a Luminex MAGPIX system (Luminex Corporation, Austin, TX), equipped with Luminex xPONENT® software using custom kits (BioRad Laboratories, Hercules, CA). Cytokine levels were expressed in picograms per milliliter (pg/mL). The cytokines from W2 and W8 samples of each individual animal were measured from the same plate to minimize intra-plate variation. Levels below the detection limit of each cytokine were defined as 0 pg/mL.
Statistical analysis
Data are shown as the mean ± standard error of the mean. Analysis by one-way analysis of variance for multiple comparisons was performed using GraphPad Prism v7 (GraphPad Software, Inc., La Jolla, CA) unless further defined. A p-value of <0.05 was considered statistically significant. For the analysis of risk factors associated with FVIII inhibitor development, a multiple linear regression model and logistic regression model were used in SAS v9.4 (SAS Institute, Cary, NC) and performed by a biostatistician (Y.Z.). Besides the multiple cytokines, other parameters, such as FVIII activity at W2 and the age at FVIII vector injection, were also analyzed.
Results
FVIII activity and the peak time point of FVIII inhibitor
FVIII-deficient mice at various ages were randomly chosen to receive AAV8.hFVIII vectors via tail-vein injection. There was no statistical difference for age at the point of entering the study in either the inhibitor-positive (anti-FVIII[+]) or -negative (anti-FVIII[–]) groups. In a pilot study, hFVIII protein expression was measured after AAV8.hFVIII-mediated gene therapy, and a good correlation was found to exist between the FVIII protein and activity measurement (data not shown). Hence, only FVIII activity after gene therapy is reported in this paper. After AAV administration, FVIII expression peaked at W2, consistent with a recent report. 18 At the time of FVIII inhibitor development, FVIII activity dropped significantly, as shown in Fig. 1A and B. The FVIII inhibitors were thoroughly developed at W8, even though some treated mice had been followed for up to 20 weeks (Fig. 1C and D). Although FVIII inhibitors could be generated as early as at W2 in a few of the treated animals, as previously reported, 17 the titer of FVIII inhibitor was usually lower in such treated animals, and substantial FVIII activity was still detectable. To model a more homogeneous scenario of no inhibitor at an early time point, any samples that showed a low titer of FVIII inhibitor (0.4 IU/mL to <1.0 IU/mL) at W2 were excluded from further cytokine analysis.
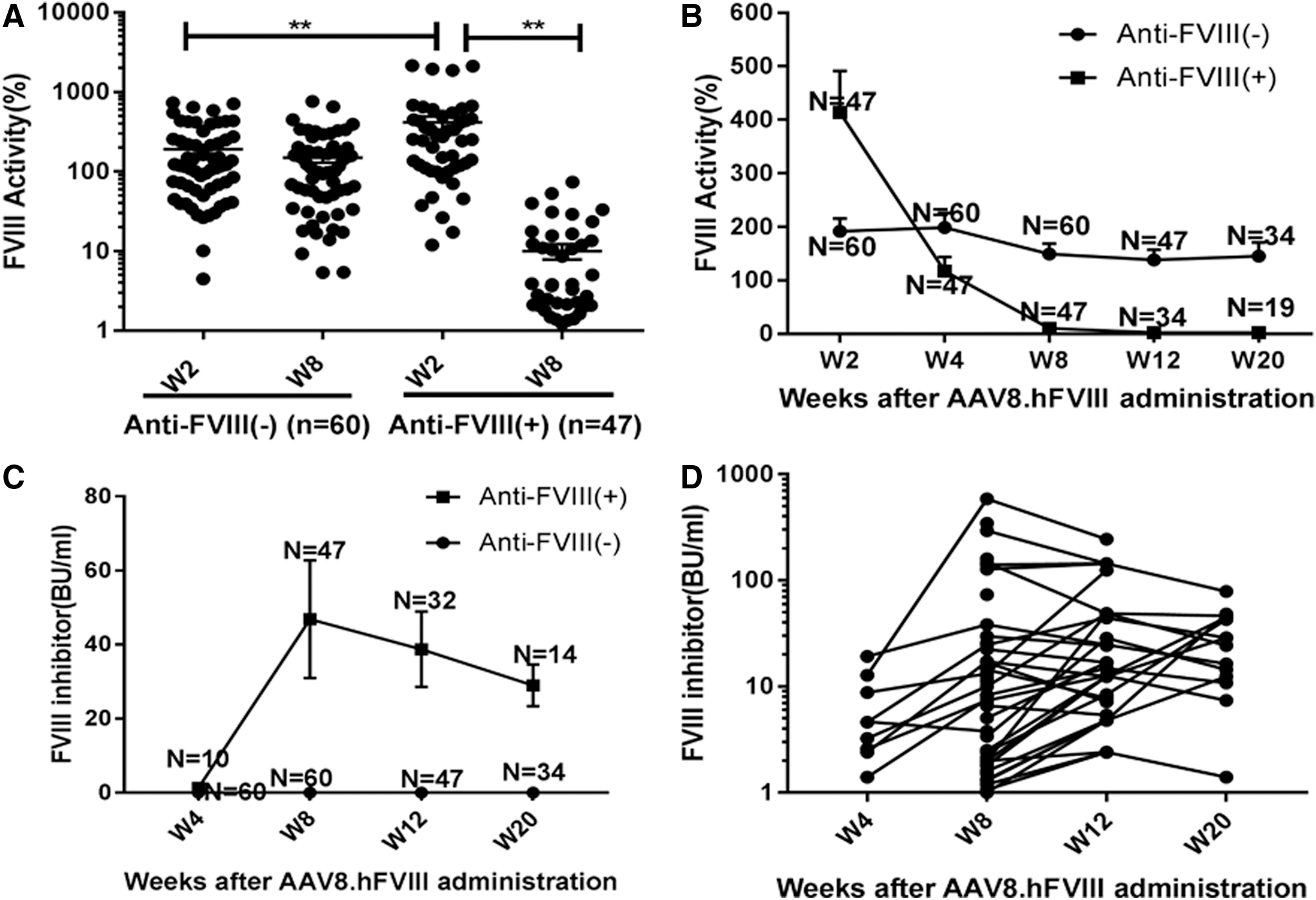
Factor VIII (FVIII) activity and FVIII inhibitor levels after AAV8.hFVIII gene therapy. Adeno-associated virus 8 (AAV8) vectors that express human FVIII were injected intravenously into FVIII/– mice at a dose of 4 × 1012 ppg/kg of body weight. Citrated plasma collected at defined time points post injection were measured for FVIII activities in percentage with recombinant human factor VIII as the standard and anti-FVIII inhibitor in Bethesda units/mL (Bethesda assay). At week 8 (W8), after the determination of inhibitor development, mice were retrospectively divided into two subgroups: anti-FVIII(–) and anti-FVIII(+).
Interestingly, FVIII inhibitor development is negatively related to mouse age at the time of AAV injection. Specifically, when older mice were treated with AAV/FVIII vectors, a relatively lower incidence of the FVIII inhibitors was observed when the relationship between age starting AAV8.hFVIII and the incidence of FVIII inhibitor formation was retrospectively summarized (Supplementary Fig. S1). The tendency was further confirmed by multivariate regression analysis (Table 1). However, the result is not associated with the AAV transduction efficiency between different ages (data not shown).
Multivariate logistic regression analysis for predictors of anti-FVIII inhibitor development (odds ratio)
An odds ratio >1.0 represents that the particular variable increases the chance of FVIII inhibitor development, and <1.0 decreases the chance. For example for the variable of age, while the age of the mouse increases one unit (i.e., 1 week), the odds for developing FVIII inhibitor decrease by 0.675 times, controlling other variables. For the variable IFN-γ W8, however, while the level increases one unit, the odds for FVIII inhibitor increase by 7.823.
FVIII, factor VIII; W2, week 2; IL, interleukin; TNF, tumor necrosis factor; IFN, interferon.
Changes in cytokine/chemokine profile
Overall, there is a wide variation in the cytokine levels, as shown in Fig. 2A and B. In addition, some samples are undetectable for certain cytokines (Supplementary Table S1). The change with the statistical difference (Fig. 2A) includes elevated levels of cytokines such as IL-1α and 1β, IL-6, IL-12p40, MCP-1, MIP-1α, MIP-2, and TNF-α. TGF-β has decreased levels at W2, but these increased significantly at W8 compared to at W2. Cytokines without statistical difference include IFN-γ, IL-2, IL-5, IL-10, IL-12p70, IL-17, MIP-1β, and RANTES (Fig. 2B).
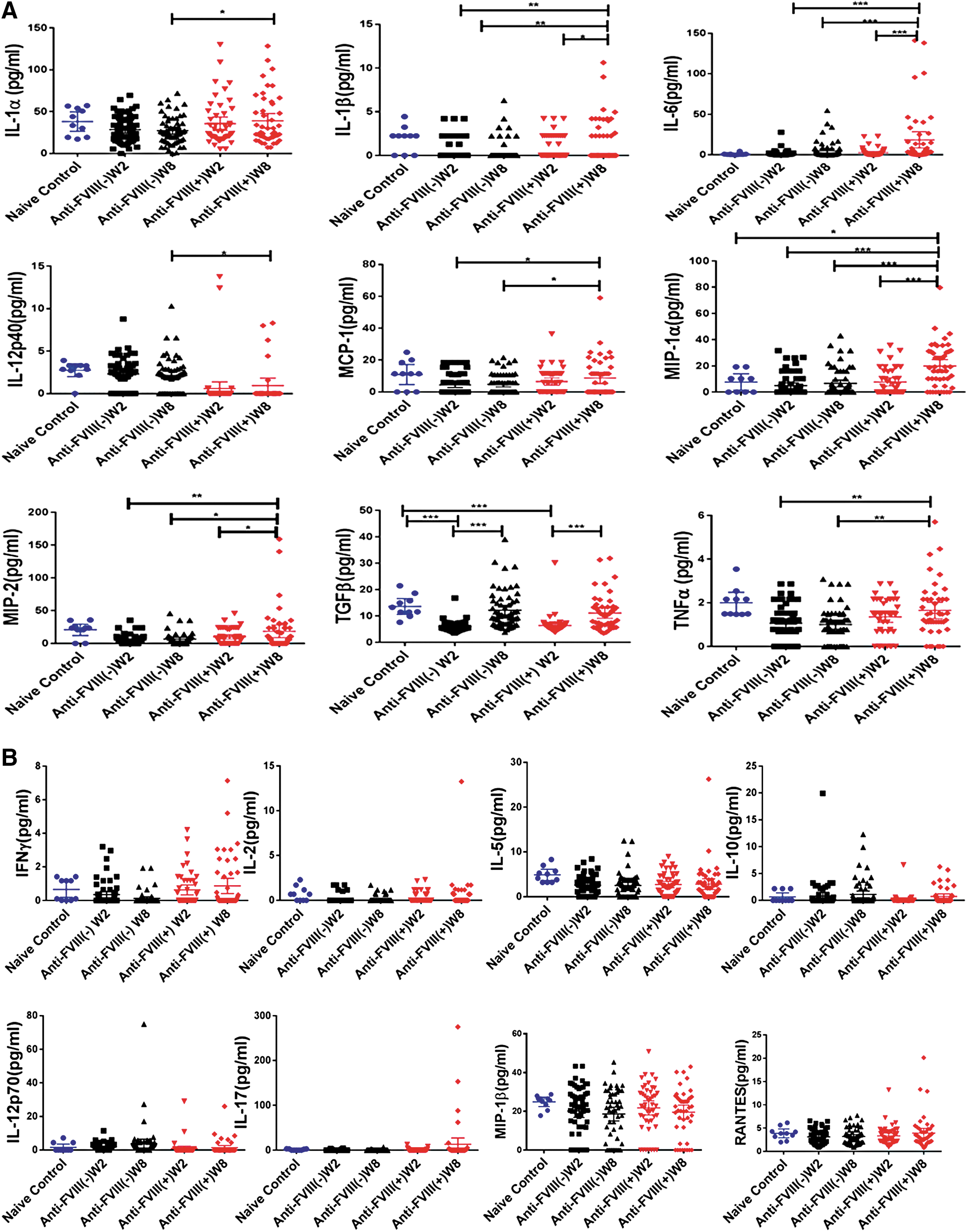
Profile of multiplex cytokines before and after FVIII inhibitor development. Multiple cytokines were measured from samples of plasma at W2 and W8.
A significant elevation of IL-1α was seen in the FVIII inhibitor-positive group when compared to the negative group at W8, and the level at W8 further increased in comparison to W2 for mice that developed a higher titer of FVIII inhibitor (Table 2). For IL-1β, the level at W8 was significantly higher than that at W2 in the inhibitor-positive group. For IL-6, the W8 level in the FVIII positive group was higher than that without the FVIII inhibitor at W8. The level at W8 was also further increased in comparison to W2 in the higher titer mice that developed FVIII inhibitors (Table 2).
Cytokine changes based on the FVIII inhibitor titer
Mice with FVIII inhibitor development were further divided based on “low” titer (<5 IU/mL) and “high” titer (>5 IU/mL), with ranges of FVIII inhibitor titer and average ± median shown in parentheses. Cytokines level are presented as the average ± SEM, and p-values were calculated using a t-test.
ns, not significant; TGF, transforming growth factor; MIP, macrophage inflammatory protein; MCP, monocyte chemotactic protein.
Interestingly, the predominant monocytes/macrophages that derived pro-inflammatory cytokines of MCP-1, MIP-1, and MIP-2 displayed the unanimous pattern that the W8 levels in the FVIII inhibitor-positive group were significantly higher than that of the W8 levels in the FVIII inhibitor-negative group. They were also higher than the W2 levels in the same FVIII inhibitor-positive group. The W8 level also further increased in comparison to the W2 levels in the higher titer mice that developed FVIII inhibitors for MIP-1α and MIP-2 (Table 2).
IL-17 plays a critical role in numerous immune regulatory functions through its involvement in eliciting and mediating pro-inflammatory responses. IL-17 synergizes with other inflammatory cytokines, including TNF-α and IL-1β, and it can further induce the expression of inflammatory cytokines and chemokines, which perpetuate inflammation by recruiting more neutrophils or IL-17-producing T cells. 19 In this study, both the IL-17 levels and numbers of detectable samples appear to be elevated in the inhibitor-positive group at W8 (Fig. 2B and Supplementary Table S1). However, there is no statistical difference between the FVIII inhibitor-positive or -negative groups at any time point due to the large individual variance.
TNF-α has been putatively recognized as a strong pro-inflammatory cytokine, and a previous study showed that anti-TNF-α therapy can minimize the risk of FIX inhibitor formation from “on-demand” FIX protein therapy after multiple hemarthrosis in animal models. 20 W8 levels in the FVIII inhibitor-positive group were significantly higher than the W8 levels in the FVIII inhibitor-negative group and were also higher than the W2 levels in the same FVIII inhibitor-positive group. Furthermore, when the inhibitor-positive animals were further divided based on the titer of the anti-FVIII inhibitor, TNF-α levels in the higher titer group (>5 IU/mL) were greater than that of the lower titer inhibitor group (<5 IU/mL) at both W2 and W8, implicating the active role of TNF-α in FVIII inhibitor formation.
Among the cytokines measured, TGF-β and IL-10 are putatively recognized as anti-inflammatory cytokines. W2 TGF-β levels in both the FVIII inhibitor-negative and -positive groups unanimously decreased compared to that of the naïve control. Nonetheless, W8 TGF-β levels returned to those that were similar to the naïve control, but were significantly higher than the W2 levels in both the FVIII inhibitor-positive and -negative groups. Due to the small numbers of samples with detectible IL-10 and the large individual variance, the changes in IL-10 levels are not statistically significant. However, the detectable samples from the inhibitor-negative (19/60 samples) and -positive (6/47 samples) animals at W2 are statistically significant (Supplementary Table S1; p < 0.05).
Overall, the measured cytokines displayed a clear pattern of a decreased anti-inflammatory cytokine profile in mice with FVIII inhibitor formation, represented by the decreased TGF-β and with elevated pro-inflammatory cytokines/chemokines at the early time point. These changes may contribute to a persistent inflammatory response that favors the development of the FVIII inhibitor after gene therapy.
Risk factor analysis
As it is reported that no direct correlation exists between the peak of FVIII expression and that of FVIII antibody formation, 18 multivariate analysis showed that changes in W2 cytokine activity can be a predictor for FVIII inhibitor formation, in which high expression of FVIII can be a contributing factor for FVIII inhibitor formation. Increased IL-6 and TNF-α at W8 also positively correlated with the FVIII inhibitor formation.
Discussion
The management of hemophilia with novel therapeutics has significantly progressed in the last decade, and a cure for hemophilia by gene therapy is translating from a promise into a reality. 21 Investigation into the mechanism of the FVIII inhibitor development after clotting factor infusion has been limited, and the FVIII inhibitor development after gene therapy has rarely been explored. In this study, the gene delivery of human FVIII with an AAV vector in FVIII–/– mice resulted in approximately 30–40% of mice developing hFVIII inhibitory antibodies, as reported previously. 22 The incidence of FVIII inhibitors negatively correlated to age at the time of AAV vector administration. Pro-inflammatory cytokines IL-1, IL-6, MCP-1, MIP-1, MIP-2, and TNF-α were significantly elevated in FVIII inhibitor-positive mice and furthermore in mice with a higher titer of FVIII inhibitors. While the anti-inflammatory cytokine TGF-β was decreased compared to the naïve control at the early W2 time point, IL-10 was detected less in the FVIII inhibitor-positive animals. Multivariate analysis showed that FVIII activity at W2 and IL-6 and TNF-α at W8 were related to inhibitor formation. However, the limitation of this retrospective study is the lack of immunological cells, especially in the re-stimulation of bulk immune cells (splenocytes) to see if the systemic cytokine changes correlated with FVIII re-stimulated splenocytes.
FVIII inhibitor formation has been assumed to be a classical T cell–dependent immune response. 23 It involves FVIII uptake/peptide binding to MHC-II molecules, antigen presentation to CD4 T cells, recognition by the T cell receptors, 24 and ultimately memory B cell activation and antibody secretion, in which the network of cytokines/chemokines plays an important role. Previous studies have demonstrated that higher levels of secreted immunomodulatory cytokines IL-10 and TGF-β, and lower levels of pro-inflammatory cytokines such as IL-17 and IL-2, contribute to the extended tolerance to rFVIII in animal models. 8,24 Consistent with the findings above in hemophilia A animal models, a large cohort study recently demonstrated that stimulation of the peripheral blood mononuclear cells (PBMCs) from inhibitor-free patients resulted in a significant increase in immunoregulatory cytokine TGF-β levels. On the contrary, the production of pro-inflammatory cytokines (i.e., IL-1, IL-6, and IL-12) was much higher in PBMCs from FVIII inhibitor-positive patients. 25 However, some clinical trials have supported the notion that there is a decreased level of pro-inflammatory cytokines TNF-α, IFN, and IL-2 in patients with FVIII inhibitors, while the anti-inflammatory cytokines IL-4 and IL-10 remained unchanged in comparison to inhibitor-free patients and normal controls. These data indicate that a dominated pattern of anti-inflammatory cytokine profile supports the development of FVIII inhibitors, as reported. 10,11 Additionally, another clinical study showed that the existence of a haplotype defined by high IL-10 synthesis is strongly associated with inhibitor development. 6 In this study, around 40% of hemophilia A mice developed FVIII inhibitors after AAV8.hFVIII administration, and a measurement of their resulting immune factor profiles demonstrated that pro-inflammatory cytokine/chemokines were elevated in mice with FVIII inhibitors. Among the elevated cytokines/chemokines, MCP-1 (also known as chemokine C–C motif ligand 2) has chemotactic activity for monocytes and basophils, and causes the degranulation of basophils and mast cells. MIP-1α (also called chemokine C–C motif ligand 3) functions to recruit and activate various circulating monocyte/macrophages and is associated with the influx, accumulation, and activation of highly destructive cells involved in inflammatory processes. 26 IL-6 stimulates the inflammatory and auto-immune processes in many diseases, including diabetes 27 and rheumatoid arthritis, 28 a disease with elevated circulating autoantibodies.
Another remarkable finding from this study was that TGF-β1 was decreased at W2 in all mice treated with AAV8.hFVIII when compared to naïve untreated animals. TGF-β1 acts as a monocyte/macrophage deactivator and is putatively recognized as an immunomodulatory cytokine to suppress the proliferation and differentiation of T cells and B cells while limiting IL-2, IFN-γ, and TNF production. It has been demonstrated that the critical role of TGF-β1 is in the development of immune tolerance in FIX-deficient mice with AAV vector liver targeting. 29 The observed decrease of TGF-β1 at the early time point may be in favor of driving the production of neutralizing antibodies to the expressed FVIII after gene therapy treatment. An increase in pro-inflammatory cytokines may further “deteriorate” into an environment for FVIII inhibitor development. Although the findings that anti-inflammatory cytokines, such as the undetectable IL-4 and IL-10, is not in accordance with clinical observation, the overall pattern supports that the pro-inflammatory, especially monocyte/macrophage derived cytokines, are critical in FVIII inhibitor development after AAV-mediated FVIII gene therapy. In contrast to the findings that a high incidence of FVIII inhibitor development occurs in mice after AAV.hFVIII liver targeting, higher immune tolerance to other transgenes (such as FIX and beta-Gal) was found in mice after AAV hepatic in vivo gene transfer compared to intramuscular application. 30,31 It has also been reported that different capsids might play a role in the development of transgene-specific B cell responses and ultimately affect the anti-hFVIII antibody formation. 18 The use of AAV8 and AAV9 capsids resulted in less FVIII inhibitor development than in AAVrh10, AAVhu37, and AAVrh64R1. 18 In addition, AAV8 is able to induce immune tolerance much more efficiently compared to AAV2 when the FIX transgene was delivered into the liver in a hemophilia B mouse model. 32 In this study, the AAV8 capsid was also used, so other factors may account for the high incidence of inhibitor development after AAV8.hFVIII delivery, such as protein length or structure. It should be kept in mind that different FVIII products may be associated with different immunogenicity between individuals, 33,34 and that the actual impact of any specific product on inhibitor formation can be varied. 35 For instance, second-generation rFVIII concentrates 36 were associated with more frequent inhibitor development in previously untreated patients with severe hemophilia, 37 although another study reported that no differences regarding the inhibitor development were observed among different FVIII products. 38 It should also be kept in mind that the administration of human FVIII transgene product per se can result in nonspecific immune response and further cytokine changes. This is evidenced by the intravenous administration of recombinant murine FVIII into hemophilia A mice, which did not induce significant anti-FVIII immune response but exhibited a high titer of inhibitory antibodies after hFVIII administration with the same dosing schedule. 39 Nonetheless, no inhibitor formation did not represent the establishment of immune tolerance at the later time. 40,41 Further studies to investigate the impact of cytokine changes on immune tolerance would be more clinically significant.
Cytokines are responsible for immunoglobulin G (IgG) subclass switching. 42 In a unique sibling study with two hemophiliacs discordant for inhibitor incidence, different cytokine profile switching has been accounted and held responsible for inhibitor formation. 43,44 Recent observations support relatively clear IgG subclass switching in hemophilia patients with inhibitor, especially IgG4 possessing the prominent inhibitory function. 45,46 However, it should be kept in mind that IgG1 is the prominent subclass in hemophilia animals with inhibitor, 47 which is not in accordance with the results from human patients. A humanized mouse model with human MHC-II molecules might be more suitable for predicting the immunogenicity of a specific FVIII origin in future studies.
Another interesting finding is that aging is one contributing factor for the risk of inhibitor development. The CANAL study reported that the incidence of inhibitors appears to be associated with age at first treatment, although after adjustment for treatment intensity, this association was less apparent. 48 Currently, AAV-mediated gene therapy, especially in hemophilia gene therapy, is only used in adult patients. The present in vivo finding may warrant further investigation before this approach can be translated into young patients.
In summary, this study shows that the decreased immune-modulatory cytokines and increased pro-inflammatory cytokines create an environment that is in favor of the immune response toward expressed FVIII and the development of antibodies against FVIII after AAV gene therapy. The higher incidence of FVIII inhibitors in younger mice receiving AAV treatment implicates that early intervention to prevent immune activation, such as blocking particular pro-inflammatory cytokines, 49 may help to promote immune tolerance and decrease the risk of anti-FVIII inhibitor development after FVIII gene/protein therapy.
Footnotes
Acknowledgments
We are grateful to the Animal Clinical Chemistry Core at the University of North Carolina at Chapel Hill for cytokine measurement with special assistance from Dr. Hyung-Suk Kim. We thank Charles Askew, Lauriel F. Earley, Nikita E. Hall, and Kelly M. Rigsbee for language editing and critical review of the manuscript. This study is partly supported by a research grant from Asklepios BioPharmaceutical to J.S., but the funder had no role in the study design, data collection and analysis, or preparation of the manuscript.
Author Disclosure
Xiao Xiao is a scientific founder of Asklepios BioPharmaceutical and a shareholder at Asklepios BioPharmaceutical. The other authors have no conflicts of interest to declare.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.