
Abstract
Adenovirus (Ad)-based vectors are efficient gene-transfer vehicles to deliver foreign DNA into living organisms, offering large cargo capacity and low immunogenicity and genotoxicity. As Ad shows low integration rates of their genomes into host chromosomes, vector-derived gene expression decreases due to continuous cell cycling in regenerating tissues and dividing cell populations. To overcome this hurdle, adenoviral delivery can be combined with mechanisms leading to maintenance of therapeutic DNA and long-term effects of the desired treatment. Several hybrid Ad vectors (AdV) exploiting various strategies for long-term treatment have been developed and characterized. This review summarizes recent developments of preclinical approaches using hybrid AdVs utilizing either the Sleeping Beauty transposase system for somatic integration into host chromosomes or designer nucleases, including transcription activator-like effector nucleases and clustered regularly interspaced short palindromic repeats/CRISPR-associated protein-9 nuclease for permanent gene editing. Further options on how to optimize these vectors further are discussed, which may lead to future clinical applications of these versatile gene-therapy tools.
Introduction
B
Due to the episomal state of AdV, the lack of a sufficient nuclear retention mechanism, and the inability to be equally passed on to daughter cells during continuous cell division, the number of AdV genomes per cell and therefore also vector-derived transgene expression are lost over time. To overcome this hurdle and to achieve long-term effects after AdV transduction of dividing cells or regenerating tissues, Ad hybrid vectors can be exploited. Ad hybrid vectors combine efficient AdV-based gene transfer with other viral or non-viral genetic elements, enabling long-term effects for the desired treatment approach.
For gene therapeutic approaches aimed at gene addition, integrases, transposases, recombinases, and DNA replicons were used in concert with AdVs to maintain therapeutic DNA within transduced cells. 9 –13 These hybrid vectors require long-term transgene expression to achieve long-lasting phenotypic effects. Recently developed hybrid vectors combine high transduction efficiencies of AdVs with designer nucleases for gene correction or gene disruption. For these DNA sequence-specific gene-editing approaches, designer nucleases such as zinc finger nucleases (ZFNs), transcription activator-like effector nucleases (TALENs), or RNA guides nucleases of the clustered regularly interspaced short palindromic repeats (CRISPR/Cas9) family were explored. Importantly, for these approaches, maintenance of transgene expression is not required because genome modifications mediated by these enzymes are permanent and inherited to daughter cells upon cell division.
Following the concept of “bringing the best of two worlds together,” the collaboration of one or more non-viral elements have been explored in almost all the existing viral vectors, resulting in promising outcomes for diverse applications. This review focuses on the most recent advances in preclinical developments using Ad hybrid vector systems utilizing either Sleeping Beauty (SB) transposase for somatic transgene integration or designer nuclease-mediated somatic gene editing (TALENs and CRISPR/Cas9 systems) that have the potential for future clinical applications.
Sb Transposase-Based Ad Hybrid Vectors
Ever since its “wakening up” in the 1990s, 14 the SB transposon system has played an essential role in genetics studies, biotechnology, and gene therapeutic approaches. 15 Wild-type SB transposase consists of the SB transposase coding region flanked by IRs. For transposition, SB transposase recognizes the IRs and mediates insertion into TA-dinucleotide sites present in the host genome by a “cut-and-paste” mechanism. In a gene-therapy setting, SB components are split into an artificial transposon containing the therapeutic cargo sequence flanked by the IRs and the transposase provided in trans for mobilization of the transposon and subsequent somatic integration. 16 To overcome the weakness of non-viral delivery for in vivo gene therapeutic approaches, researchers explored viral vectors for efficient delivery of the SB transposase integration machinery. This included AAV-, 17 LV-, Ad-, 18 herpes simplex virus, 19 and baculovirus. 20 This review focuses on using AdVs for delivery of the SB transposase system that were first introduced in 2002. 18 Since the HCAdV vector system was already available at that time, Ad/SB transposase systems are solely based on this AdV type.
When the first adenovirus/SB hybrid vector was constructed, 18 only an early version of SB transposase with relative low efficiency was available. 14,18 Over the past decade, the intrinsic activity of the SB system was enhanced by creating a series of hyperactive SB transposases, and engineering of transposons with optimized 3′ and 5′ IRs. The hyperactive SB versions such as SB11, 14 HSB5, 21 and SB100X 22 showed increasing enzymatic activities in vitro. 22 –24 In particular, the latest version SB100X 22 of SB transposase displays a 100-fold higher transposition activity and increased integration into the host genome compared to the original SB version, resulting in enhanced effects. 22,25
A major obstacle for the Ad/SB transposase hybrid vector system was the inefficient transposition from the recombinant adenovirus genome, which mainly persists as a linear DNA monomer. 1 In fact, there is evidence that circular transposon donors such as plasmids serve as better substrates than linear DNA (e.g., adenovirus genomes) for SB transposase-mediated transposition. 1 To solve this problem, the Flp/FRT recombination system was introduced to release the circular transposon from the linear AdV genome. To prove the efficiency of this hybrid vector model, a LacZ marker gene and human coagulation factor IX as a therapeutic gene were successfully delivered in small animals. 18 To treat large-animal models (cats, dogs, sheep, pigs, and primates), the approach was also translated from mice into a large-animal disease model. In this study, the first hyperactive SB transposase (HSB5) was explored in the context of HCAdV. This hybrid vector mediated efficient somatic integration of a canine coagulation factor IX (cFIX) expression cassette in liver and long-term expression of cFIX in a canine hemophilia B model. 26 The maximum tolerated hybrid vector dose and potential genotoxicity associated with SB-mediated integration was investigated in detail. Integration profile analysis revealed a close to random integration pattern with a slight bias toward genes. Note that TA dinucleotides serving as SB transposase target site were predicted to exist approximately 1 × 108 times in the human genome. 17,27 This safety profile and the fact that SB transposase was already explored in the clinic using non-viral delivery 28 also provides hope that the Ad/SB transposase hybrid vector system can be moved forward toward clinical trials in the future.
While the hemophilia model took advantage of the liver-targeting capacity of human adenovirus type 5 (Ad5), the accumulated knowledge from the adenovirus research opened up new directions for the adenovirus/SB system. By identifying CD46, which is expressed on all human hematopoietic stem and progenitor cells (HSPGs) as a high-affinity major receptor for a number of human adenoviruses, 29 researchers have applied a series of serotype-switch approaches. This included adenovirus type 35 (Ad35)-based vectors, or AdVs containing the Ad35 fiber or fiber knob (Ad5/35), thereby efficiently transducing human and nonhuman primate HSPCs in vitro. 30,32 Based on this knowledge, it was recently shown that an even further advanced AdV/SB transposase hybrid vector system utilizing the hyperactive SB transposase SB100X can be also applied in vivo for stable transduction of HSPCs. 33 In non-viral approaches, this hyperactive SB transposase was demonstrated to result in long-term transgene integration in HSPCs. 22 Furthermore, it was shown that a hybrid vector utilizing SB100X for transposition shows improved integration efficiencies compared to the Ad/SB transposase hybrid vector, taking advantage of HSB5. 25 Building up on this data set, a recent study provided evidence for efficient in vivo targeting of HSPCs in a CD46+ humanized mouse model using an HCAdV/SB100X hybrid vector based on the Ad5/35. 33 Here, the authors mobilized HSPCs cells from bone marrow using a combination of Plerixafor (AMD3100) and granulocyte colony-stimulating factor (G-CSF). 34 This specific combination of two drugs leads to greatest quantities of HSPCs, finally overcoming the physical barriers formed by the bone-marrow stroma. Taken together, Ad/SB transposase hybrid vectors used in concert with AdV5/35 HCAdV delivery enables gene therapy targeting HSPCs. A schematic overview of the mode of action of the AdV/SB transposase hybrid vector system is shown in Fig. 1A.
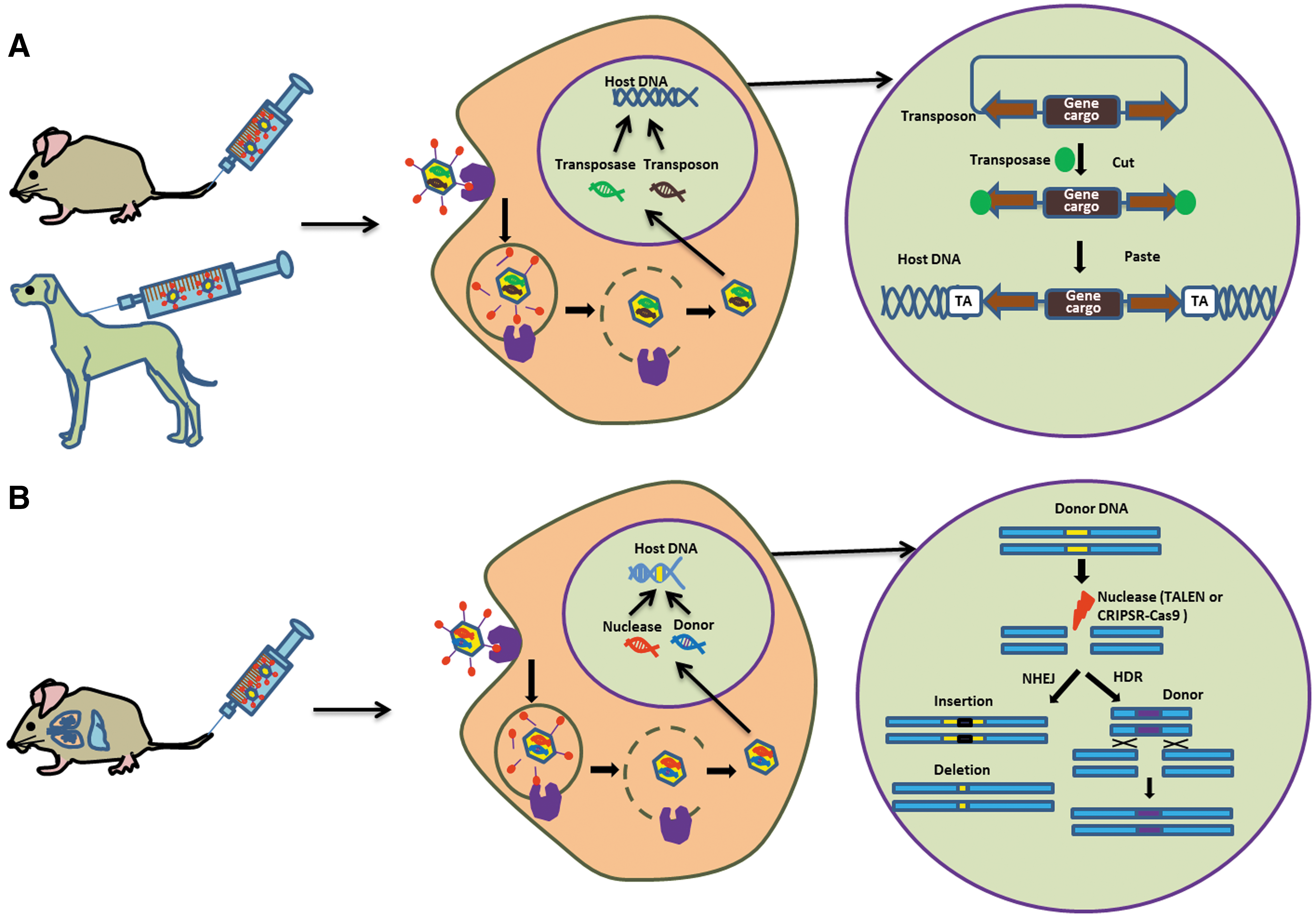
Preclinical in vivo applications of adenovirus (Ad) hybrid vectors either utilizing Sleeping Beauty (SB) transposase or designer nucleases for permanent genome modification and their mode of action. Adenoviral vectors (AdVs) infect host cells by binding of fibers to receptors on the cell surface and then enter cells via endocyctosis. Acidification of endosome releases the partially uncoated virion into the cytoplasm, which then migrates to the nucleus. After entry into the nucleus via nuclear pores, viral DNA including gene modification tools are released into the nucleus.
Ad Hybrid Vectors for Delivery of Designer Nucleases
In various studies, Ad hybrid vectors were used to deliver designer nucleases to achieve permanent treatment effects. TALENs and CRISPR/Cas9 were especially applied for targeted genome engineering. The strategy is usually based on double-stranded DNA break (DSB) induction at defined chromosomal target sequences, activating DNA repair via non-homologous end-joining (NHEJ) or homologous recombination (HR), resulting in gene disruption, gene repair, or gene addition.
TALENs contain a DNA-binding domain of transcription activator-like effectors (TALEs) originating from the plant pathogen Xanthomonas. 35,36 TALEs have a modular structure containing four different repeats, consisting of about 34 conserved amino acids, wherein two variable residues at positions 12 and 13 mediate the interaction with one of the four nucleotides of the DNA target site. To construct a customized TALE-DNA binding domain, the different TALE repeats can be arranged sequentially to obtain sequence-specific binding properties. To use this molecule for genome editing, FokI nuclease domain is fused at the C-terminus of the DNA binding domain. 37,38 Since FokI nuclease is only active after dimerization, two TALEN molecules need to bind to complementary DNA sequences at a distance of about 20 bp to achieve efficient DSB induction.
The CRISPR/Cas9 system is a RNA-guided nuclease (RGN) comprised of a ribonucleoprotein complex originally identified as a part of the bacterial innate immune system. 39 The Cas9 nuclease enzyme incorporates a chimeric guide RNA (gRNA). The gRNA recruits Cas9 to its DNA target site that is complementary to 18–20 distal nucleotides at the 5′ end of the gRNA. 40 The target sequence needs to be followed by a protospacer adjacent motif (PAM) consisting of three nucleotides. The PAM sequence can be different, depending on the bacterial species from which the Cas9 enzyme is derived. Upon target site binding, Cas9 generates a DSB. By simply modifying the 20 distal 5′ nucleotides of the gRNA expression unit, the CRISPR/Cas9 system can be customized to bind and cut a desired DNA sequence specifically followed by a PAM. 41 Because of its simplicity, CRISPR/Cas9 became the most widely used designer nuclease for targeted gene-modification approaches in eukaryotic cells.
Due to their large cargo capacity, AdVs are well suited to construct hybrid vectors equipped with relatively large genes encoding designer nucleases. First approaches for adenoviral delivery of TALENs employed two early-generation AdVs, each encoding a single TALEN gene of an AAVS1 specific TALEN pair. Interestingly, highly repetitive TALEN genes were relatively stable if contained in AdVs, whereas the same genes underwent rearrangements within the TALE repeat region when packaged in LV vectors. 42 In line with this latter study, a detailed protocol on how to construct and characterize AdVs for the delivery of AAVS1-specific TALENs was presented, with the objective of providing tools for integration of transgenes into a safe harbor in the host genome by HR. 43 By using these vectors together with an Ad donor vector delivering a DNA donor cassette for HR-mediated integration into the AAVS1 locus, efficient target site modification was demonstrated. In contrast to LV-mediated nuclease expression used together with LV- or non-viral based HR donor delivery, adenoviral nuclease expression in concert with adenoviral HR donor delivery resulted in most precise genome modification. 44 At the same time, CRISPR/Cas9-based gene editing of the AAVS1 locus was also investigated using AdVs. Skeletal muscle progenitor cells and bone marrow–derived mesenchymal stromal cells were successfully co-transduced with one AdV containing the Cas9 gene and a second AdV containing the corresponding AAVS1 gRNA. In a direct comparison using either AdV co-delivery of the AAVS1-specific TALEN pair or AdV co- delivery of the CRISPR/Cas9 system, similar efficiencies of both systems were observed. 45 As CRISPR/Cas9 provides similar genome-editing efficiencies and is more convenient to work with, most recent studies related to Ad hybrid vectors exploited this nuclease system.
For instance, one study that explored an AdV encoding the complete machinery of a Mcc1-specific CRISPR/Cas9 system showed efficient gene knockout in 3T3 fibroblasts. Mcc1-specific CRISPR/Cas9 gene-editing efficiencies following adenoviral delivery were compared to LV-mediated co-delivery, where one LV contained the gRNA and the other Cas9. For both systems, equal gene-editing efficiencies were observed. 46 Moreover, the study presented a first in vivo application using a single Ad hybrid vector encoding a mutated Cas9 gene (Cas9n) and two gRNA expression units, specific for the CEBPα transcription factor gene involved in liver glucose and lipid metabolism. Cas9n was used with the intention to reduce possible off-target effects, as it is only capable of introducing nicks instead of DNA double-strand breaks (DNA-DSB). After tail-vein injection, efficient CEBPα gene disruption and strong reduction of CEBPα expression was detected in the liver of treated mice, showing that CRISPR/Cas9-mediated gene knockout upon adenoviral delivery can inhibit factors that negatively influence metabolic pathways. 46
Another Ad hybrid vector delivering CRISPR/Cas9 for targeted gene disruption was also used in an antiviral approach to establish human immunodeficiency virus type 1 (HIV-1) resistance in primary CD4+ T cells. Here, a chimeric Ad5/35 vector successfully delivered Cas9 linked to EGFP via an E2A peptide, together with a gRNA specific for the HIV co-receptor CCR5 to patient-derived CD4+ T cells. Upon transduction, efficient CCR5 gene disruption and decrease in CCR5 expression on the cell surface was observed. Most importantly, CD4+ T cells that were transduced by the CCR5-specific CRISPR/Cas9 AdV5/35 hybrid vector showed decreased susceptibility to HIV-1 infection. 47
Furthermore, Ad hybrid vector systems were exploited in antitumor studies. One study analyzed a hybrid vector containing a EGFR mutation-specific gRNA and Cas9 to arm an oncolytic AdV. Here, vector-mediated disruption of the oncogenic EGFR mutation site resulted in enhanced cancer cell killing and tumor size reduction in a xenograft mouse model of human lung cancer. 48
Besides using hybrid vectors as oncolytic tools, CRISPR/Cas9 expressing AdVs were successfully applied to engineer oncogenic chromosomal rearrangements in mice to generate a model for the preclinical investigation of adenocarcinoma formation in the lung. After co-transduction with a Cas9 containing AdV and a second AdV delivering two gRNAs specific for the Eml4 and Alk genes, respectively, inversions of the region between these loci were induced at high frequency, resembling Eml4–Alk dysregulation in human adenocarcinomas. Moreover, these chromosomal rearrangements, which induced adenocarcinoma formation in mice, showed histopathological features identical to those observed in human adenocarcinomas. 49
Another example of CRISPR/Cas-AdV application for disease modeling was shown for the nonalcoholic steatohepatitis (NASH). An AdV vector delivering CRISPR/Cas9 targeting Pten, which is a negative regulator of the PI3K-AKT pathway, mediated efficient Pten gene editing in CRISPR/Cas9 AdV-infused mice. Treated mice developed hepatomegaly and features of NASH, providing a model to study human liver diseases. 50
In contrast to gene-disruption approaches, CRISPR/Cas9-containing Ad hybrid vectors were also exploited in preclinical approaches to correct mutations underlying inherited diseases such as Duchenne muscular dystrophy (DMD). 51 –53 AdVs containing DMD-specific CRISPR/Cas9 alone or together with DMD-specific TALENs were applied for endogenous DMD repair through NHEJ. This repair mechanism incorporates insertions and deletions (in/dels) at out-of-frame sequences for reading frame resetting and splice acceptor knockout for DNA-level exon skipping. Furthermore, CRISPR/Cas9 or CRISPR/Cas9-TALEN multiplexing can be used for targeted exon removal. Here, genome editing after adenoviral delivery of designer nuclease genes were proven to be versatile and robust for repairing DMD mutations. Up to 37% of corrected DMD templates of patient-derived muscle progenitor cells showed efficient gene correction. Note that AdV-mediated transient designer nuclease expression lead to permanent and regulated restoration of dystrophin synthesis from corrected DMD alleles in dystrophic muscle cells from DMD patients. 52 Another study demonstrated that in vivo intramuscular injection with two AdVs containing Cas9 and two DMD-specific gRNAs binding to intronic sequences flanking the mutated exon 23, respectively, mediated efficient exon 23 excision in the mdx DMD mouse model. Furthermore, CRISPR/Cas9 AdV treatment restored dystrophin expression and dystrophin–glycoprotein complex formation at the sarcolemma of skeletal muscles, providing evidence for permanent gene correction of DMD after in vivo adenoviral CRISPR/Cas9 delivery. 53
CRISPR/Cas9-containing Ad hybrid vectors were also exploited in preclinical approaches to correct mutations underlying inherited diseases such as sickle-cell disease (SCD). 54 In these studies. CRISPR/Cas9-mediated enhanced correction of patient-derived induced pluripotent stem cells (iPSCs) in the context of SCD was detected. The sickle-cell mutation was corrected by delivering CRISPR/Cas9 utilizing an AdV followed by nucleoporation with a 70 nt single-stranded oligodeoxynucleotide correction template. Correction efficiencies of up to 67.9% were obtained, demonstrating that adenoviral delivery of CRISPR/Cas9 provides a rapid and efficient method for generation of gene-corrected and patient-specific iPSCs for therapeutic applications. 54 An outline of AdVs used for delivery of designer nucleases and their mode of action is displayed in Fig. 1B.
Perspectives of Ad Hybrid Vector Systems
AdVs are widely used gene transfer vehicles offering a number of advantages. Due to their large cargo capacity, AdVs are well suited to construct hybrid vectors that not only transport a gene of interest for gene addition approaches, but can also be equipped with gene modification or integration machineries. The exploitation of the SB transposase system was the first significant achievement allowing long-lasting treatment using Ad hybrid vectors in small- and large-animal models. Target cells to deliver hyperactive SB transposase systems in vivo were hepatocytes and HSPCs, which were mobilized from the bone marrow. In the future, this may allow translation toward clinical applications for the treatment of other liver-based genetic diseases besides hemophilia B (e.g., hemophilia A, alpha-1-antitrypsin deficiency, and Wilson's disease) or genetic diseases that can be treated by stable transduction of HSPCs such as severe combined immune deficiencies, adrenoleukodystrophy, and Wiskott–Aldrich syndrome.
HCAdV-based hybrid vectors in particular offer the outstanding possibility of transporting large DNA fragments of foreign DNA of up to 35 kb, allowing a flexible setup of contained DNA fragments. This is in sharp contrast to AAV vectors, which provide a relatively small packaging capacity of up to 4.8 kb. With respect to the Ad/SB transposase hybrid vector system, for instance regulatory DNA sequences such as inducible or tissue-specific promoters can be added. Considering the construction a one-vector-system comprising all components (SB transposase, Flp recombinase, and transposon) for SB transposase-mediated transposition from AdV, only HCAdV may be considered as the vector of choice because only this vector system would offer the required packaging capacity. However, it needs to be taken into account that expression of Flp recombinase and SB transposase would lead to destruction of the complete AdV already during vector production in the producer cell line. Thus, the molecular setup of this one-vector system should include inducible or cell type–specific promoters for expression of recombinases. Another issue for the Ad/SB transposase hybrid vector system that remains to be addressed in detail is that of induced immune responses directed against the encoded recombinases (SB transposase and Flp recombinases) and genotoxic side effects due to activity of recombinases in transduced cells.
Furthermore, for delivery of designer nucleases such as TALENS and CRISPR/Cas9, gene-packaging capacities of viral vectors are limiting. In particular, for AAV vectors, two vectors are required for efficient delivery of the TALEN system, and for the CRISPR/Cas9 system, the size of AAV is limiting, especially if one considers gRNA multiplexing. Furthermore, as nucleases that are permanently expressed under the control of constitutive promotors which might cause unwanted side effects such as genotoxicity and immune responses against these foreign proteins, transient expression using inducible or tissue-specific promoters would be desirable. Ad and LV but unlikely AAV vectors may provide the packaging capacity to use these promoters. As shown in direct comparisons of genome-editing efficiencies of identical CRISPR/Cas9 systems delivered using either LV or AdVs, similar gene-editing effects were observed.
The full potential of AdVs for designer nuclease delivery has not been fully explored. Only in one study was the complete CRISPR machinery including gRNA delivered within one single vector. Note that there is no existing study making use of the HCAdV system to deliver all genetic components of TALENs or CRISPR/Cas9 within one single vector. This would ensure that all components of the respective nuclease system would be delivered and expressed in the same target cell. For a two-vector system, co-delivery of two TALEN genes of a respective TALEN pair or Cas9 and the gRNA contained in two separate vectors harbors the risk of hitting different cells, which would reduce the intended therapeutic effect. Therefore, further development is needed to construct the ultimate hybrid vector providing optimal safety and efficiency for future clinical studies. Nevertheless, for some approaches such as those aiming at targeted integration into safe harbor loci via HR following designer nuclease induced DNA-DSB induction, a two-vector-system may remain indispensable.
With respect to gene-editing approaches requiring homology-directed repair (HDR) or HR-based gene insertion, it remains to be investigated which vector for efficient donor DNA delivery is most suitable for the specific approach and which molecular form (linear or circular DNA, or double- or single-stranded DNA) represents the best donor for the HR reaction. Also, the size of the donor DNA and the length of the flanking homology regions for HR remain to be determined. For instance, as the packaging capacity of AdVs allows large HR donor templates to be delivered, the impact of the different length of homology arms flanking a respective insert would be an issue that needs to be evaluated next in the context of AdV-delivered HR donor vectors. In addition, it seems to be appealing to incorporate further mechanisms to improve HR-directed insertion. With respect to AdV, one study found that adenovirus early genes (E1B55K and E4orf6) can increase efficiencies of HDR. Furthermore, it appears plausible to flank HR cassettes with the same recognition sites recognized by the respective designer nuclease that is used to introduce the DNA DSB at the intended chromosomal locus. This could process the HR donor template in a way that would enhance or favor HDR.
Besides genetic components required for efficient genome editing using designer nuclease systems, the viral vector itself could be improved. In particular, immune responses against the incoming adenovirus capsid proteins remain a major challenge that needs to be addressed. Limitations associated with AdV include the stimulation of immune responses, which is well established for the most commonly used vector type based on Ad5. It is known that anti-Ad5 immunity, including pre-existing immunity, exists in up to 90% of analyzed individuals, 55 and induction of adenovirus-specific CD4 and CD8 positive T lymphocytes 56,57 can eliminate the applied vector and transduced cells. 58,59 Moreover, it was shown that even for HCAdV, systemic administration can lead to lethal acute toxicity in baboons when applied at a high dose. 60 To extend the potential of AdV further for efficient delivery of designer nucleases or the SB transposase system and to avoid these unwanted side effects, genetic 61 or chemical 62 modifications of the capsid or other modes of delivery (e.g., pseudo-hydrodynamic delivery 63,64 ) can be explored. Moreover, alternative Ad types could be investigated. For instance, the cloning and reconstitution of a series of different Ad types circulating in the human population was described. 65 In many different cell lines that were infected with these reporter AdV types, Ads were identified that showed higher transduction rates and enhanced reporter gene expression compared to AdV5. These serotypes represent promising candidates for further conversion into gene-delivery vectors that can help to improve Ad hybrid vectors further regarding tissue or cell tropisms as shown for AdV5/35 vectors. 33 Independent of the method used (genetic or chemical Ad capsid modification or the use of alternative Ads), modified AdVs can improve in vivo applications of Ad hybrid vectors.
In summary, AdVs provide a versatile platform to develop tailored vectors that can be specifically designed for the specific mutation causing a genetic disease. In the future, this may lead to novel clinical concepts in personalized medicine.
Footnotes
Acknowledgments
This work was supported by the German Duchenne Foundation Aktion Benny und Co e.V., Witten Herdecke University internal research funding (IFF2017-12) to E.E-S, the SPP1230 provided by the German Research Foundation (DFG) to A.E., and the Chinese Scholarship Council (CSC) to J.G.
Author Disclosure
The authors do not have anything to disclose.