
Abstract
Gene therapy is a promising approach in the treatment of inherited and common complex disorders of the retina. Preclinical and clinical studies have validated the use of adeno-associated viral vectors (AAV) as a safe and efficient delivery vehicle for gene transfer. Retinal pigment epithelium and rods—and to a lesser extent, cone photoreceptors—can be efficiently targeted with AAV. Other retinal cell types however are more challenging targets. The aim of this study was to characterize the transduction profile and efficiency of in silico designed, synthetic Anc80 AAVs for retinal gene transfer. Three Anc80 variants were evaluated for retinal targeting in mice and primates following subretinal delivery. In the murine retina Anc80L65 demonstrated high level of retinal pigment epithelium and photoreceptor targeting with comparable cone photoreceptor affinity compared to other AAVs. Remarkably, Anc80L65 enhanced transduction kinetics with visible expression as early as day 1 and steady state mRNA levels at day 3. Inner retinal tropism of Anc80 variants demonstrated distinct transduction patterns of Müller glia, retinal ganglion cells and inner nuclear layer neurons. Finally, murine findings with Anc80L65 qualitatively translated to the Rhesus macaque in terms of cell targets, levels and onset of expression. Our findings support the use of Anc80L65 for therapeutic subretinal gene delivery.
Introduction
O
AAV is a small, nonpathogenic, nonenveloped, icosahedral, replication-deficient virus that encodes up to 4.7 kb of the transgene cassette of choice that can then be delivered to multiple types of cells and tissues. More than 100 natural isolates of AAVs have been described so far, many of which result in highly distinct levels and patterns of transduction and cell specificity after subretinal injection. 13,20,21 For example, in the adult mouse retina, most serotypes will result in retinal pigment epithelium (RPE) and more limited photoreceptor transduction, a limitation which was described to be overcome by AAV5, 8, and 9. 22 –24 Inner retina cell types have proven a more challenging target however. Several approaches to optimize current AAV vectors seek to improve transduction of inner and outer retinal neurons and glia cells via different routes of administration. These studies have been met with relative success, but none of these efforts has provided a single vector with an overall more ubiquitously improved response. 25 –32
Cone photoreceptor targeting and transduction has also been less documented, especially in a quantitative manner. Despite successful application of AAV-based gene replacement therapy in different animal models of cone or cone–rod dystrophies, 33 –48 some studies have indicated that there might be a discrepancy in cone targeting between serotypes with high photoreceptor tropism in the primate retina. 49 –54 Cone photoreceptors are responsible for color vision and acuity, so it is not surprising that most patients undergoing visual degeneration only report symptoms when cone-mediated vision starts to deteriorate. Therefore, efficient cone transduction is of the utmost importance when designing future therapeutic vectors, not only to directly target cone degeneration and functional loss in primary cone dystrophies, but also for use in neuroprotective treatments for secondary cone loss in rod-specific disorders. The majority of AAV-based ocular clinical trials have used AAV2 so far, and in at least one of these trials, the lack of functional improvement has been attributed to low levels of transgene expression, 6 an effect that could be countered by using a different serotype with higher capacity for photoreceptors targeting and/or transduction.
In an effort to develop antigenically distinct AAVs and to study the structure–function relationship of AAV, we previously reported on an effort that reconstructed viral capsids along the putative evolutionary lineage of AAV using in silico phylogenetic and statistical modeling. 55 Anc80 is the most distal evolutionary node for which we inferred the sequence using ancestral sequence reconstruction (ASR) and is the predicted common ancestor of most known primate AAVs that are considered for gene therapy applications, including serotypes 1–3 and 6–9, but excludes AAV4 and AAV5. The uncertainty provided by the maximum likelihood ASR method was captured in an Anc80 variant library. Members of this library were evaluated for their ability to yield high titer infectious AAV-like particles. The 65th clone from that screen, Anc80L65, underwent extensive characterization and revealed a potent gene therapy vehicle with broad and unique properties in murine and nonhuman primate (NHP) liver as well as muscle and retina targeting in mouse. 55 The aim of this study was therefore to further characterize the retinal tropism of Anc80L65 and two additional Anc80 variants alongside standard AAV serotypes widely used for retinal gene delivery.
Materials and Methods
Ancestral lineage reconstruction, library, and vector selection and production
Ancestral AAV capsid sequences and lineage reconstruction was done using maximum-likelihood methods and is described in full in. 55 Prior in vitro characterization of the functional variants tested in this study are described by Zinn and colleagues 55 Briefly, a preliminary clonal screen of 776 clones from the 2,048-member Anc80 library was performed. These clones were tested in a functional assay for genome containing particle yield and HEK293 transduction. Anc80 vectors with superior performance on the combination of those two vector functional tests were progressed for further testing. For this study, three top-performing vectors were selected for further testing. Vector constructs used in this study were packaged as single-stranded genomes and expressed the enhanced green fluorescent protein (eGFP) driven by a ubiquitous promoter (cytomegalovirus [CMV]) and further regulated by a woodchuck hepatitis virus post-transcriptional response element (WPRE) to enhance transgene expression.
Vector production, purification, and tittering are described in detail in other publications. 55,56 AAV preparations were titrated by TaqMan qPCR amplification (Applied Biosystems 7500, Life Technologies) on DNAse I–resistant vector genomes copies. Primers and probes detecting promoter, transgene, or poly-adenylation signal coding regions of the transgene cassette were evaluated. Yields for the different vectors preparations used were as follows (in genome capsids per mL): AAV8, between 5E12 and 9E12; AAV2, between 1.7E12 and 2E12; AAV9, 7.2E12; Anc80L65, between 2E12 and 7E12; Anc80L27, between 1E12 and 6E12 and Anc80L121, 1E12. The purity of AAV preparations was evaluated by SDS-PAGE gel electrophoresis. For the NHP studies, vector preparations were titrated 3 times and tested for endotoxin levels (only preparations below 10 EU/mL were injected).
Capsid modelling of Anc80 variants
The SWISS-MODEL structure modelling server 57 (Swiss Institute of Bioinformatics) was used to generate pseudoatomic models of Anc80L27, Anc80L65, and Anc80L121 VP3 using AAV8 crystal structure (PDB 2QA0) as a template. Sixtymer PDBs were further reconstructed using the UCSF Chimera package, 58 (Resource for Biocomputing, Visualization and Informatics), and prepared for electrostatic calculations using the PDB2PQR tool 59 with the PARSE forcefield. Protonation states were assigned at pH7 using PROPKA 60 (Jensen Research Group). Electrostatic calculations were run using the Adaptative Poisson–Boltzmann Solver software. 61 Briefly, the linear Poisson–Boltzmann equation was solved on a 400 × 400 × 400 Å grid to a resolution of 1 Å, using the single Debye–Hückel boundary condition, with the following parameters: dielectric constant of solute, 2; dielectric constant of solvent, 78.5; 0.150M monocations/anions with an exclusion radius of 2 Å. 62 Calculated electrostatic potentials were further projected on the solvent accessible surface of viral capsids and visualized between +/− 5 kT/e using the PyMOL Molecular Graphics System, Version 1.6.0.0 (Schrödinger, LLC).
Animals and in vivo vector administration
Mouse
Wild-type C57BL/6J mice (6–8 weeks old) were purchased from Charles River Laboratories and kept at the Schepens Eye Research Institute (SERI) Animal Facility. All animal procedures were performed in accordance with protocols approved by the institutional animal care and use committees at SERI and conformed to the guidelines on the care and use of animals adopted by the Association for Research in Vision and Ophthalmology (Rockville, MD). Animals were anaesthetized with ketamine/xylazine and subretinal injections were performed with 2 μL of AAV vectors for a 2 × 109 total genome capsids injection dose per eye.
Nonhuman primate
Experiments with rhesus monkeys were performed at New England Primate Research Center. All experimental procedures were approved by the Office for Research Subject Protection, Harvard Medical Area Standing Committee on Animals, the Harvard Medical School Institutional Animal Care and Use Committee. Animals were sedated with ketamine or telazol in combination with dexdomitor. No vitrectomy was performed on these animals, but pressure was relieved by performing an anterior chamber tap of roughly the equivalent volume as the injected vector volume (150 μL). Using a 39-gauge polyimide-tipped cannula, 150 μL of AAV was administered subretinally near the superior vascular arcade at a dose of 1 × 1010 total genome capsids. There was no steroid regimen for these animals initially except for a single perioperative subconjunctival dose of dexamethasone, which is short-acting. However, one animal (358-08) received an intravitreal injection in the contralateral eye as part of a separate study (and thought not to affect the results presented here) and was treated with one dose of intravitreal steroids (triamcinolone acetonide, 40 mg/mL) at 3 weeks postsurgery. After recovering from anesthesia, the animals were monitored weekly/fortnightly until the end of the study for general well-being and with eye exams including indirect ophthalmoscopy. At around 8–12 weeks postsurgery monkeys were euthanized and eyes were harvested.
Fundus imaging
Mouse fundus imaging was performed using a Micron III Retinal Imaging Microscope (Pheonix Research Labs, Pleasanton, CA). Animals were anaesthetized with ketamine/xylazine. One percent Tropicamide Ophthalmic Solution (Akorn) was used to dilate pupils and 0.3% Genteal (Novartis) was applied on the corneal surface prior to imaging. Images were taken at the maximum light intensity with a gain setup of 10. Primate fundus imaging was performed on five animals in the week prior to the end of the study (see Supplementary Table S1; Supplementary Data are available online at
Histology
Sample processing
Mouse eyes were collected and fixed in 4% paraformaldehyde (PFA, Sigma-Aldrich) on ice for 1 hour. Corneas and lenses were dissected out and eye cups were fixed for an extra 30 min in 4% PFA. Dissected eye cups were then submerged and frozen in optimal cutting temperature compound (Tissue-Tek O.C.T, Sakura) and kept at −20°C prior to cryosectioning. Nonhuman primate eyes were dissected after euthanasia and an incision was made on the sclera to allow the fixative to enter the eye. Eyes were incubated in 4% PFA for 24 h at 4°C. On the following day, cornea, lens, and iris were dissected and eye cups were then incubated for another 24 h in 4% PFA at 4°C followed by a 24 h incubation in 20% sucrose for 24 h at 4°C. Eye cups were then dissected into five pieces (superior, inferior, temporal, nasal, and central) and each was frozen in tissue freezing medium (TBS) and kept at −20°C prior to cryosectioning. Sections were done at 12 μm for mouse eyes and 15–18 μm for NHP using a Leica CM1950 cryostat. Flat-mounts for mouse retinas was done following the same protocol for fixation as described above with additional steps of dissecting the retina out and incubating it in 20% sucrose for 30 min at room temperature (RT) before proceeding with immunohistochemistry.
Immunohistochemistry
Antibodies used are listed in Supplementary Table S2. The basic protocol for immunohistochemistry for retinal sections was as follows: 1 h incubation with block solution (1% BSA; 0.1% Triton-X; 5% NGS/NDS) at RT followed by overnight incubation with primary antibody at 4°C. On the next day, slides were washed with 1 × phosphate-buffered saline (PBS) and then incubated with Alexa Fluor conjugated secondary antibody at 1:500 dilution for 2 h at RT followed by 1 × PBS washes and incubation with 4′,6-diamidino-2-phenylindole (DAPI) solution for 30 min at RT. Slides were then mounted using fluorescent mounting media (DAKO), allowed to dry, and imaged. For labelling of retinal flat-mounts, dissected retinas were blocked in 1% BSA, 3% Triton-X and 5% NGS for 1 h at RT then incubated for 12–16 h at 4°C. The following day, retinas were washed in 1 × PBS and incubated for 12–16 h at 4°C in a mix of DAPI solution and Alexa Fluor conjugated secondary antibody. Retinas were then washed in 1 × PBS several times and mounted using Prolong Gold antifade reagent (Life Technologies) and allowed to dry at RT prior to imaging. All imaging was done using a Leica TCS-SP5 Upright Confocal Laser-Scanning Microscope. Confocal laser intensity for the green and red channels were consistent across all images (30% and 55% respectively). Gain settings in the green channel varied between panels. All GFP-positive images showed native GFP expression (no antibody used) except for the image shown in Fig. 3E, panel An80L121, where anti-GFP antibody was used.
Cone count protocol
Cone counts were performed on flat-mount of retinas dissected from injected eyes at 4 weeks postinjection (PI). Immunohistochemistry labelling for the cone-specific marker cone arrestin was performed as described above. For each group confocal images were taken from the injected area as a single image (not z-stack) focused at the cone cell body level with settings kept consistent across each channel for all images. The merge image of the blue, green, and red channels (DAPI, GFP, and cone arrestin, respectively) was then used for processing. Image processing and automated cone counts were done using a script programed and run by FiJi ImageJ. A step-by-step diagram of the process is shown in Supplementary Fig. S1.
Qualitative real-time PCR
For comparing relative GFP transcript levels after subretinal delivery using Anc80 variants and contemporary AAV serotypes, eyes were collected from different timepoints after injections (n = 4–6 per group/timepoint) and mRNA was extracted from whole eye cups using the RNeasy kit (Qiagen). Real-time PCR was performed in triplicates using the TaqMan® gene expression master mix (Thermo Fisher Scientific) on an ABI 7500 Real Time PCR System (Applied Biosystems). Relative transcript levels were assessed using the 2-ΔΔCT method with GAPDH as the reference gene (Taqman Assay Mm99999915_g1; catalog No. 4331182; Life Technologies). The following primers were used for GFP detection (5′–3′): agcaaagaccccaaccagaa (forward) and ggcggcggtacagaa (reverse) and probe was from Life Technologies.
Results
In vivo retinal transduction by ancestral AAV variants
Anc80 was previously predicted to be the closest common ancestor of AAV serotypes 1–3 and 6–9 (Fig. 1A). The closest known AAV is rh.10 which diverges in over 8% in terms of sequence. To account for the uncertainty of the maximum likelihood ASR model, a library of Anc80 variants that permutated 11 dimorphic residues at fixed positions was constructed. 55 Based on particle yield and HEK293 in vitro infectivity, three Anc80 capsid variants, Anc80L27, Anc80L65, and Anc80L121, were selected for in vivo evaluation of retinal transduction 55 (data not shown). In Fig. 1B the amino acid present in each of these 11 sites is compared against AAV8 and AAV2 capsid sequences. The three variants differ from each other in only nine positions and have varying degrees of homology to AAV8 and AAV2 at these positions. Structural mapping of the variant sites predicts four to be exposed on the external side of the capsid, which results in a unique electrostatic profile for each variant, as shown in Fig. 1C (iii–v). More specifically, R/K 169 and T/S 206 are located within the VP1-2 domains. Based on the models of the Anc80 variants, R/K 313 and Q/E 413 are located on the luminal surface of the capsids, whereas N/D 612 is predicted to be buried and located at the threefold interface. Finally, T/E 463, A/T 496, N/S 565, and Q/E 579 are expected to be exposed on the external side of the capsid, in the vicinity of the threefold interface, resulting in the unique surface charge profile for each variant.
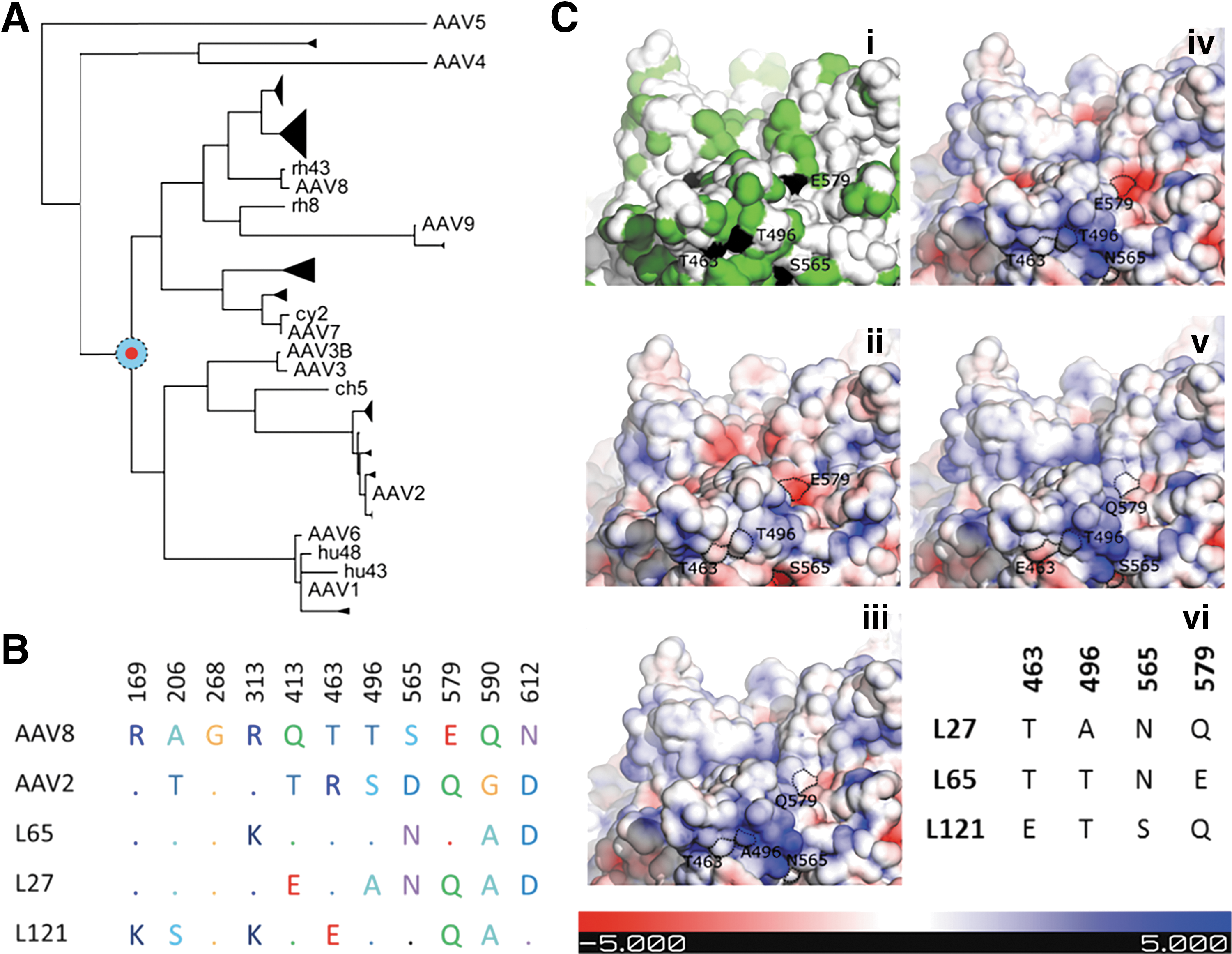
Molecular characterization of Anc80 library variants.
Next, high titer preparation of the selected Anc80 variants, alongside AAV2 and AAV8 controls, were produced in order to interrogate the impact of this structural variation on retinal targeting. Since in the majority of ocular AAV-based gene replacement clinical trials the treatment agent is delivered via the subretinal route, the aim of this study was to evaluate the tropism of the novel Anc80 vectors only after subretinal delivery. All vectors were injected subretinally into adult wild-type male C57Bl/6 mice (6–8 weeks old) at equal doses. Transgene expression was monitored through fundus imaging of the retina at 1, 2, 3, and 4 weeks PI. Representative images for the 4 weeks PI timepoint for all serotypes injected is shown in Fig. 2A. Fundus expression patterns for AAV2 and AAV8 were concordant with the expected performance for these serotypes, in that AAV8 demonstrated higher level of retinal transduction. On average, Anc80L27 and Anc80L65 resulted in strong transgene expression, whereas Anc80L121 generated a less intense expression pattern. Next, eyes were collected for histological analysis at 4 weeks PI (Fig. 2B). All three Anc80 variants were capable of targeting RPE cells and photoreceptors, albeit at differing levels, while AAV2 led to near-exclusive RPE and AAV8 potent RPE and photoreceptor targeting, in line with published results. 22,50 Anc80 variants, and to a lesser extend AAV8, also targeted a substantial number of inner nuclear layer (INL) cells as illustrated in Fig. 2C. Results show Muller glia transduction for all serotypes except AAV2, and for Anc80 variants, a number of additional neuronal retinal cell types were clearly GFP positive.
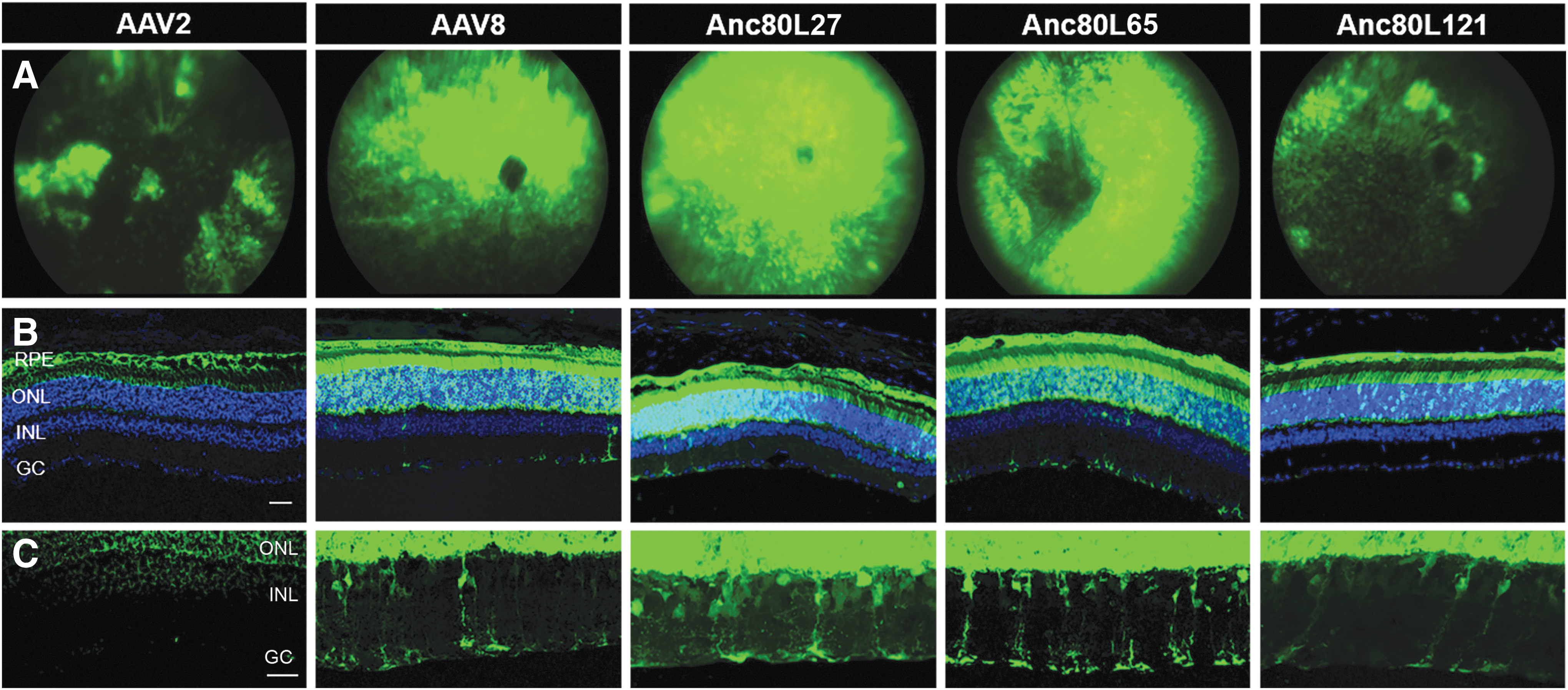
Comparison of transgene expression after subretinal delivery using Anc80 variants, AAV8 and AAV2.
Efficient and unique cell-specific Anc80 INL transduction pattern
To further characterize the INL transduction pattern, cell-specific staining was performed. Retinal sections were subjected to immunohistochemistry using antibodies against specific markers for horizontal, bipolar, amacrine, ganglion and Muller glia cells (Fig. 1; Supplementary Table S2 for antibodies). Different INL targeting profiles were observed with each of the three variants. Variant Anc80L27, and to a much lesser extend Anc80L121, were capable of targeting horizontal cells (Fig. 3A); while all three variants transduced Müller glia cells (Fig. 3D). Strong amacrine cell targeting was seen with the Anc80L27 variant only (Fig. 3C), while none of the three variants was capable of transducing bipolar cells (Fig. 3B). Ganglion cell transduction was seen in two variants but to differing efficiencies; Anc80L27 appears to have the strongest affinity to ganglion cells, while only week and sparse targeting is seen by Anc80L65 and Anc80L121. The ganglion cell transduction pattern for Anc80L27 and Anc80L65 seen in Fig. 3E was also confirmed by GFP positive ganglion cell fibers in the optic nerve of injected eyes (Fig. 3F), where a much stronger GFP expression was seen in the Anc80L27 variant compared to Anc80L65. Variant Anc80L121 did not show any GFP positive ganglion cells on retinal sections but surprisingly weak GFP expression was observed in the optic nerve, whether from ganglion cell or astrocytes fibers remains to be confirmed.
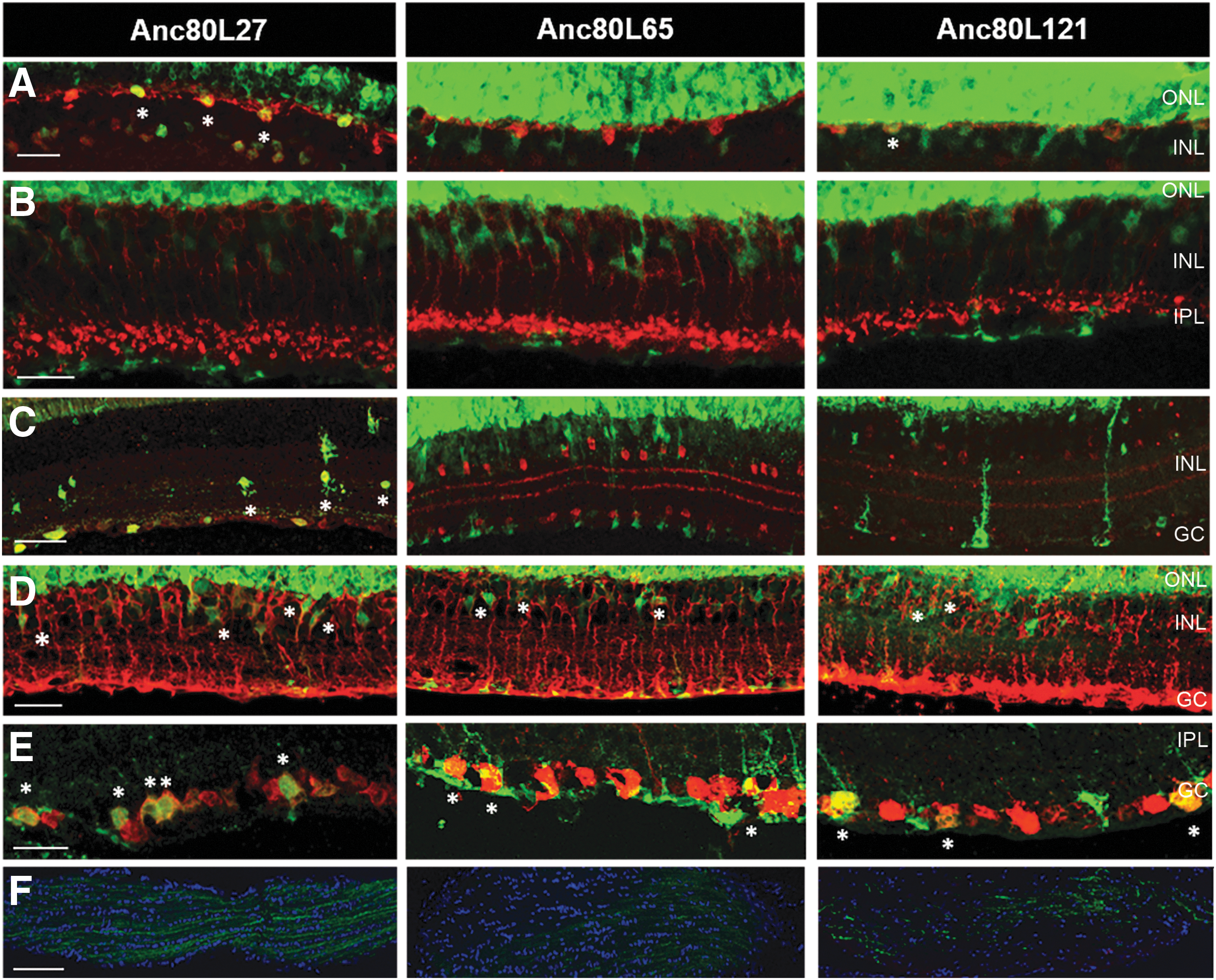
Transduction of INL neurons, glia, and ganglion cells in eyes injected subretinally with Anc80 variants expressing a GFP transgene. The different panels show confocal images of retinal sections after staining for retinal neurons and cell markers. GFP intensity in the ONL was overexposed to reveal the weaker GFP signal observed in the other retinal layers.
Targeting of cone photoreceptors
Given that all three Anc80 variants showed high transduction capacity for outer nuclear layer (ONL) cells, which are dominated in number by rod photoreceptors, we sought to quantify the extent of cone photoreceptor transduction among the AAV serotypes tested. Previous studies have shown that both AAV8 and AAV9 are capable of targeting cone photoreceptors at similar levels in the murine retina 22 but AAV9 is substantially better than AAV8 at transducing cones in the NHP retina. 50 Figure 4A shows the colocalization of GFP positive injected areas of the retina with the mammalian cone-specific marker cone arrestin after subretinal delivery of the three Anc80 variants. Despite colocalization being observed in all three variants, it was also noted that cone transduction was not uniform and that not all cone cells were being targeted. Therefore a more quantitative analysis was performed, comparing the three ancestral variants against conventional serotypes that have high photoreceptor transduction. The selected control serotypes were AAV8, due its high affinity to photoreceptors, and AAV9, which has been shown to have improved cone tropism. 50 This quantitative analysis was done using an automated ImageJ script that was run on single layer confocal images of AAV injected retinal flat-mounts labelled with cone arrestin. Images were taken with the same gain settings, using a 20 × lens plus a 2.5 × optical zoom, and focus was aimed at the cone cell body layer. Due to the irregular nature of retinal flat-mounts, areas imaged that were focused on the outer/inner segment area, cell axon and synapse terminals were eliminated from the analysis (Supplementary Fig. S1). The ImageJ script would then split the red and green channels and convert them to binary images. Processing steps to isolate the cone cells were applied to the red channel, creating a mask of numbered cone cells in the given image. This mask was then applied to the green channel, and the mode intensity for each cone cell was recorded (Supplementary Fig. S1). The same process was applied to an uninjected control image. The highest mode intensity for the uninjected background control (bckg – max mode intensity = 20) was multiplied by three and that value (60) was set as the cutoff for cone cells considered GFP positive.

Comparison of cone transduction efficiency between Anc80 variants and contemporary AAV serotypes.
Figure 4B shows a graph of the percentage of the total number of cones counted for each AAV serotype that were also considered GFP positive. Both Anc80 variants Anc80L65 (67%) and Anc80L121 (53%) had comparable levels of cone targeting compared with AAV9 (48%) and AAV8 (41%) with Anc80L65 showing the highest percentage of transduced cones. Variant Acn80L27 showed the lowest level of cone targeting at around less than half of what was seen for AAV8 at 18%. Figure 4C shows the histogram distribution of all cone cells counted in each group. Despite the almost 20% increase on cone targeting seen when comparing AAV9 to Anc80L65, the natural variation of subretinal injections within the groups meant that Anc80L65 and Anc80L121 were not statistically significant compared to AAV9 and AAV8. The cutoff mark (60) for cones considered GFP positive based on mode intensity is shown on the graph, as is the overall mean mode intensity for all counted cones (Fig. 4C).
Faster onset of expression of Anc80 variants compared with contemporary AAVs
During the course of the above experiments it was noted through fundus imaging that animals injected with two out of the three Anc80 variants (Anc80L27 and Anc80L65) seemed to have a faster onset of GFP expression compared with AAV8 and AAV2 (Fig. 5A). Fundus imaging at the initial PI timepoints showed that already at 1 day PI, considerably higher levels of GFP were being expressed in eyes injected with Anc80L65 and Anc80L27 compared with AAV8. This was maintained at 3 days and 1 week PI (Fig. 5A), even though at 1 week PI, eyes injected with AAV2 and AAV8 were starting to show more sustained and visible levels of GFP expression. This pattern was also confirmed in histological sections of GFP fluorescence, as shown in Fig. 5B. At 3 days PI, strong RPE expression is already present in both eye injected with the Anc80 variants, while very weak expression is seen in AAV8-injected eyes and almost none was observed in AAV2-injected eyes.

Rapid onset of expression of Anc80 variants.
By 1 week PI, all serotypes started to show GFP fluorescence in the RPE, but both Anc80 variants still remained at considerably higher levels compared with AAV8 and AAV2. By 4 weeks PI, GFP expression in AAV8 injected eyes has finally picked up and reached a similar level of expression compared with Anc80L27 and Anc80L65. This is then maintained at a later timepoint of 12 weeks PI. However, fundus or histology images are not reliable methods for GFP quantification; therefore, to quantify this fast onset and higher level of transgene expression observed in eyes injected with these two Anc80 variants, eyes were collected for real-time quantitative PCR (qPCR) analysis at two of the early timepoints (3 days and 1 week PI) and at the last timepoint of 12 weeks PI. Results for the qPCR are shown in Fig. 5C and are normalized for AAV2 expression at the 3 days PI timepoint (ΔΔCt). Interestingly, these results demonstrate that at 3 days PI, AAV8 expression levels are 10-fold higher compared with AAV2, but Anc80L27 and Anc80L65 are an impressive 60- and 148-fold higher, respectively. At 1 week PI, Anc80L27 has increased its expression levels considerably, with a 211-fold increase. Anc80L65 has increased to 188-fold higher levels, while AAV8 and AAV2 remain around 50-fold. At the final timepoint of 12 weeks, where expression levels are believed to have been stabilized, we still observe higher expression of the Anc80 variants which now show a 159- and 155-fold increase for Anc80L27 and Anc80L65, respectively, compared with AAV2 at 3 days PI. AAV8 has stabilized at an 89-fold increase, while AAV2 showed an unexpected decrease from 1 week PI, which was not reflected in the fundus or histology data (17-fold, possibly due to injection quality in this group).
Analogous tropism of Anc80 variant in primate retina
Next we evaluated if the retinal tropism of the Anc80L65 variant observed in the mouse could be replicated in a NHP retina. Rhesus macaque animals were injected subretinally at a dose of 1 × 1010 total genome copies of the same
Furthermore, fundus imaging of a cohort of the eyes are shown in Fig. 6A (i–v), with white light images on the left panel and green fluorescent images on the right. Images 6A (i) and 6A-ii are from AAV9-injected eyes taken at 5 weeks PI (358-08 OD and 411-11 OD, respectively); image 6A-iii is from an AAV5-injected eye at 8 weeks PI (338-04 OD); and images 6A-iv and 6A-v are from Anc80L65-injected eyes taken at 5 weeks PI (411-01 OS) and 8 weeks PI (338-04 OS), respectively. Consistent with previous studies, we observed that in AAV5-injected retinas transgene expression was restricted to the bleb region, while in AAV9- and Anc80L65-injected retinas expression was seen beyond the bleb. 50,54 However, the small number of NHP animals used here does not allow for an in-depth and quantitative characterization of bleb × lateral spread of transgene. The difference in expression levels of the GFP transgene in the NHP retina was then confirmed in retinal sections (Fig. 6B) where comparison of AAV9-, AAV5-, and Anc80L65-injected eyes indicates a much stronger GFP fluorescence in the ONL and RPE after transduction with Anc80L65.
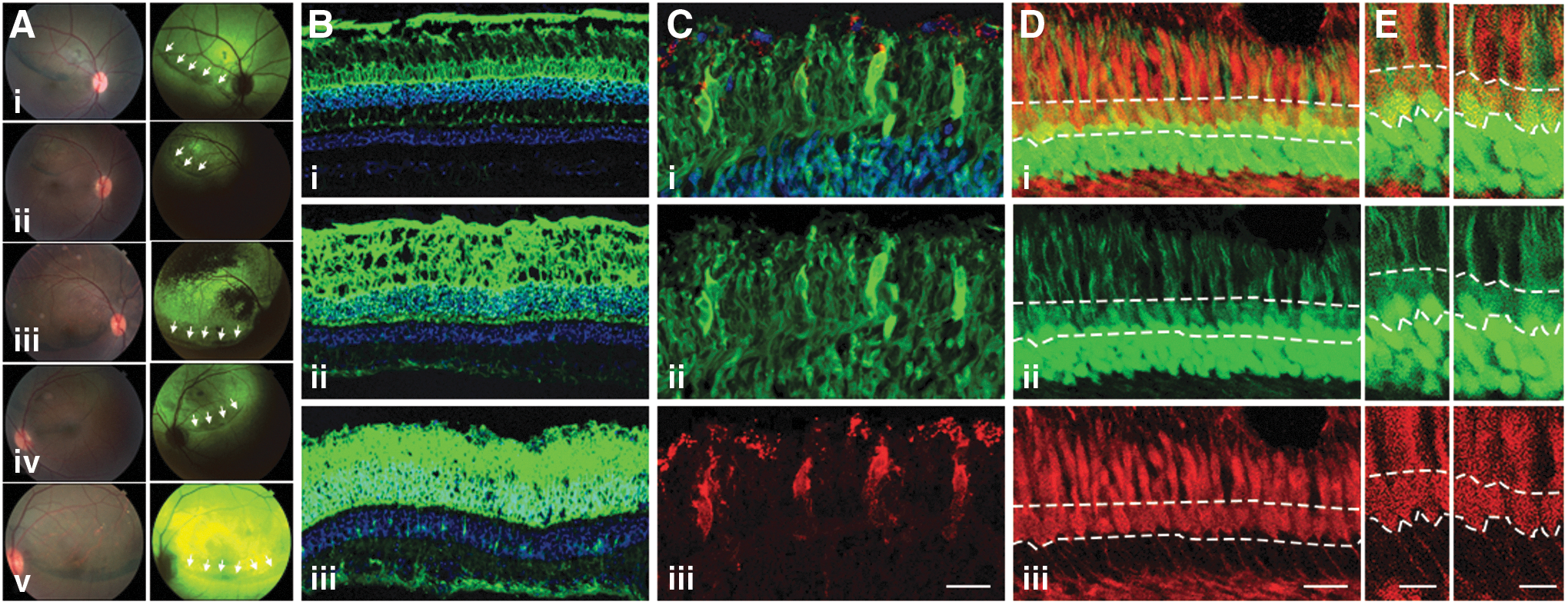
Retinal tropism of Anc80L65 in the nonhuman primate (NHP) retina.
Since the mouse data showed that Anc80L65 was capable of transducing cone photoreceptors, we next wanted to evaluate whether this was also true in the NHP retina. Colocalization of GFP-positive photoreceptors with two different cone markers in eyes injected subretinally with Anc80L65 is shown in Fig. 6C–E. Panel C shows GFP colocalization with peanut-agglutinin (PNA; red) labelling of cone outer segments of eye 431-99 OS, while panels D and E show cone arrestin labelling (red, eye 338-04 OS) colocalizing with GFP in the cone cell bodies. However, despite a number of GFP-positive cones observed, the GFP signal in cones appear on average weaker than in rods and the small number of injected eyes in the NHP study invalidated any quantitative analysis.
Discussion
The recent successful applications of gene therapy for inherited eye disorders have provided valuable information on the safety and efficiency of the currently approved AAV2 vector. 16,63 –65 AAV-based vectors are emerging as attractive vehicles for gene replacement therapies due to its safety and transduction profiles and a range of targeting properties to the various cell type and therapeutic targets in the retina. 13,17,20 However, despite the current extent of information and studies regarding characterization of different AAV serotypes, there are very few vectors currently available that can be used effectively in a wider set of applications. An AAV vector with a broad tropism that could be used in different applications provides a significant advantage on which to develop a platform. For those indications requiring specific targeting, modalities of transcriptional regulation can be then combined with a vector system for which safety and efficacy data is available from other programs.
The distinct tropisms observed in different AAV serotypes is thought to be mediated by the presence of unique elements on the capsid surface generated by sequence variation of the capsid proteins. 66,67 However, modifications on capsid protein sequences have to always take into account structural constraints imposed by the secondary, tertiary, and quaternary interactions of the 60 monomers that form the icosahedral AAV capsid. 66,68 Keeping these constraints in mind, the reconstruction of the AAV lineage and characterization of the furthermost ancestral node recently published by Zinn and colleagues 55 has provided a set of viable libraries of ancestral AAVs from which these structure–function relationships can be investigated. From this effort, one variant—Anc80L65—was described as an infectious and highly stable viral particle with a broad in vivo transduction profile. This preliminary in vivo transduction data indicated a particularly high affinity of Anc80L65 for targeting retinal cells, implying that other variants from within this library might also share similar retinal affinity. Indeed, the results presented here confirm this phenotype for two other variants from the Anc80 library, Anc80L27 and Anc80L121. All three variants—Anc80L27, Anc80L65, and Anc80L121—are capable of efficiently targeting retinal cells via subretinal delivery, and not only did the three variants show extremely high affinity to RPE and photoreceptor cells, but they were also capable of transducing neurons from the INL, Muller glia, and ganglion cells. These results demonstrate a broader potential for Anc80 to target INL neurons and glia via subretinal administration when compared with AAV8, which has only been shown to occasionally transduce ganglion cells and Muller glia. 22,23 Furthermore, Anc80L65 and Anc80L27 variants were shown to have a much faster onset of expression compared with AAV8 and AAV2, with Anc80L65 already showing transgene expression at one day post injection, a property that is relevant experimentally in aggressive retinal degeneration models and possibly in clinical settings where acute intervention is desirable. In addition, our data illustrates that Anc80L65 expression is kept at sustained higher levels compared with AAV8 for up to 12 weeks. Faster onset of expression was also seen in the NHP retina, where Anc80L65 expression onset was, on average, 1 week ahead of AAV9 and 2 weeks ahead of AAV5 (Supplementary Table S1).
Despite the high sequence homology between the variants, it is interesting that a distinct phenotype was observed for each variant. For example, the fast onset reported in variant Anc80L65 was not seen in variant Anc80L121 and was seen to a lesser extent in Anc80L27. Horizontal cell targeting was not observed with Anc80L65 but was present using variants Anc80L27 and Anc80L121. Anc80L27 was the only variant capable of transducing amacrine cells and showed the highest affinity for ganglion cells. These three variants only differ in nine positions in the capsid sequence, which offers a unique opportunity to study the effect of structure–function relationships on viral tropism and helps define how these associations are formed (Fig. 3). Indeed, despite the high homology between the variants, structural mapping of viral capsid proteins shows a diverse electrostatic potential for each variant that could account for these functional differences. Modeling these variants can offer us some insight but comes with limitations. The crystal structure has only been defined for the AAV VP3 capsid protein; therefore, information provided by this approach is based on VP3 only structure and not the native VP1, VP2, and VP3 capsid.
Of the three variants tested here, Anc80L65 has emerged as a strong candidate for retinal gene replacement capable of targeting multiple retinal cell types of therapeutic relevance in mouse and NHP. Of particular interest is the finding that Anc80L65 is at least as good as AAV8 and AAV9 in targeting cone photoreceptors in the murine retina. In this study we were able to, for the first time, fully quantify cone targeting using AAV vectors in the mouse retina. We show that both AAV8 and 9 are capable of targeting only around 40–50% of mouse cone photoreceptors, while Anc80L65 reached 65% of cells in an equal dose comparison. Although not statistically significant, it remains to be seen whether using a cone-specific promoter could drive cone transduction by Anc80L65 to be significantly higher than AAV8 or AAV9. Despite the low target rate observed in the mouse retina, it has been reported that efficiency of cone targeting in the retina of NHPs is substantially increased when using AAV9. Previous studies have shown that, within the injected area, AAV9 at a dose of 1 × 1010 vg is capable of transducing 40% of cones, 50 while AAV5 at a dose of 1 × 1013 vg targets 5–12% of cones. 69 Although the delivery of a therapeutic transgene to primate cones was achieved using AAV5 in only one study, 69 the ability of AAV5 to target primate cones efficiently after subretinal delivery remains unclear, with studies showing no cone targeting at all, 48 weak or locally restricted transgene expression, 53,69 or robust cone targeting across a variety of retinal areas. 54 Other serotypes that have also been shown to efficiently target cone photoreceptors include AAV7m8, 52 AAV8BP2, 52 and AAV2tYF, 70 although proper quantification of cone targeting by these capsids is still needed. Different experimental approaches like choice of promoter, serotype, and postoperative analysis could explain these discrepancies between studies. However, it is most likely that vector dose has the strongest effect on cone targeting, with only concentrations above 1 × 1010 vg capable of efficiently transducing cones. 49,50,52,54
Due to the small number of NHPs used in this study, it was unfortunately not possible to quantify cone transduction in eyes injected with Anc80L65 compared with AAV5 and AAV9. The preliminary results shown here indicate that Anc80L65 is capable of targeting cones in the primate retina, but a more quantifiable and in-depth evaluation of this novel vector is needed to confirm whether it performs in a similar, or indeed better (as shown here for the mouse retina), range as AAV9 and/or AAV5. It would also be interesting to evaluate whether there is differential cone subtype/retinal location targeting by Anc80. Many applications for retinal gene therapy rely on efficient transduction of cone photoreceptors. Since these specialized sensory cells are responsible for high acuity and color vision, their loss is usually felt early and more acutely by patients with inherited retinal degeneration. Even in cases of rod-specific genetic lesions where cone cell dismiss is a secondary effect to a rod cell death, efficient cone targeting would therefore be essential in efforts to protect and preserve vision.
In summary, this study presents an in-depth evaluation of the efficiency and tropism of conventional and designer AAVs for broad retinal gene therapy applications in a small and large animal model. Our work illustrates the impact of minimal changes in capsid composition on aspects relevant to experimental and clinical gene transfer applications. Primarily, we identified a novel AAV with broad potential applications for retinal gene therapy following subretinal injection that can serve as an alternative to the current AAV technologies with in addition phenotypes of fast onset of expression, cone photoreceptor targeting, and access to the majority of other retinal cell types.
Footnotes
Acknowledgments
We would like to thank Dr. Petr Baranov for his help with ImageJ scripting and Dr. Tatjana Jakobs for the kind gift of the ChaT antibody. This work was supported by a donation from Giving/Grousbeck, the Ush2A consortium, the Candyce Henwood Fund, and grants through the Foundation Fighting Blindness, Research to Prevent Blindness, and NIH 5DP1EY023177 (L.H.V.), the Australian Research Council (L.S.C.), and the Harvard Medical School Department of Ophthalmology Age-Related Macular Degeneration Center of Excellence (L.S.C.).
L.S.C. and L.H.V. were responsible for the study outline and experimental design, with L.H.V. responsible for overall supervision of the project. L.S.C. directly conducted and/or supervised all experiments. R.X. and S.S. assisted in murine injections and animal monitoring. E.Z. was responsible for Anc80 variants selection. S.P. conducted the computational analysis and modelling of AAV variants. S.J.W. was responsible for immunohistochemistry and image analysis. A.L. was responsible for NHP sample preparation and analysis. J.I.C., L.A.K., and L.L. conducted the nonhuman primate surgeries and ocular evaluations. L.S.C. and L.H.V. wrote the manuscript.
Author Disclosure
L.H.V. is an inventor on several technologies licensed to pharmaceutical and biotechnology companies including Anc80 technology, which was licensed to Astellas, Vivet Therapeutics, Lonza Houston, and Selecta Biosciences. L.H.V. receives research funding from and consults for Lonza Houston and Selecta Biosciences. L.H.V. is cofounder and equity holder of GenSight Biologics, a retinal gene therapy company. L.H.V. is also a member of the Scientific Advisory Board of GenSight Biologics and NightStarX. For authors L.S.C, R.X., S.J.W., A.L., E.Z., S.P., S.S., J.I.C., L.A.K., and L.L., no competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.