
Abstract
Mutations in the phosphodiesterase 6A gene (PDE6A) result in retinitis pigmentosa (RP) type 43 (RP43) and are responsible for about 4% of autosomal recessive RP. There is currently no treatment for this blinding condition. The aim of this project was to use a large-animal model to test a gene supplementation viral vector designed to be translated for use in a clinical trial for the treatment of RP43. Seven Pde6a–/– puppies were given sub-retinal injections of an adeno-associated viral vector (AAV) serotype 2/8 delivering human PDE6A cDNA under control of a short rhodopsin promoter (AAV8-PDE6A). Three puppies received ∼1 × 1011 vg in one eye and four puppies ∼5 × 1011 vg/per eye, with both eyes being injected in two animals. In vivo outcome measures included vision testing and electroretinography (ERG), as well as fundus and spectral domain–optical coherence tomography imaging. Some puppies were euthanized and their eyes processed for immunohistochemistry. All puppies had improved rod-mediated vision in the treated eye. ERGs showed improved rod-mediated responses in the higher-dose group but in only one of the lower-dose group animals. Receptor+ thickness was preserved and photoreceptor morphology improved in the treated retinal regions in all puppies. Treatment resulted in PDE6A transgene expression, accompanied by much increased levels of Pde6b, in rod outer segments in the injected retinal regions. There were several indications of improved retinal health in the PDE6A-expressing regions, including lack of abnormal cyclic guanosine monophosphate accumulation, appropriate rod opsin localization to the outer segments with a large reduction in mislocalization to other regions of the rod cell, and reduced Müller cell activation. Additionally, cone photoreceptors showed morphological improvement in the treated region, with normal-appearing inner and outer segments. AAV8-PDE6A gene supplementation therapy restored rod vision in Pde6a–/– puppies and preserved retinal morphology. These positive outcomes are an important step toward a human clinical trial to treat PDE6A-RP.
Introduction
T
Recombinant AAV vectors have become the favored delivery tool for retinal gene therapy because of their transduction capacity for different retinal cell types. These vectors have been shown to be safe in clinical trials, only induce a low level of immune response, and provide for long-lasting transgene expression in differentiated cells such as the retinal cells. They rarely integrate into the host DNA, reducing the potential risk of serious effects from insertional disruption of important genetic loci. However, they have the disadvantage of having a limited carrying capacity. 9
Retinitis pigmentosa (RP) is a group of hereditary photoreceptor dystrophies that classically result in a degeneration of rod photoreceptors followed by a slower loss of cone photoreceptors. There is an initial loss of dim-light vision, constriction of visual fields, and, as cones become more severely affected, loss of acuity progressing to legal blindness. RP has an incidence of approximately 1/4,000.
10
It shows genetic heterogeneity with autosomal, recessive, autosomal-dominant, X-linked modes of inheritance (see RetNet, the Retinal Information Network:
Mutations in the rod-specific cyclic guanosine monophosphate phosphodiesterase (PDE6) complex are estimated to cause 36,000 cases of RP worldwide, and yet there is currently no treatment. 12,13 PDE6 is present in rod photoreceptor outer segments where it plays an integral role as part of the phototransduction cascade. PDE6 is a heterotrimer consisting of alpha and beta active subunits combined with two inhibitory gamma subunits. Activation of PDE6 by G-protein action to remove the gamma subunits allows the phosphodiesterase to hydrolyze cyclic guanosine monophosphate (cGMP). A reduction in cGMP leads to closure of cyclic nucleotide-gated channel (CNG) in the outer segment plasma membrane and hyperpolarization of the cell. The stimulation of rods by light results in hyperpolarization of the cell, which is signaled to second-order neurons by a reduction in glutamate release at the rod synaptic terminal. Mutations in both the alpha and beta subunits of PDE6 are linked to RP, 14 –16 and mutations in PDE6A cause RP type 43 (RP43). There are also both small- and large-animal models for both PDE6A- and PDE6B-RP. 17 –20 Gene supplementation therapy in PDE6-mutant animal models has been reported and has proven challenging in some of the more severe models due to the rapid loss of rod photoreceptors. 12,21 –26
Previous gene supplementation therapy in the Pde6a dog model using the canine Pde6a cDNA and a ubiquitous promoter delivered in an AAV serotype 8 with a tyrosine 733 capsid mutation resulted in Pde6a transgene expression in rod photoreceptors, improvement in rod-mediated vision, a small ERG improvement, and photoreceptor preservation in the treated region. However, there were some adverse retinal changes, including retinal rosette formation and patches of degeneration. 22 Expanding on the initial encouraging result, the current study reports improved outcome measures with minimal adverse effects using a vector supplying the human PDE6A cDNA driven by a rod-specific promoter given to a small number of puppies. Improving efficacy of therapy and demonstrating safety in animal models is an important step toward a human clinical trial for the treatment of PDE6A-RP.
Materials and Methods
Animals
All procedures were in compliance with the ARVO Statement for the Use of Animals in Ophthalmic and Visual Research, and approved by the Institutional Animal Care and Use Committee. Dogs were from a Pde6a-mutant colony housed at Michigan State University. The animals used were all homozygous for a previously described frame-shift mutation in Pde6a, 27 and were housed under a 12-hour light:dark cycle. Eight animals were used in the study. Four animals were injected with a lower dose, and four animals were injected with a higher dose (Table 1).
Details of animals, volume and dose injected, and outcome
Animal 16-021 had a congenital cardiac defect and was sickly and was therefore removed from the study.
ERG, electroretinography; VT, vision testing (four-choice exit device); SD-OCT, spectral domain–optical coherence tomography.
rAAV vectors cloning and production
Standard cloning techniques were used for vector assembly. All sequence manipulations were verified by sequencing. The expression cassette consisted of a 0.8 kb rod photoreceptor-specific human rhodopsin (hRHO) promoter, 28 and the full-length (2.58 kb) human PDE6A cDNA, which was amplified from human retinal cDNA with the following primers: forward 5′-AAAGCGGCCGCCACCATGGGCGAGGTGACAGCAG-3′ and reverse 5′-GGAGCGGCCGCTTACTGGATGCAGCAGGAC-3′. The expression cassette also contains a 0.54 kb woodchuck hepatitis virus post-transcriptional regulatory element (WPRE) with mutated WXF-open reading frame and a 0.2 kb bovine growth hormone polyadenylation signal (BGHpA). 29 In a second step, the described expression cassette and the pGL2.0 backbone were assembled to the final pGL2.0-hRHO-hPDE6A-mWPRE-KanR plasmid using a NEBuilder cloning kit (New England BioLabs, Ipswich, MA). The plasmid backbone contains randomized, synthetic DNA fragments (Eurofins Genomics, Ebersberg, Germany), the ITRs from the pSub201 cis-plasmid, 30 a kanamycin resistance gene (KanR), and the pUC18 (Clontech Laboratories, Inc., Mountain View, CA) origin of replication. Production of plasmid DNA and AAV vectors was performed according to standard manufacturing procedure by Aldevron (Fargo, ND) or previously described processes at the Laboratoire de Thérapie Génique, 31 UMR 1089 (Nantes, France), respectively. The pDP8-KanR helper plasmid was used, and the recombinant viral genome was packaged in a wild-type AAV8 serotype capsid.
Sub-retinal injection
Sub-retinal injections in puppies were performed, as previously described. 32 Briefly, under general anesthesia and direct observation through an operating microscope using a Machemer Magnifying Vitrectomy Lens (Ocular Instruments, Bellevue, WA) to visualize the posterior segment, a RetinaJect injector (SurModics, Inc., Irvine, CA) was introduced into the vitreous via a sclerotomy about 2.5 mm posterior to the limbus aiming to pass through pars plana. To limit the pressure increase within the eye post injection, which was noted to be a problem in injections in very young animals, an aqueocentesis was performed of approximately 100 μL prior to the sub-retinal injection. Following the aqueocentesis, the RetinaJect was advanced across the vitreous to target the central retina superior to the optic nerve head. Immediately following completion of the procedure, fundus images were captured using a RetCam II video fundus camera (Clarity Medical Systems, Pleasanton, CA) to record images showing the position/extent of the sub-retinal bleb.
Electroretinography
Electroretinography was performed, as previously described. 33 The procedure was performed under general anesthesia (inhaled isoflurane), globes were positioned in primary gaze using stay sutures of 4-0 silk (Ethicon, Inc., Piscataway, NJ), and the pupils dilated using 1% tropicamide (Mydriacyl; Alcon Laboratories, Honolulu, HI) and 10% phenylephrine hydrochloride (AK-Dilate; Akorn, Inc., Buffalo Grove, IL). Full-field flash ERGs were recorded using ERGJet lenses (Fabrinal SA, La Chaux-de-Fonds, Switzerland) with platinum needle reference and ground electrodes (Grass Instruments, Natus Medical, Inc., Pleasanton, CA) using an Espion 2 electroretinography system with a ColorDome Ganzfeld stimulator (Diagnosys, Inc., Lowell, MA).
A dark-adapted (scotopic) series of increasing flash luminances from below response threshold to a strong stimulus, which in normal dogs elicits a mixed rod–cone response, was recorded followed by a 5 Hz rod flicker. Light-adapted (photopic) single-flash and flicker responses were recorded following light adaptation (10 min at 30 cd/m2).
Vision testing
Assessment of vision was performed using a custom four-choice device previously developed and utilized. 34 –36 Briefly, this consisted of a box with four exit tunnels. For each “run,” the end of only one tunnel was open, this tunnel being randomly selected. The puppies were trained to exit the device through the open tunnel. The first tunnel entered was noted as the “exit choice,” and the time to exit the tunnel was recorded. A series of seven lighting levels were used from bright room light down to a dim light that tests rod-mediated vision in dogs. Thus, a range of scotopic, mesopic, and photopic vision was tested. Each eye was tested separately by fitting an opaque contact lens in the contralateral eye. Fourteen repeat “runs” were performed at each light level for each eye, allowing the mean correct exit choice and mean exit times to be calculated. This is a very objective method of vision testing, allowing statistical analysis.
An obstacle course was also used in two puppies under scotopic conditions to provide video recordings of visual performance. 37 This provided a more subjective assessment of visual performance.
In vivo retinal imaging
Complete eye examinations including slit-lamp and indirect ophthalmoscopy were performed to monitor for any evidence of inflammation or ocular lesions following sub-retinal injection.
22
Wide-angle color fundus images were regularly captured using the RetCam II. Confocal scanning laser ophthalmoscopy (cSLO) and spectral domain–optical coherence tomography (SD-OCT) imaging (Spectralis HRA + OCT; Heidelberg Engineering, Inc., Carlsbad, CA) was performed under general anesthesia, as previously described.
22
Single line and volume scans were performed across the fundus, with particular attention to the interface between the treated and untreated regions (the edge of the injection bleb). Injected (treated) and control eyes were imaged at 2 and 5 months post injection in the low-dose group, and at 2 and 4 months post injection in the high-dose group. The thickness of the Receptor+ (REC+) layers was measured. The REC+ thickness is a representative measure of the total length of the photoreceptor (from the inter-digitation zone, which is the interface of retinal pigment epithelium with the outer segments, to the outer plexiform layer).
38
REC+ thickness was measured using the integrated Heidelberg Eye Explorer (HEYEX) software in five regions throughout the injected area (Supplementary Fig. 1A; Supplementary Data are available online at
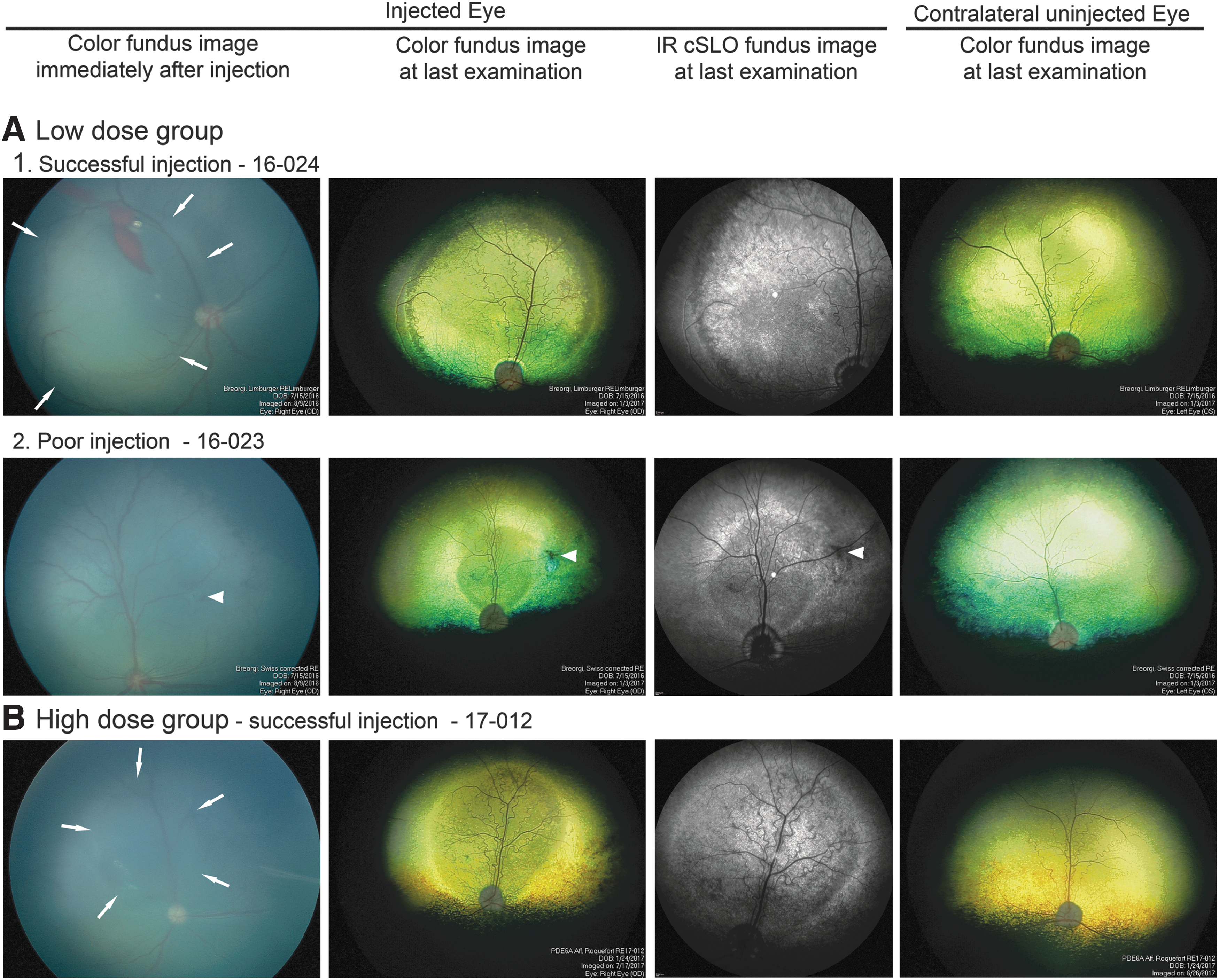
Fundus images of injected and uninjected (control) eyes. The left column shows the image captured immediately after injection. The fundus has a dark blue/gray color because at the injection age, the tapetum has not developed. The second column shows the fundus appearance at the last outcome measure, and the adjacent third column is an infrared (IR) cSLO image of the same eye. The fourth column is the fundus image of the contralateral uninjected eye at last examination.
Retinal layer thickness color “heat maps” were prepared using the HEYEX software, as previously performed. 39
Immunohistochemical labeling
Following euthanasia, eyes were removed and processed for immunohistochemistry (IHC), as previously described. 40,41 Slides were stained with specific antibodies, as summarized in Supplementary Table S1. Cell nuclei were stained with Hoechst33342 or DAPI. Confocal images from the immunolabeled cryosections were collected using a Leica SP8 confocal laser scanning microscope (Wetzlar, Germany) and an Olympus FluoView 1000 (Center Valley, CA).
Statistical analysis
The mean REC+ thickness measurements were compared using a t-test. A mixed-effect model using RStudio was used to analyze the scotopic ERG amplitudes and vision-testing outcomes between treated and control eyes using the equation below
42
:
where β is the parameter vector, X is the independent variable matrix, α i is the animal-level residual, and the ɛ i is the individual observation-level residual. To provide more details, one-tail t-tests were also performed to compare treated versus untreated for each different light stimulus for ERGs and for each different light background for vision testing.
Results
Functional outcome measures
The results are summarized in Table 1.
Low-dose group
Four puppies were given a sub-retinal injection in one eye, aiming to deliver a final dose of 1–2 × 1011 vg of the AAV8-PDE6A vector sub-retinally per eye. The other eye remained as an untreated control. In two animals (16-024 and 16-021), a large sub-retinal injection bleb was achieved (Fig. 1A1). Unfortunately, animal 16-021 had to be removed from the study due to the presence of a congenital cardiac anomaly, and results from this animal were not included in the analysis. In one animal (16-023), a bleb formed but was found to have flattened by the time the post-injection fundus images were obtained (Fig. 1A2). In the final animal (16-025), only a small bleb developed at the first injection attempt. Therefore, a second injection was performed at a separate region in the same eye, but this was seen to have flattened by the time the post-procedural fundus image was captured.
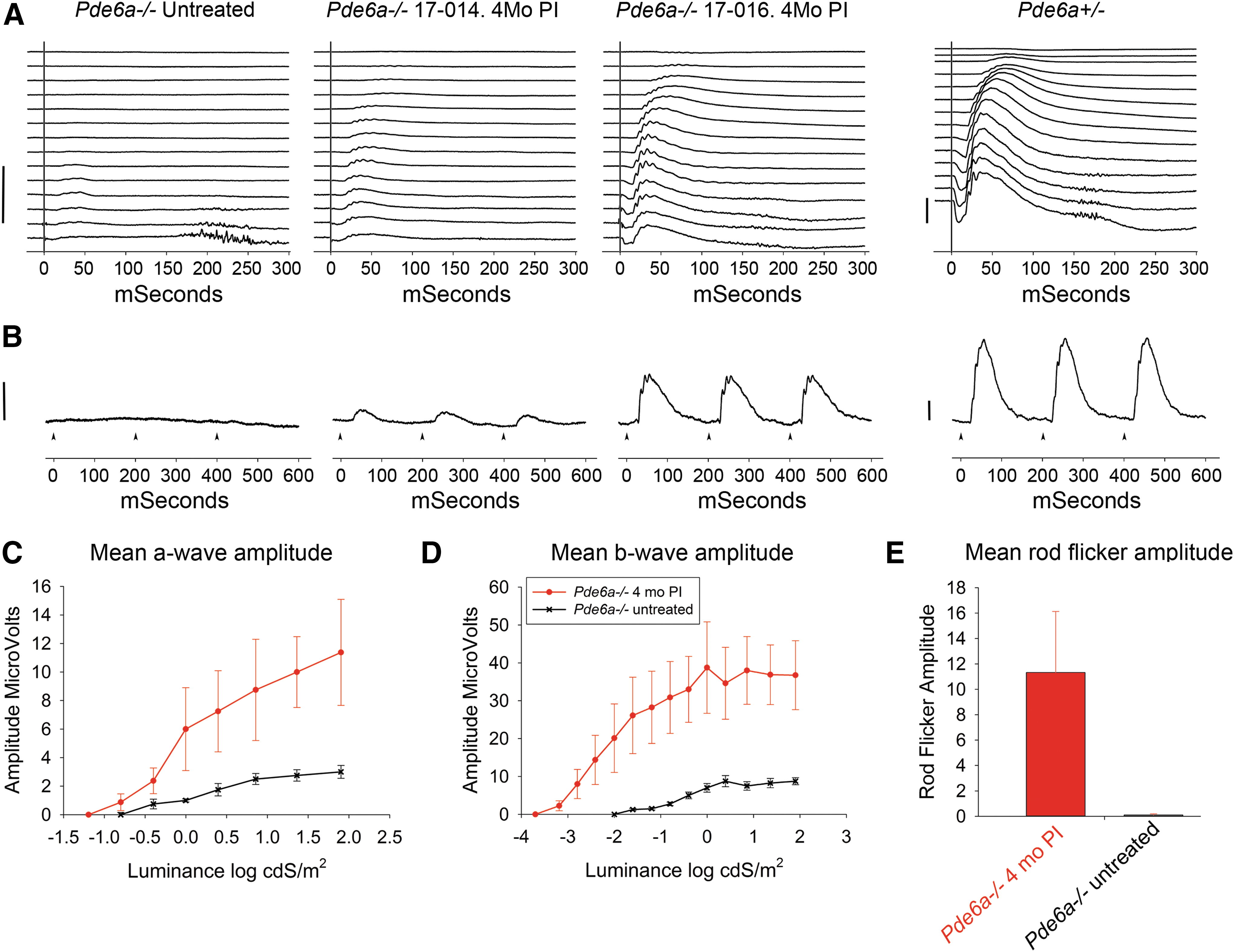
ERG results at 4 months post injection—high-dose group.
The three animals in the low-dose group retained in the study had variable degrees of improved rod function. All three had improved rod-mediated vision by 5 months post injection, as described below. However, only animal 16-024, which had the best injection, had an improvement in rod-mediated ERG, with a marked improvement in the dark-adapted ERG response threshold and in the a- and b-wave amplitudes. This rescue of rod function was present by the 2-month post injection time point, and was maintained at 5 months post injection (Supplementary Fig. S2). The treated eyes in the remaining two animals (16-023 and 16-025) did not show an increase in scotopic ERG amplitudes compared to the untreated control eyes (Supplementary Fig. S2). There was no significant difference in the amplitudes of the light-adapted ERGs between treated and control eyes during the study (data not shown).
Animal 16-024 showed a marked improvement in performance in the vision-testing device at the low (scotopic) light level, which exclusively tests rod-mediated vision when comparing the treated eye to untreated eyes at both 2 and 5 months post injection (Supplementary Figs. S3 and S4). Vision testing in the animal with the rapid flattening of the injection bleb (16-023) showed no improvement in visual performance at 2 months post injection but had improved performance (both in correct choice of exit and exit time) at 5 months post injection (Supplementary Figs. S3 and S4). Vision testing in animal 16-025 (which had a small but maintained bleb and a second larger bleb that flattened rapidly) showed improved scotopic vision at both 2 and 5 months post injection (Supplementary Figs. S3 and S4).
High-dose group
A second cohort of animals was injected with a higher titer, aiming to deliver a total dose of ∼5 × 1011 vg into the sub-retinal space. In two animals in this high-dose group (17-012 and 17-016), the sub-retinal injection resulted in the formation of a large bleb that was maintained at the post-injection imaging time point (Fig. 1B). The next two animals had less optimal results of injections in the first eye: in animals 17-014 and 17-015, the injection in the first eye resulted in a good sized bleb, but this had flattened by the time the post-injection imaging was performed. It was hypothesized that the rapid flattening of the bleb was due to reflux of the injected vector into the vitreous through the retinotomy. This would considerably shorten the contact time between vector and photoreceptors compared to the successful injections where the sub-retinal fluid is absorbed over the subsequent 1–2 days. In view of this hypothesis, and the findings of an association with a lack of ERG rescue in the low-dose group, rapid flattening of the bleb was considered to be suboptimal, and therefore sub-retinal injections were performed in the contralateral eye of those animals. The injections in the second eye of these two animals resulted in the formation of an acceptable bleb that remained present at the post-injection imaging. On fundus examination, all blebs resolved over the expected time course (the subsequent 24–48 h). The outcome measures of the eyes of the four animals with a successful sub-retinal injection not associated with rapid flattening were used for statistical analysis of measures at 2 and 4 months following injection. At 2 months, the scotopic ERGs from all eyes considered to have a successful injection showed evidence of rescue of rod function. Figure 2A and B shows the dark-adapted ERG waveforms at 4 months post injection from an eye with a moderate ERG rescue (animal 17-014, OS) and an eye with good ERG rescue (animal 17-016, OD) compared to an affected untreated representative control Pde6a–/– eye and an age-matched unaffected Pde6a+/– control animal/eye (which has ERG waveforms indistinguishable from wild-type dogs). There is an improvement in both response threshold and amplitudes. The mean ERG amplitudes are shown in Fig. 2C–E and show the improvement in response threshold (i.e., a lowering of stimulus response threshold of ∼0.5 and ∼1.5 log units for a- and b-waves, respectively) and marked increase in amplitudes (p < 0.01). Supplementary Figure S5 shows the rescue present at both time points and the results from individual puppies. There was a small but not statistically significant decrease in mean ERG amplitudes between the 2- and 4-month time points. As with the low-dose group, there was no significant difference in light-adapted ERG responses between treated and control eyes during the study (data not shown).
Vision-testing results for the high-dose group are shown in Fig. 3 and Supplementary Fig. S6. When using the successfully treated eyes at the lowest light levels, the animals chose the correct exit tunnel more frequently (close to 100% of the time) and were able to exit the device more rapidly compared to the results from untreated control eyes. At low lighting levels, untreated Pde6a–/– animals are unable to see the correct exit. At brighter lighting levels, the untreated puppies at this age can choose the correct exit tunnel because those lighting levels are within their cone functional range. There was a statistically significant improvement in both exit choice and time to exit in the treated eyes compared to age-matched untreated controls (p < 0.01). Two unilaterally treated animals (17-012 and 17-016) were also tested using an obstacle course under scotopic conditions, allowing rod-mediated vision from the treated and untreated eyes to be compared. The transit times were much faster with the treated eyes and were made with no collisions with obstacles. With the untreated eyes, the dogs were slow and hesitant and had several collisions. A video of the obstacle course testing of 17-016 is provided as Supplementary Video S1.

Vision testing at 4 months post injection—high-dose group.
Structural outcome measures
The results are summarized in Table 1.
Low-dose group
Fundus examination by indirect ophthalmoscopy, color fundus photography, and infrared cSLO imaging showed that the injected region of the retina demonstrated features indicating retinal preservation in all three injected eyes in the low-dose group. While the surrounding untreated retina and control eyes developed tapetal hyperreflectivity over the study period (up to 5 months post injection), this was not apparent within the injected area, which also showed preservation of superficial retinal vasculature (Fig. 1). In dogs, the amount of light reflection from the tapetum is an indicator of the degree of absorbance of light as it passes through the retina to be reflected back from the tapetum to the observer or the camera. Thinner retinas absorb less light, giving a fundus appearance known as tapetal hyperreflectivity, which is taken as a clinical indicator of retinal thinning. In vivo retinal imaging using SD-OCT confirmed that there was preservation of the outer retina in the region of the sub-retinal injection in all three animals (Fig. 4). In addition, the definition of the zones that represent the region of the photoreceptor inner and outer segments showed improved definition, but only in the injected region, indicating improved photoreceptor morphology (later confirmed on retinal sectioning). These zones include the more reflective zones of the external limiting membrane (ELM), ellipsoid zone (EZ), and interdigitation zone (IZ), with the less reflective zones between ELM and EZ and between EZ and IZ representing inner segments and outer segments, respectively. In untreated retinal regions and control untreated eyes, these zones could not be visualized. This improvement in the treated area was present in all three puppies, but it was most apparent in animal 16-024, which had the best sub-retinal injection outcome of the three animals in this group. For evaluation of photoreceptor survival, the REC+ thickness was measured. There was significant preservation of REC+ thickness in all three treated eyes in the low-dose group compared to control regions (Fig. 4A2). At 2 months post injection, REC+ thickness was significantly greater in the treated eyes (81.4 ± 6.5 μm) compared to the control (58.5 ± 5.7 μm; p = 0.002). At 5 months post injection, the thicknesses were 63.0 ± 9.3 versus 40.8 ± 4.6 μm (p = 0.015). The decline in mean REC+ thickness between 2 and 5 months post injection was statistically significant (p = 0.036). The heat maps in Fig. 4B1 show the preservation of REC+ thickness across the injected area. These are shown for the animal with the best outcome (16-024), but also for animal 16-023, which had a rapidly flattening bleb but showed improvement in visual function. With the latter animal, the REC+ thickness preservation was not so dramatic.
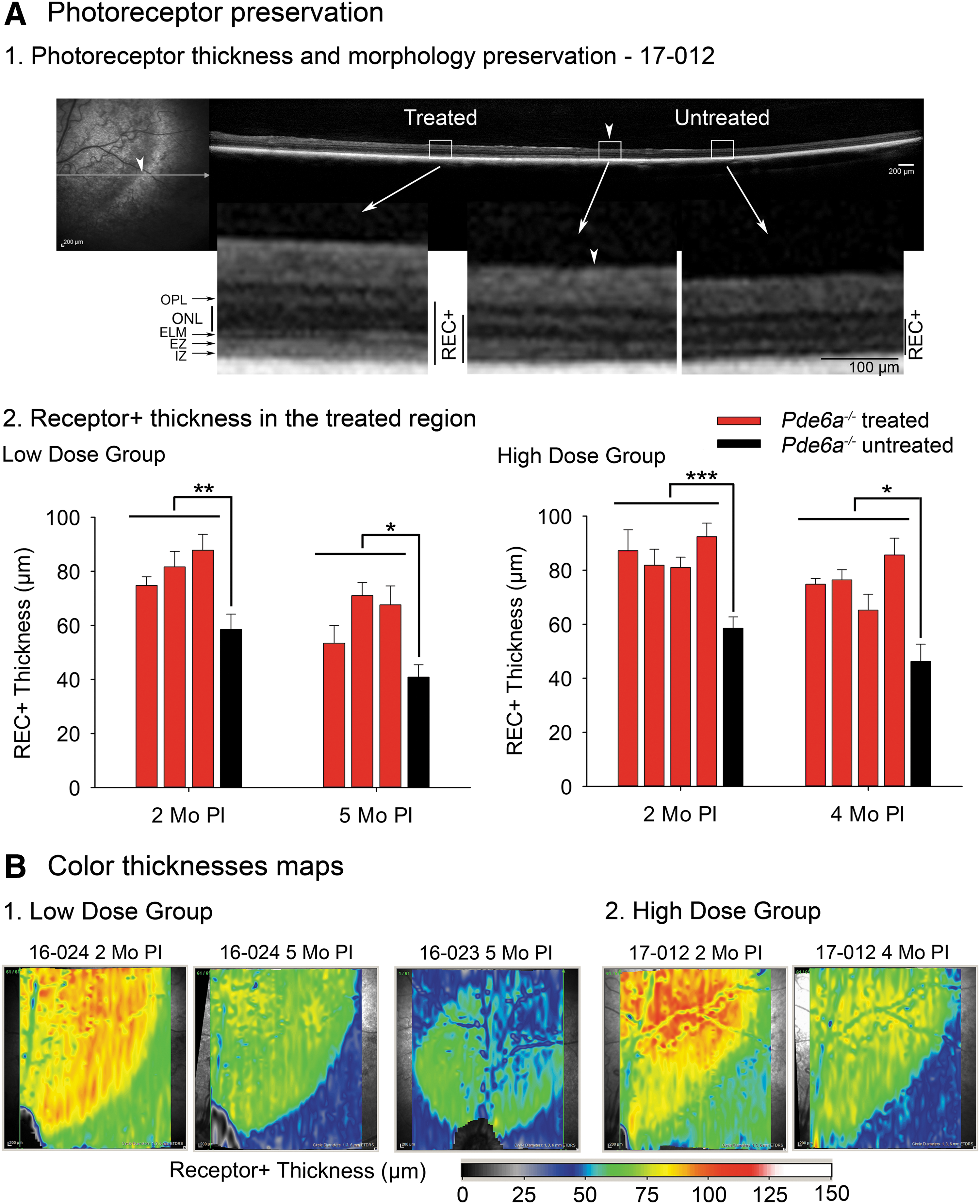
Preservation of photoreceptor thickness and morphology in the treated area.
High-dose group
Similar to the low-dose group, all the successfully treated eyes showed indicators of retinal preservation on indirect ophthalmoscopy, fundus color photography, and infrared cSLO. Again, the treated areas did not develop tapetal hyperreflectivity and showed superficial retinal blood vessel preservation (Fig. 1B). SD-OCT cross-section imaging showed improvement in zones representing photoreceptor inner and outer segments within the treated areas (Fig. 4A1). The mean REC+ thickness within the treated area was significantly preserved in all successfully treated eyes at 2 months (85.6 ± 5.3 vs. 58.5 ± 5.7 μm; p < 0.001) and 4 months (75.5 ± 8.4 vs. 46.2 ± 6.4 μm; p = 0.013; Figs. 4A2 and 4B2). As with the low-dose group, the mean REC+ thickness declined between the 2- and 4-month time points (p = 0.025).
Immunohistochemistry
Animals from the low-dose group were euthanized and their eyes processed for IHC after the 5-month post injection outcome measures had been collected. The animals in the high-dose group are being maintained for long-term monitoring of the duration of rescue. Retinal sections across the injection bleb were labeled with an anti-PDE6A antibody and showed PDE6A expression in the outer segment of photoreceptors in the treated regions of all three injected eyes, but not in the untreated regions of the same eye or in the contralateral untreated eyes (Fig. 5). In addition to appropriate PDE6A labeling (Fig. 5A), Pde6b expression in the outer segments was dramatically increased (Fig. 5B). In untreated regions of Pde6a–/– puppies, there was very low or absent labeling for Pde6b. Expression of the two active subunits of PDE6 resulted in reduced photoreceptor cyclic GMP (cGMP) levels, shown by an absence of labeling for cGMP in the treated region (Fig. 5A). Loss of PDE activity in the rod photoreceptors of Pde6a–/– puppies resulted in accumulation of cGMP in untreated regions. Sections across the junction between injected and uninjected retinal areas clearly showed the association of loss of cGMP labeling with expression of the transgene (Supplementary Fig. S7). In untreated retinal regions, rod opsin was markedly mislocalized to the outer nuclear layer. PDE6A transgene expression rescued this mislocalization, and in the treated region, rod opsin was predominantly expressed in rod outer segments (Supplementary Fig. S8). The treated retinal region also showed a reduced immunolabeling for glial acidic fibrillary acidic protein (GFAP) compared to the untreated retinal regions (Fig. 5B). GFAP upregulation, indicative of Müller cell activation, is a feature of this, as well as many other retinal degenerative conditions. 43 –45 Labeling of cone photoreceptors showed a dramatic improvement in cone inner and outer segment morphology in the treated region (Fig. 5C). In the untreated retina, cones at this age are stunted, with shortened inner and outer segments. In the treated retinal regions, cone inner and outer segments were markedly longer than in the untreated regions and control Pde6a–/– eyes.
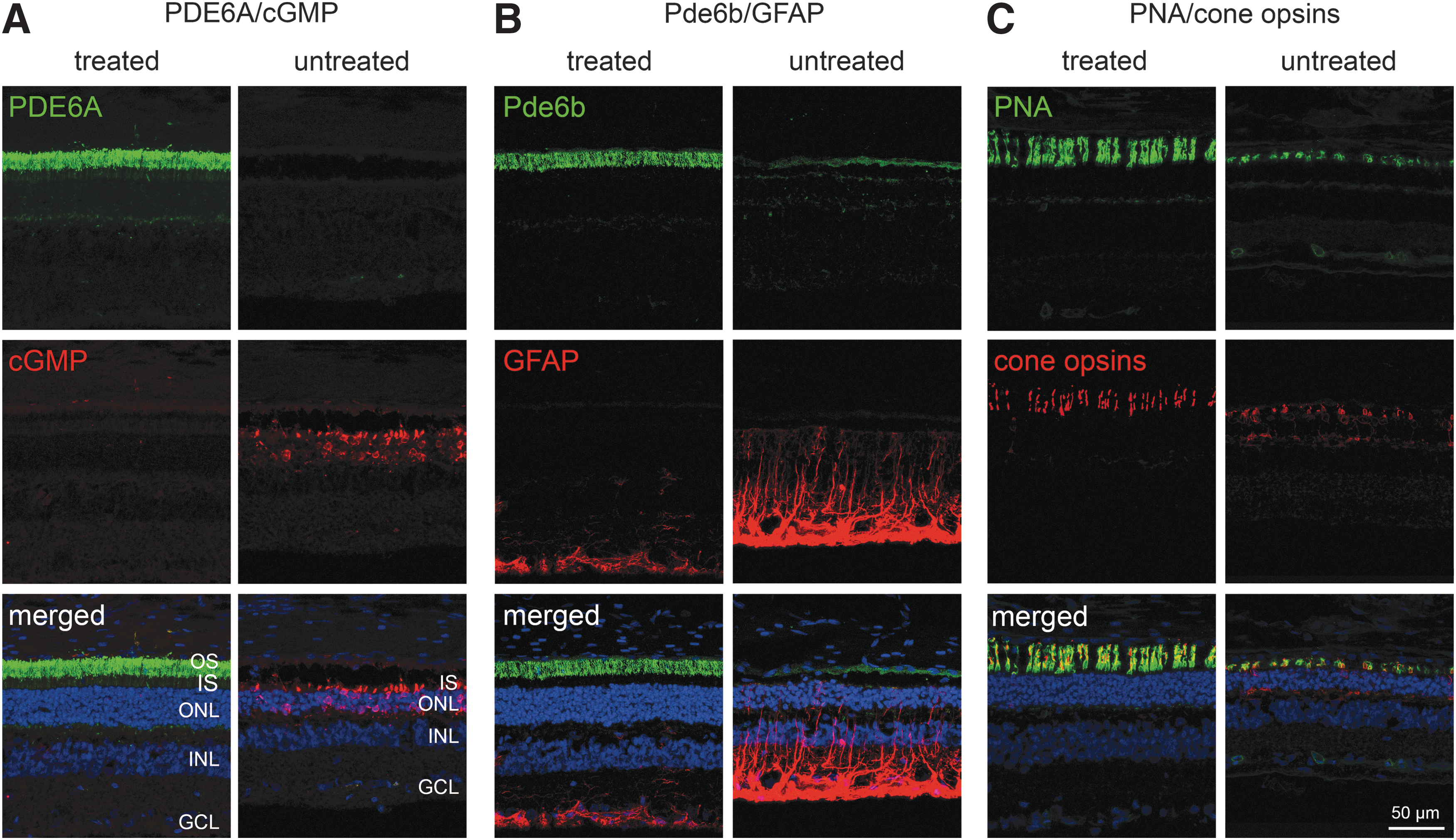
Immunohistochemical analysis of treated compared to untreated regions—low-dose group. Retinal sections images from animal 16-024. In all sections, the ONL thickness was preserved with more photoreceptor rows (blue) surviving in the treated compared to the untreated region.
Discussion
This study shows that gene supplementation therapy with an AAV8 vector delivering human PDE6A under control of a human rhodopsin promoter efficiently transduces rod photoreceptors in the Pde6a–/– dog model of PDE6A-RP, restoring their function. Untreated animals lack rod function. They are effectively blind at low lighting levels and only show cone-mediated ERG responses. 19,34 Some degree of rod rescue was apparent in all seven treated animals. This ranged from just an improvement in rod-mediated vision but no increase in dark-adapted ERG waveform amplitudes to improvement in both rod-mediated vision and ERG waveforms. The finding of improved vision without an improvement in ERG amplitudes indicates that vision testing using the four-choice vision-testing device can be a more sensitive indicator of improved retinal function than full-field ERG. This is in line with findings in previous studies of gene supplementation therapy in the Rpe65–/– dog. 36 There appear to be similar findings in human patients in RPE65 gene therapy trials where improved vision can be detected but where there is no recordable ERG improvement. 1 –3,36 Improvement in vision may result from contributions from a smaller number of functional photoreceptors than is needed to generate an electrical signal recordable when performing a full-field ERG. In the current study, only one animal in the lower-dose group had an improvement in ERG response threshold and amplitudes, whereas the entire high-dose group had an ERG improvement. However, following the observation in the low-dose group that two of the sub-retinal injection blebs flattened rapidly following injection and that those eyes appeared to have poorer functional rescue, this was considered to be a technical failure, and the second eye was injected when treating the subsequent cohort of puppies with the higher dose of vector. Because of this change in protocol between the two groups, the efficacy of the two doses cannot be directly compared. A complete dose-optimization study would be required to establish the most effective safe dose and is planned for the future.
The improvement in dark-adapted ERG response threshold and a- and b-wave amplitudes seen in one of the low-dose puppies and all of the high-dose puppies proves that there is rod function. This is an improvement on the initial proof-of-concept gene supplementation studies with the Pde6a–/– dog model where sub-retinal injection of an AAV8 with a 733 tyrosine to phenylalanine mutation delivering the canine Pde6a cDNA under control of a ubiquitous promoter restored rod vision but did not result in an improvement in ERG waveform threshold or amplitudes. 22 The current study demonstrates that the human PDE6A cDNA under control of a human rhodopsin promoter can substitute for canine Pde6a. This is important, as it allows the use of this large-animal PDE6a-RP model in preclinical safety and efficacy trials to help progress PDE6A gene supplementation therapy into the clinic.
Expression of the transgene also resulted in a major improvement in Pde6b expression, with appropriate localization to the outer segment, whereas in uninjected areas, there was very reduced or absent labeling for Pde6b. This demonstrates the importance of Pde6a expression for normal expression of Pde6b. Concurrent expression of both alpha and beta Pde6 subunits is required for the formation of a function PDE6 heterotrimer. The treatment also resulted in preservation of retinal structure detectable by both in vivo imaging (Fig. 4) and also in the retinal sections used for IHC (Fig. 5). The treated regions had improved definition of the SD-OCT layers that represent the inner and outer segments of the photoreceptors. IHC showed that this in vivo indicator of photoreceptor health translated into improved morphology detectable in the retinal sections used for IHC. The REC+ thickness measurement from SD-OCT images was used, as this encompasses the entire length of the photoreceptors. This layer was significantly preserved in the treated region of all successfully treated eyes. Heat maps representing the REC+ thickness clearly show the preservation of this layer in the treated retinal regions. This was even apparent in eyes with a suboptimal sub-retinal injection. In those eyes, the preservation was associated with PDE6A labeling of photoreceptors detected by IHC. A decrease in REC+ thickness was noted between the two time points at which it was assessed. The intention is to continue to monitor the high-dose group to see if the preservation of REC+ thickness is maintained. In a gene supplementation study in a different canine RP model (due to a mutation in Cngb1, which results in a slower retinal degeneration), it was noted that there was an initial continued loss of REC+ thickness in the treated retinal regions, and that after a period of a few months, this plateaued. 46 It is suspected that an initial decline in REC+ thickness occurred because of the death of rods that were not transduced had inadequate levels of transduction or were already destined to die. Longer-term monitoring of the high-dose dogs will show if the same halting in loss of REC+ thickness occurs with the more severe retinal degeneration seen in the Pde6a–/– dog. The retinal preservation in the treated retinal regions became apparent on fundus examination, as the preserved retina did not develop the tapetal hyperreflectivity and superficial retinal blood vessel attenuation that is a feature of the untreated retinal regions. This could also be detected in the color fundus images, as well as the infrared cSLO images (Figs. 1 and 4A1). The IHC study (Fig. 5) showed that, in addition to preservation of rods, cone morphology was much improved. In the Pde6a–/– dog, cones are stunted from an early age and progressively die secondarily to the rod loss. 19 In the treated region, cones appeared to have a much more normal morphology, with well-developed inner and outer segments (Fig. 5C). This improvement in the morphology of the inner and outer segments of the photoreceptors seen on retinal sections could be detected by SD-OCT in vivo as improved definition of the zones representing these regions. Longer-term studies will determine if cone improvement is maintained in the treated regions. If this is the case, cone functional preservation will be detectable by ERG in the longer term because there is a progressive decline in cone ERGs with progression of the condition in the untreated eye.
This study shows improved results of gene supplementation therapy with the PDE6A-RP dog model. A vector construct that may be translatable to human clinical trials was shown to be effective in restoring rod function and in preserving retinal structure. Importantly, this was not associated with the adverse effects previously found with gene supplementation therapy in this model, where retinal rosette formation and patches of retinal degeneration were seen in addition to the regions of retinal preservation. 22 The reason for this difference between the two studies is not clear, as there were several differences between the constructs, including in AAV serotype, promoter (tissue specific in the current study), and the addition of a WPRE to the expression cassette in the current study. Further important studies are required to confirm the safety of this viral vector, to establish an optimal dose, and to investigate whether rescue is possible at later disease stages. The phenotype in the dog is of a rapid photoreceptor loss, meaning that there is the opportunity to study the effect of intervention at different stages of retinal degeneration within a practical timeframe. This is a very important aspect to investigate because it is likely that human patients may have significant photoreceptor loss when they present for gene supplementation therapy. A genetic study by Tsang et al. with a PDE6 mouse model suggests that if adequate Pde6B expression is achieved, photoreceptor degeneration may also be halted at later stages of disease. 47 The large-animal model of RPE43 used in the current study will be valuable for the additional preclinical safety and efficacy trials needed prior to a human clinical trial.
Footnotes
Acknowledgments
The authors would like to thank Janice Querubin for her help with ERG and vision testing, Heather Defore for her help with anesthesia and animal care, and Kerstin Skokann for excellent technical help. This work was supported by Tistou and Charlotte Kerstan Stiftung and the Myers Dunlap Endowment for Canine Health.
Author Disclosure
C.S., M.W.S., M.B., and S.M. are part of a patent application related to this publication. L.M.O. and S.M.P.-J. have no competing financial interests.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.