
Abstract
Striated muscle wasting occurs with a variety of disease indications, contributing to mortality and compromising life quality. Recent studies indicate that the recombinant adeno-associated virus (serotype 6) Smad7 gene therapeutic, AVGN7, enhances skeletal and cardiac muscle mass and prevents cancer-induced wasting of both tissues. This is accomplished by attenuating ActRIIb intracellular signaling and, as a result, the physiological actions of myostatin and other ActRIIb ligands. AVGN7 also enhances isolated skeletal muscle twitch force, but is unknown to improve systemic muscle function similarly, especially exercise capacity. A 2-month-long dose-escalation study was therefore conducted using 5 × 1011, 1 × 1012, and 5 × 1012 vg/mouse and different tests of systemic muscle function. Body mass, skeletal muscle mass, heart mass, and forelimb grip strength were all increased in a dose-dependent manner, as was the fiber cross-sectional area of tibialis anterior muscles. Maximal oxygen consumption (VO2max), a measure of metabolic rate, was similarly enhanced during forced treadmill running, and although the total distance traveled was only elevated by the highest dose, all doses reduced the energy expenditure rate compared to control mice injected with an empty vector. Such improvements in VO2max are consistent with physiological cardiac hypertrophy, which is highly beneficial and a normal adaptive response to exercise. This was particularly evident at the lowest dose tested, which had minimal significant effects on skeletal muscle mass and/or function, but increased heart weight and exercise capacity. These results together suggest that AVGN7 enhances striated muscle mass and systemic muscle function. They also define minimally effective and optimal doses for future preclinical trials and toxicology studies and in turn will aid in establishing dose ranges for clinical trials.
Introduction
T
Cancer is just one of many disease indications associated with muscle wasting, which also occurs with muscular dystrophies and myopathies, heart failure, chronic obstructive pulmonary disease (COPD), chronic kidney disease (CKD), peripheral neuropathies, and nerve damage, as well as with age-related sarcopenia. 16,17 In fact, more than half of people >80 years of age suffer from sarcopenia, which progressively develops 18 and significantly increases the risk of hospitalization, disability, and mortality. Sarcopenia is also the primary determinant for developing frailty syndrome 19,20 and costs the healthcare system an excess of $183 million in nursing-home expenses alone, 21 which is just a fraction of the estimated total costs for treating muscle wasting in general. Thus, novel therapeutics capable of preventing muscle wasting and of restoring muscle mass and function after wasting has occurred could potentially transform the treatment of many diverse disease indications.
Several notable studies suggest that attenuating the actions of myostatin, a myokine that inhibits processes required to maintain striated muscle mass, 22 could potentially help in treating muscle wasting disease. 23 –25 Circulating myostatin levels are elevated in many of the aforementioned states and thus, may contribute to the muscle wasting that develops with each. 26 –29 In fact, activation of myostatin/activin receptors (ActRIIb) inhibits processes normally associated with muscle fiber hypertrophy and simultaneously stimulates muscle atrophy via the activation of the E3 ubiquitin ligases MuRF1 and MAFbx. 30 –32 The promiscuity of ActRIIb receptors for circulating ligands in addition to myostatin, namely growth/differentiating factor (GDF)11 and activin, may have little physiological relevance, but could potentially contribute to muscle wasting in some catabolic conditions, as ActRIIb activation independent of ligand induces muscle atrophy, while ActRIIb immunoneutralization stimulates muscle hypertrophy. 33 –36
Several developing therapeutics for muscle wasting diseases have targeted myostatin or ActRIIb. 1,37 Despite early successes with preclinical studies, clinical trials with these drugs have produced either mixed results or serious off-target effects that compromised blood vessel integrity. 38 A novel gene therapeutic using recombinant adeno-associated virus vectors with high tropism for striated muscle (serotype 6) that overexpresses Smad7 in these tissues (rAAV6:Smad7; i.e., AVGN7) was therefore developed. 39 This targeted approach avoids the non-specific side effects of attenuating ActRIIb ligands in the circulation and, in addition, significantly increases muscle mass and strength in healthy mice. Furthermore, AVGN7 prevented cancer-induced muscle wasting in mice bearing C-26 tumors and in mice co-injected with AAV6 vectors carrying myostatin or activin cDNA. It also restored muscle mass after cachexia was induced and attenuated ActRIIb signaling and the expression of MuRF1 and MAFbx.
Although AVGN7 was demonstrated to increase isolated muscle twitch force in healthy and tumor-bearing mice, systemic or in vivo muscle function was not assessed and only a single vector titer with systemic delivery was tested. The current studies were therefore performed to identify a minimal effective dose and to determine if AVGN7 could enhance systemic muscle function in vivo. The results are the first to demonstrate enhanced exercise capacity with myostatin/ActRIIb attenuation. They also suggest that the cardiac hypertrophy that develops from attenuating these pathways pharmacologically is consistent with physiologic and not pathologic hypertrophy.
Materials and Methods
Vector synthesis and animal maintenance
AVGN7 (i.e., rAAV6:Smad7) and an rAAV6 empty vector (EV) control were synthesized in the Research Vector Core of the Raymond G. Perelman Center for Molecular and Cellular Therapeutics, Children Hospital of Philadelphia. The expression construct contained a CMV promoter, mouse smad7 cDNA, and the SV40 poly-A region, all of which were flanked by two AAV2 inverted terminal repeats, as previously described. 39 Vector quality control included proprietary bioburden, and endotoxin assays and genome titers were determined using a proprietary quantitative polymerase chain reaction assay. Sterile vector was supplied in phosphate-buffered saline (PBS) with 0.001% Pluronic F-68 and was ultimately diluted in PBS to a working concentration before injecting.
All animal procedures were preapproved by the Institutional Animal Care and Use Committee (IACUC) at Washington State University. Mice were housed in an environmentally controlled room on a 12 h light:dark cycle with access to food and water ad libitum. Female C57BL/6 mice (3 months old) were selected from an established breeding colony and randomly assigned to one of four groups receiving 0, 5 × 1011, 1 × 1012, or 5 × 1012 vector genomes (vg)/mouse. For systemic delivery of AVGN7, mice were first anesthetized with a single intraperitoneal injection of 250 mg/kg of freshly prepared 2,2,2-tribromoethanol. Stock AVGN7 was diluted in sterile PBS and mice were injected retro-orbitally (r.o.) with a volume of 150 μL.
Assessing grip strength
Forelimb grip strength was quantified using a Columbus Instruments Grip Strength Meter and a T-bar attachment. Assays were performed just before mice were injected and 1 and 2 months after. Mice were held by their tails, and after both paws gripped the bar, mice were gently pulled backward until their grip was broken. This assay was performed five times for each mouse, with 1 min intervals between measures and while rotating between individual mice with consecutive measures. For each time point, individual mean values were then used to calculate a single representative value/mouse and these values were used as in-group replicates.
Assessing exercise capacity
Forced treadmill running and VO2max tests were used to assess exercise capacity on a four-lane Oxymax FAST Modular Treadmill System from Columbus Instruments (Columbus, OH), as previously described. 40 The chamber air circulation rate was set at 0.6 L/min for all tests and the shock unit settings was 2.5, which corresponded to a frequency, duration, and intensity of 1 Hz, 200 ms, and 0.65 mA, respectively. Before testing the mice, they were first introduced to the chambers and treadmills and exposed to the shock apparatus with three consecutive days of acclimation training. This consisted of 5 min on a stationary treadmill followed by an additional 5 min of walking at a speed of 5 m/min. Shock accumulation was monitored with each session and acclimation was verified by the reduction of shocks over time. Mice were considered acclimated when receiving three or fewer shocks.
VO2max tests were performed the day before injecting mice and again 2 months after injecting. They began by once again introducing mice to a stationary treadmill set at a 25° slope. After 5 min, the treadmill speed was then programmed to increase to 5, 9, 12, and 15 m/min with 5 min intervals. The speed then increased by 1.8 m/min every 2 min until the mice became exhausted, as indicated by their failure to reengage the treadmill for 10 consecutive seconds. The VO2max for each mouse was identified by a peak VO2 (mL/kg/h) and a concurrent and sharp rise in the respiratory exchange ratio (RER), which also occurred just before exhaustion. Shock units were then deactivated to allow the exhausted mice to rest while other mice continued running. Caloric expenditure was calculated using CV = 3.815 + 1.232 × RER, which was derived from the linear positive correlation between heat per liter O2 (kcal). This in turn was calculated using Heat = CV × VO2, RER, and total energy expenditure (kcal).
Assessing muscle morphology and histology
Mice were euthanized 24 h after the final VO2max test. Tibialis anterior, gastrocnemius, and quadriceps skeletal muscles were removed, weighed, and flash frozen in isopentane cooled to −140°C. Frozen muscles were then sectioned (6–8/muscle) at a thickness of 10 μm on a cryostat and stored at −80°C. Sections were fixed for 10 min in 4% paraformaldehyde/PBS and then stained with hematoxylin and eosin (H&E). Cross-sectional area and feret diameter were quantified on at least four images/muscle using ImageJ. These values were used to calculate means representing each individual mouse, which in turn were used to calculate group means. Hearts were removed, cleared of non-cardiac tissue, cut open, and rinsed in PBS to remove blood clots. They were then blotted dry before weighing. Tibia were also removed and their lengths were used to normalize heart mass.
Western blotting
Protein was extracted from frozen tibialis anterior muscles that were first powdered in liquid N2 before homogenizing in lysis buffer (1% SDS; 10 mM of Tris-HCl, pH 8.0; 100 mM of NaCl; 3 mM of MgCl2; 0.5% NP-40; 10 mM of NaF). Each homogenate was diluted in equal volumes of 2 × SDS sample loading buffer and polyacrylamide gel electrophoresis was performed using 5–20% acrylamide gradient gels. Proteins were then electro-transferred to nitrocellulose membranes, which were then blocked in 5% nonfat dry milk/TBST (0.1% Tween-20; 50 mM of Tris-HCl, pH 7.6; 150 mM of NaCl) for 1 h at room temperature. Subsequently, membranes were incubated overnight in 5% milk/TBST that contained primary antibodies for Smad7 (2 μg/mL; 42-0400; Thermo Fisher Scientific) and β-actin (1:1,000; JLA20; Developmental Studies Hybridoma Bank). After washing three times for 5 min in TBST, membranes were incubated with secondary antibodies (IRDye 800CW goat anti-rabbit and IRDye 680 goat anti-mouse; LI-COR Biosciences) in 1% milk/TBST, rewashed, and then imaged using the Odyssey Infrared Imaging System (LI-COR Biosciences). Samples from each group were distributed equally on each of two gels and the Smad7 band intensity was normalized to that of β-actin for each sample.
Statistical analysis
Differences between means were determined using Prism software (GraphPad Software) to perform either a one- or two-way analysis of variance coupled to Tukey's post hoc test for multiple mean comparisons. Data are expressed as the mean ± standard error of the mean and differences were considered significant at p ≤ 0.05. In the figures, differences between means are often indicated by letters. Any letter shared between groups indicates no difference, whereas different letters indicate significance.
Results
Smad7 protein levels and body mass
Relative Smad7 protein levels (normalized to those of β-actin) were assessed in tibialis anterior muscles only. In mice injected with 1 × 1012 and 5 × 1012 vg AVGN7, Smad7 protein levels were approximately 150% and 200%, respectively, of those in muscles from control mice injected with an empty vector (Fig. 1A). These levels were significantly different from each other and from controls, although Smad7 levels in the latter were similar to those in mice injected with 5 × 1011 vg AVGN7. Such changes in muscle Smad7 protein had a profound effect on body mass and on the mass of individual skeletal muscles and the heart.
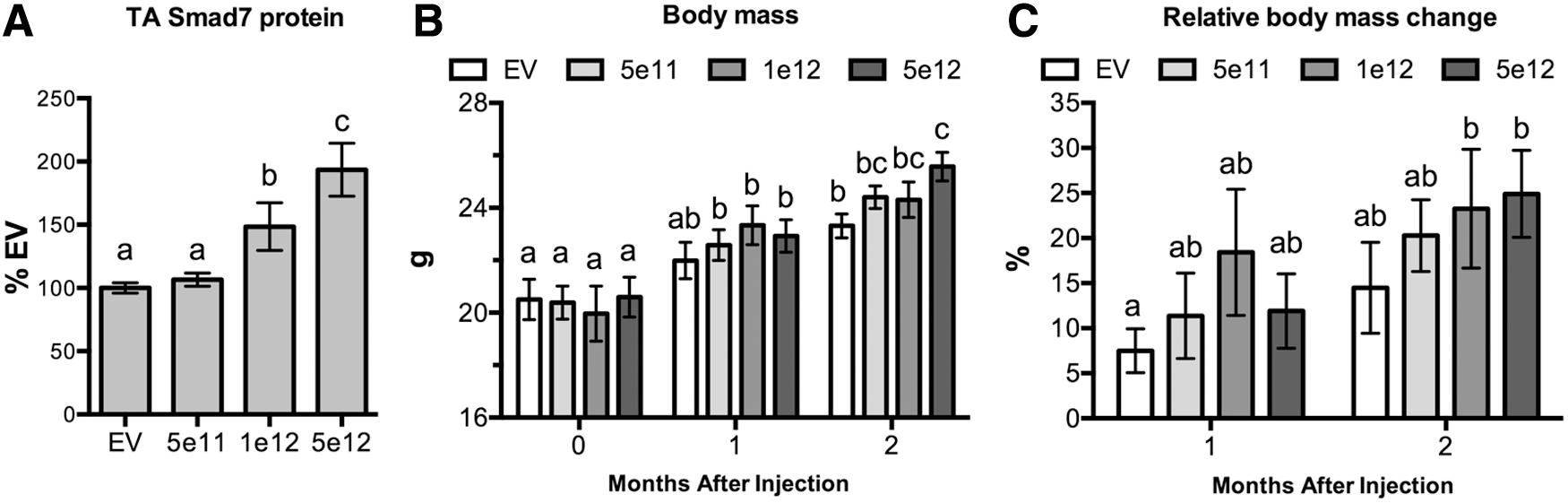
Dose-responsive changes in body mass. Two-month-old mice were injected PO with the following titers of AVGN7 (rAAV6:Smad7): 5 × 1011 (5e11), 1 × 1012 (1e12), and 5 × 1012 (5e12). Control mice were injected with 5e12 of empty vectors (EV).
The mean body mass of each randomly assigned group was similar at the beginning of the study (Fig. 1B). Although the body mass of treated mice, irrespective of injected dose, was not different from controls after 1 month, all doses of AVGN7 increased body mass over the initial values. This stimulatory effect was even more evident after 2 months, as the mass of mice injected with 5 × 1012 vg AVGN7 was significantly greater than that of control mice. These differences resulted from a 25% increase in mice injected with the higher two doses compared to 15% in control mice (Fig. 1C).
Striated muscle mass and cross-sectional area
Not surprisingly, more differences between groups were detected when comparing the mass of different skeletal muscles, including the tibialis anterior, gastrocnemius, and quadriceps, as well as the heart (Fig. 2A–D). Indeed, AVGN7 induced a dose-responsive increase in both skeletal and cardiac muscle mass, as multiple differences in each tissue examined were detected between individual doses and between each dose and controls. The greatest relative difference between the highest dose tested and controls, as well as the largest differences between doses, occurred in the tibialis anterior followed by the quadriceps (Fig. 2E–G). Both muscles are almost exclusively composed of type II fibers, whereas the gastrocnemius is a mixed-fiber muscle.

Dose-dependent differences in muscle weights.
The heart was also sensitive to treatment with dose-dependent increases in normalized heart mass occurring at the higher two doses (Fig. 2D and H). In fact, the relative change in heart mass increased significantly with each dose (Fig. 2H) and this did not occur when assessing changes in any skeletal muscle. This apparent heightened sensitivity to AVGN7 likely reflects the preferential tropism of AAV6 vectors for cardiac versus skeletal muscle. 41 It also supports previous studies reporting physiologic cardiac hypertrophy in the hearts of myostatin-null mice 42 –44 and other studies indicating that myostatin and/or ActRIIb attenuation stimulates cardiac hypertrophy. 39,45
Increases in skeletal muscle fiber cross-sectional area were previously reported when AVGN7 was injected intravenously or when tibialis anterior muscles were injected intramuscularly. 39 Similar results were obtained in the current study, as dose-dependent increases in cross-sectional area were clearly evident with H&E staining while, in addition, muscle morphology appeared normal (Fig. 3A–D). No differences were detected in the number of total nuclei, muscle fiber nuclei, or muscle central nuclei, suggesting that these markers of muscle regeneration were unaffected by AVGN7.
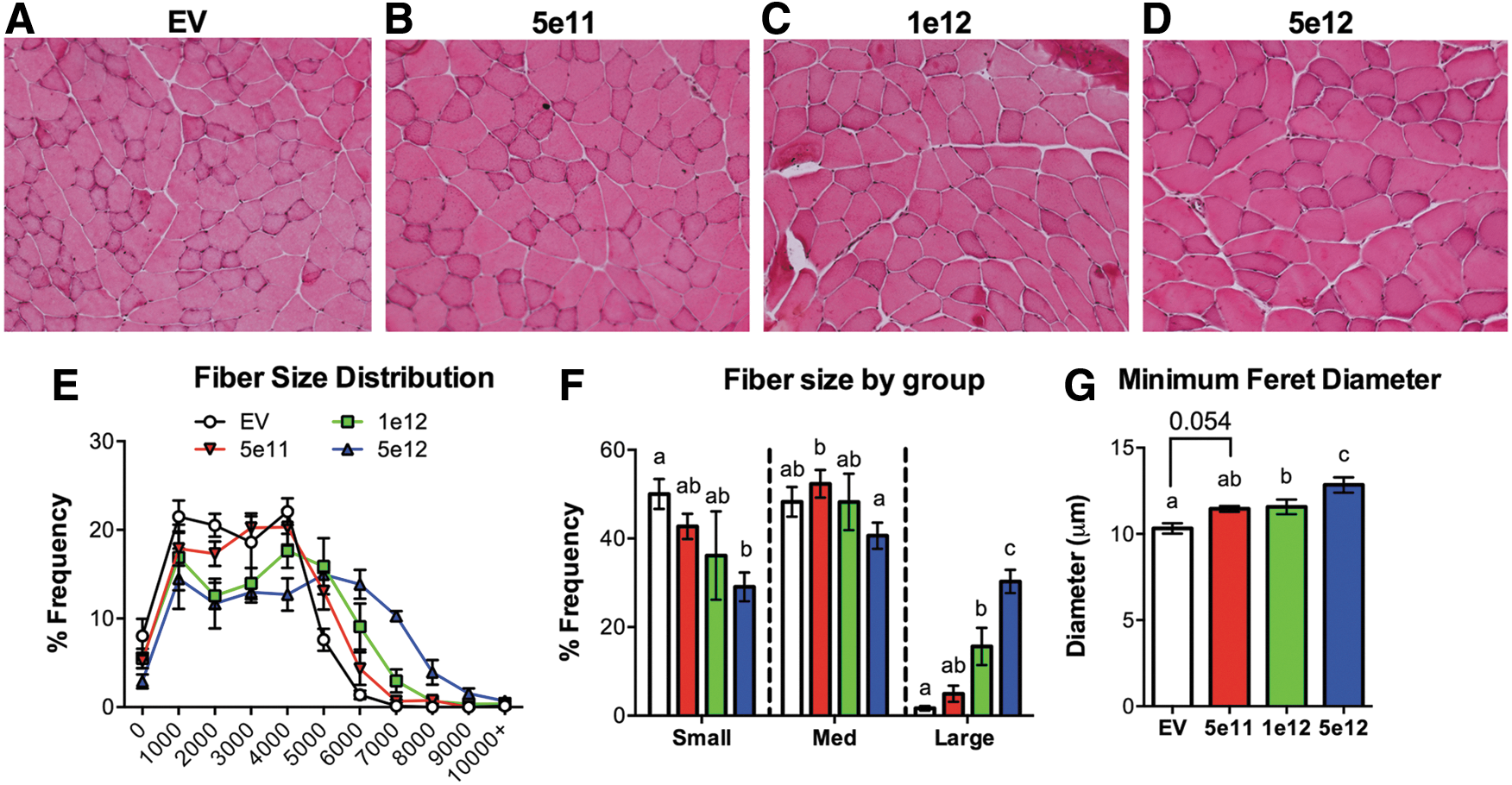
Dose-dependent differences in muscle fiber cross-sectional area.
Plotting the fiber size distribution revealed a shift toward an increase in large fibers in the AVGN7-treated mice, as the distribution curves skewed negatively as the dose increased (Fig. 3E). Pairwise comparisons of binned fiber sizes indicated that there were indeed significantly fewer small fibers and an increase in large fibers in AVGN7-treated mice (Fig. 3F). This positive dose effect was also reflected in the minimum feret diameter (Fig. 3G), an alternative measure of fiber size. Despite the fact that in each assessment, no significant difference was detected when comparing the 5e11 and 1e12 dose groups, non-significant trends consistent with a dose response were evident.
Grip strength
The only significant difference in absolute grip strength detected after 1 month of treatment occurred in mice injected with the highest dose, as mean levels in these mice were significantly higher than in all other groups (Fig. 4A). Nevertheless, a dose–response pattern was detected after 2 months where the highest two doses were significantly different from one another and from control mice. Comparing the relative changes between months further demonstrated a dose–response relationship even in the absence of significant differences in absolute strength, as for example in mice injected with 1 × 1012 vg. Absolute strength in these mice was similar to that of mice injected with an empty vector or 5 × 1011 vg after 1 month. However, the relative change was significantly greater than controls yet not different from the other groups. This “intermediate” effect was lost by 2 months, as grip strength was greater than control or low-dose mice and similar to that of high-dose mice. Such a progressive development of significance is typically seen in dose–response curves and demonstrates assay sensitivity at sub-maximal levels. The lack of an effect in mice injected with 5 × 1011 vg reflects the similar lack of differences in muscle and body mass when compared to controls and is not surprising when considering that TA Smad7 levels were not significantly elevated above those in control mice.
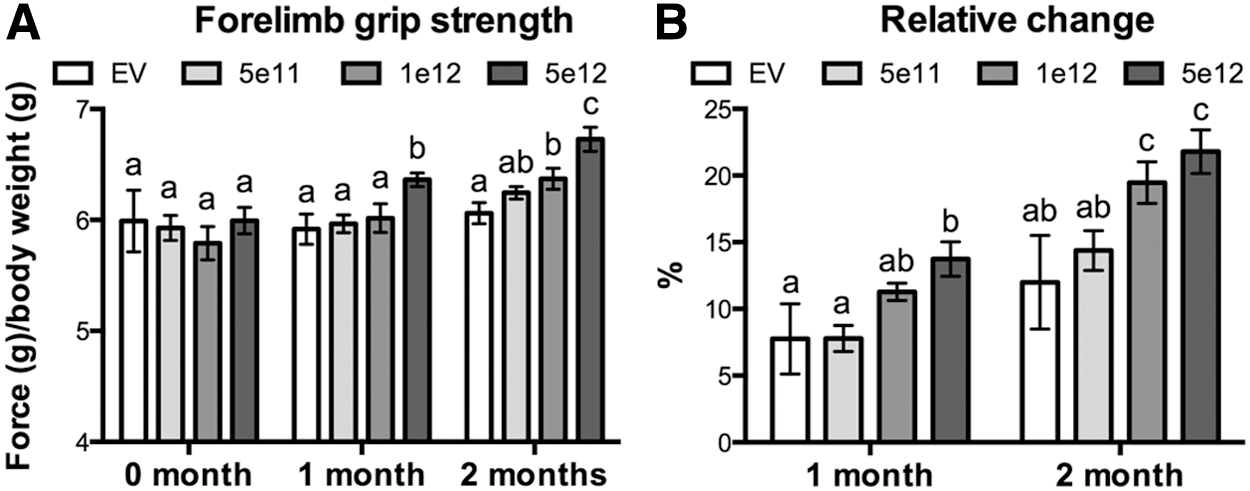
Dose-dependent changes in forelimb grip strength.
Exercise capacity
The VO2max of treated mice was significantly greater than controls at all doses, although the relative change between VO2max tests only increased in mice receiving the higher two doses (Fig. 5A and B). These latter differences were substantial and were four- to fivefold higher than the change in control mice. Treating mice with AVGN7 also increased running distance and the relative change between tests in a dose-responsive manner (Fig. 5C and D). This reflects a similar improvement with AVGN7 treatment, although significance was only demonstrated at the highest dose tested. The total energy expenditure of treated mice, except for those receiving 1 × 1012 vg, was less than that of controls (Fig. 5E and F). Moreover, energy expenditure in control mice increased by 20% between tests, whereas it decreased or stayed the same in treated mice, despite differences in running distance. This resulted in significantly different energy expenditure rates, which were lower than controls in all AVGN7-treated mice (Fig. 5G and H). This is probably best illustrated by the relative change in the expenditure rate, as it increased between tests in control mice and decreased in AVGN7-treated mice. These data together suggest that AVGN7 increases exercise capacity by increasing VO2max and this allows mice to run farther and more efficiently.

Dose-dependent changes in exercise capacity.
Discussion
The data herein provide the first evidence of enhanced exercise performance with ActRIIb attenuation, whether by Smad7 overexpression or by directly attenuating ActRIIb receptors or their ligands. The studies also establish a broad dose–response range that can be further refined for future toxicology studies and validate an additional route of administration (retro-orbital injections) for systemic delivery of AVGN7 in murine models. Body mass, muscle mass and cross-sectional area, heart mass, grip strength, VO2max, and running distance all increased with AVGN7 treatment, while the total calories expended during forced exercise as well as the energy expenditure rate both decreased. Some of these data are also the opposite of those obtained with Smad7-null mice where muscle mass and cross-sectional area as well as forelimb grip strength are all lower than those of wild-type mice. 46 These results together indicate that systemic treatment with AVGN7 increases exercise capacity in healthy mice and, considering previous studies, has the potential to influence skeletal and cardiac muscle function similarly in pathologic models.
Previous studies determined that AVGN7 (i.e., rAAV6:Smad7) can prevent tumor-induced muscle wasting in two different models of cancer cachexia. 39 It also enhances muscle mass and twitch force proportionally, whether administered locally or systemically, and can prevent myostatin-, activin-, or Smad3-induced muscle atrophy. Mechanistically, Smad7 overexpression prevented Smad3 phosphorylation and activation and downregulated the E3 ubiquitin ligases MuRF1 and MAFbx. This consequently prevented protein degradation and increased protein synthesis. It also increased the number and size of type IIa skeletal muscle fibers and the size of type IIb/x fibers.
Such changes in fiber type size and distribution are consistent with data reported herein, as fast-twitch muscles with a greater percentage of type II fibers (tibialis anterior and quadriceps) were more responsive than those with a lower percentage (gastrocnemius). All of these changes are also opposed by the Smad7-null mouse phenotype, which has a higher percentage of oxidative type I fibers and a lower percentage of glycolytic type II fibers in different muscles. 46 The slow-twitch soleus muscles of Smad7-null mice are similar in mass to those of wild-type mice. This further suggests that fast-twitch muscles are possibly more likely to be affected by AVGN7, although additional testing of slow-twitch muscles are needed. In addition, Cohen et al. used a muscle injury model and proliferation studies with primary muscle satellite cells to demonstrate that muscle regeneration is delayed in Smad7-null mice. 46 These data possibly suggest that AVGN7 has the potential to enhance muscle regeneration in response to injury, although this has yet to be demonstrated.
Mice were previously injected intravenously via the tail vein with 5 × 1012 vg. This increased Smad7 protein levels approximately twofold in tibialis anterior muscles, which is comparable to data reported herein (Fig. 1). Changes in muscle mass were also similar, as 20–50% increases in muscle mass were previously reported, depending on the muscle, while the same vector dose increased muscle mass by 20–40% in the current study. However, the previous study lasted 4 weeks, whereas the current study lasted 8 weeks. The better response in the previous study could be due to the route of administration, intravenous versus retro-orbital in the current study, or to many other factors including mouse strain (BALB/c vs. C57BL/6) or size (older/larger mice used previously) or even to normal inter-assay variability. Notwithstanding, obtaining significant results despite these differences helps to validate each study, especially as vectors were synthesized at different facilities.
Dose responsiveness was demonstrated in nearly every metric assessed, with the highest dose being optimal and producing significant differences in all metrics and at all time points. The relative efficacy, however, of each dose often varied between metrics and this resulted in metric-specific minimal effective doses. For example, the minimal effective dose for maximizing body mass, muscle mass, and grip strength was 1 × 1012 vg, while for influencing heart mass and exercise capacity, it was 5 × 1011 vg. Higher doses would likely be even more effective, as a saturation effect was not demonstrated. Moreover, longer treatment periods, even with low doses, would likely prove more efficacious as changes in muscle mass and strength accumulate.
This is an important consideration given current limits to the commercial manufacturing capacity of AAV-based gene therapeutics that are just now beginning to approach the scale needed to support disease indications with large patient populations or those that require large therapeutic doses. 47 Using traditional scaling, 48 the estimated human dose range that is comparable to the range tested herein is approximately 1 × 1012–1 × 1013 vg/kg, requiring approximately 8.5 × 1013–8.5 × 1014 vg to treat an average adult male cancer patient (50th percentile, 40–65 years old, weighing 82 kg), for example, or 1.8 × 1013–1.8 × 1014 vg to treat an average 5-year-old boy for Duchenne muscular dystrophy. Commercial production of clinical-grade AAV vector (i.e., following good manufacturing practice guidelines) is now approaching 1 × 1017 vg/batch and this could support up to 5,555 doses of AVGN7 for every batch, depending upon the clinical need (i.e., disease indication, patient characteristics, and dose). This highlights the importance of establishing human equivalent minimal effective doses for different patient populations, as this information is not only necessary for clinical trials, but will also help prioritize targeted disease indications. More importantly, it suggests that the current industry manufacturing potential can meet most of the development and clinical requirements.
In an untrained animal, cardiac function/performance is the primary factor influencing VO2max during high-intensity exercise, as the influences of improved skeletal muscle performance generally occur after cardiac performance is maximized. 49 The lowest AVGN7 titer significantly enhanced VO2max and energy expenditure metrics without consistently increasing muscle mass, body mass, or grip strength. These former changes are therefore due to enhanced performance of cardiac rather than skeletal muscle. This is supported by the well-established preferential tropism of AAV6 vectors for cardiac over skeletal muscle 41 and suggests that the AVGN7-induced cardiac hypertrophy is physiologic rather than pathologic. Cardiac output and responsiveness to β-agonists, cardiomyocite contractility, Ca2+ transients, and Ca2+ cellular loads, as well as cardiac muscle SERCA2a levels, are all enhanced or elevated in myostatin-null mice, 37,42 –44 which is analogous to treating with AVGN7. Myostatin also inhibits cardiomyocyte size and protein synthesis in vitro, 50,51 all of which again suggests that the cardiac hypertrophy of AVGN-treated mice is a physiologic benefit.
Cardiac and respiratory failure is the principle cause of morbidity and mortality among patients with Duchenne and many other muscular dystrophies. 52,53 It is also a substantial concern for cancer patients due to cardiac cachexia and because many chemotherapeutics can damage the myocardium. 54,55 The fact that AVGN7 enhances cardiac function, especially during a high-intensity test, is highly significant. Indeed, it not only suggests that AVGN7 is equally capable of improving both skeletal and cardiac function, but also that it could be used specifically to address some forms of cardiac dysfunction, possibly congestive heart failure. The cardiac remodeling that occurs with ischemic damage and a myocardial infarction helps to maintain cardiac output. Long term, however, the resulting fibrosis compromises cardiac function and can lead to heart failure. Thus, replacing this pathologic hypertrophy with physiologic hypertrophy, potentially with AVGN7 treatment, could significantly improve the clinical treatment of myocardial infarctions. It should be noted that skeletal muscle wasting also occurs with heart failure and is likely due to the endocrine actions of cardiac-derived myostatin, 28 which of course is antagonized by AVGN7.
Future proof-of-concept studies are nevertheless needed to determine definitively whether AVGN7 can in fact restore cardiac function in different models of heart failure. These should include an in-depth assessment of the underlying mechanisms that influence myocardial contractility at the organismal and cellular level and whether the mechanistic response to AVGN7 treatment truly mirrors that which occurs in myostatin-null hearts. Proof-of-concept studies are also needed with other animal disease models and will be complemented by ongoing dose-refinement and toxicology studies, as together, all of these studies will aid in the development of AVGN7 for a prioritized class of disease indications. It should be noted that the potential for abusing AVGN7 for performance enhancement in competitive athletics is well recognized. The authors' goals therefore also include developing tests for quantifying exposure to AVGN7 or any other AAV serotype that expresses Smad7.
Footnotes
Acknowledgments
This work was supported by a grant to B.D.R. from the Commercialization Gap Fund of the Washington Research Foundation.
Author Disclosure
B.D.R. is the founder and majority owner of AAVogen. He is also the co-discoverer and patent holder of AVGN7. The other authors were student or postdoctoral trainees or are currently faculty at Washington State University. They were not employed by AAVogen and have no financial interests in the company or its intellectual property. Thus, they declare no conflicts of interest.