
Abstract
Interleukin 35 (IL-35) is a relatively newly identified cytokine required for the regulatory and suppressive functions of regulatory T cells (Treg), playing an important role in the prevention of autoimmune diseases. This study used mesenchymal stem cells (MSCs) as the gene-delivery vehicles for IL-35 gene therapy and investigated their protective effects in Concanavalin A (Con A)–induced autoimmune hepatitis. Results showed that IL-35 gene modified MSCs (IL-35-MSCs) can specifically migrate to the injured liver tissues and significantly narrow the necrosis areas of injured livers. IL-35-MSCs prevented hepatocyte apoptosis by reducing the FASL expression by mononuclear cells. Although MSC treatment can alleviate liver injury to some extent, IL-35-MSCs showed a stronger protective effect, which means some novel mechanisms exist. The results show that IL-35-MSCs could decrease the level of interferon gamma secreted by liver mononuclear cells through the JAK1-STAT1/STAT4 signal pathway. In summary, this study thus demonstrates a novel and efficient treatment for Con A–induced fulminant hepatitis through negatively regulating the secretion of interferon gamma, thus providing a novel therapeutic approach for this devastating liver disease.
Introduction
R
In addition to the feature of immune regulation, mesenchymal stem cells (MSCs) can accumulate at the site of damage or inflammation and exhibit multipotency into a variety of mature cells. Because of these characteristics, MSC therapy has attracted considerable attention for the treatment of various diseases for many decades. 5 Their potential may be amplified by transforming MSCs with genes that could improve their therapeutic ability. Previous studies have demonstrated that MSC gene modification, including incorporation of exogenous genes such as IL-10, 6 hepatocyte growth factor, 7 IL-7, 8 and foxp3, 9 successfully improves the therapeutic ability of MSCs. In a previous study, the IL-35 gene was integrated into MSCs, and it was proven that the IL-35 gene-modified MSCs (IL-35-MSCs) can effectively inhibit the proliferation and function of CD4+ T cells in vitro. 10 Concanavalin A (Con A)–induced hepatitis is a well-documented murine model that closely resembles the pathology of human autoimmune hepatitis (AIH). 11 This study aimed to explore the protective role of IL-35-MSCs in Con A–induced liver injury and to provide some insights into novel and promising therapeutics for refractory liver diseases.
Materials and Methods
Animals
Male C57BL/6J mice were purchased from the Institute of Radiation Medicine Laboratory Animal Center (Tianjin, China). They were maintained in the animal facility under standard conditions, according to institutional guidelines. All animal protocols were reviewed and approved by the appropriate institutional review committee.
Reagents
Con A (type IV) was purchased from Sigma–Aldrich (St. Louis, MO).
IL-35 gene cloning and lentivirus production
Murine IL-35 cDNA was amplified from the pSecTag2A-IL35 plasmid (a gift from the Glasgow Biomedical Research Center), and subcloned into pCCS-Lv201 to generate the recombinant vector pCCS-IL35-Lv201. Lentivirus generation was performed by using GeneCopoeia 293Ta cells, according to the instructions of the lentivirus package kit (Lenti-Pac TM HIV; GeneCopoeia, Rockville, MD). Cell culture supernatants were collected and concentrated with the Lenti-Pac concentration kit (GeneCopoeia).
Ad-MSC culture and transfection
MSCs from mouse adipose tissues grew to sub-confluence after 7 days and showed a fibroblast-like morphology. Confluent third-passage cells were used for lentivirus transfection. A multiplicity of infection (MOI) of 10 was selected, and 5 μg/mL of polybrene (Sigma–Aldrich) was added. Green fluorescent protein (GFP)-positive transduced Ad-MSCs were sorted by a BD FACS Aria II instrument (BD Biosciences, San Jose, CA) for further experiments.
T cell–mediated hepatitis model and cell transplantation
To induce hepatitis, C57BL/6J mice (6–8 weeks old) were injected intravenously with Con A (15 mg/kg body weight). For the survival study, the dose of Con A was increased to 20 mg/kg body weight. Sex- and age-matched C57BL6/J mice were divided into three groups and intravenously injected with IL-35-MSCs, MSCs, or phosphate-buffered saline (PBS). Two hours later, Con A was administrated intravenously. After cell transplantation, blood samples were collected, and the liver tissues were taken and prepared for further analysis.
Assay for serum transaminase activity
Mouse serum samples were obtained at different time points post Con A injection. Serum alanine aminotransferase (ALT) activities were measured using a commercially available test kit (Rong Sheng Biotech, Shanghai, China) based on methods recommended by the International Federation of Clinical Chemistry.
Histological examination
Liver tissues were harvested after treatment with Con A, fixed in 10% formalin, and embedded in paraffin. Five-micron sections were affixed to slides, and stained with hematoxylin and eosin (H&E). Images ( × 100) were acquired on an Olympus DX71 (Olympus Imaging America, Center Valley, PA). The liver damage extent was quantified by the necrosis area as a percentage of total area using Image-Pro Plus v6.0 software (Media Cybernetics, Silver Spring, MD). Four different fields were randomly chosen in every slide, and the average necrosis percentage was calculated.
Immunohistochemistry
To quantitate the apoptosis level of hepatocytes, sections were stained with cleaved caspase 3 antibodies (Abcam, Cambridge, MA). The primary antibody was at a dilution of 1:100. Negative control sections, stained only with secondary antibodies without primary antibodies, were performed to assess the nonspecific staining. The specimens were stained according to the instructions of the strept avidin–biotin complex (SABC) kit. Stained sections were photographed using an Olympus DX71 (Olympus Imaging America). At high magnification ( × 400), 10 non-overlapping views were randomly observed, and the protein expression index (PEI) was calculated: PEI % = caspase-3 positive cell numbers/total observed cell numbers.
In vivo tracking detection of IL-35-MSCs
Liver tissues were collected and fixed by OCT for frozen section to detect the GFP distribution. Five-micron sections were affixed to slides. Images ( × 100) were acquired on an Olympus DX51 (Olympus Imaging America).
Preparation of liver mononuclear cells
Mouse livers were removed and pressed through a 200-gauge stainless steel mesh. The liver cell suspension was collected, suspended in PBS, and centrifuged at 50 g for 5 min. Supernatants containing mononuclear cells (MNCs) were collected and resuspended in 40% Percoll (GE Healthcare, Chicago, IL). The cell suspension was gently overlaid onto 70% Percoll and centrifuged for 30 min at 1,260 g. MNCs were collected from the interphase and washed twice in PBS.
Real-time polymerase chain reaction for gene transcription
Total RNA was extracted from liver MNCs by using TRIzol Reagent (Invitrogen, Carlsbad, CA) and reverse-transcribed with a Quantscript RT Kit (Tiangen, Beijing, China). mRNA expression was quantified by SYBR Premix Hotmaster Taq (Tiangen), and RPLP0 12 was used as an internal normalizing gene. The primer sequences used were as follows: RPLP0 forward: 5′-GAAACTGCTGCCTCACATCCG-3′; RPLP0 reverse: 5′-CTGGCACAGTGACCTCACACG-3′; interferon gamma (IFN-γ) forward: 5′-ATGAACGCTACACACTGCATC-3′; IFN-γ reverse: 5′-CCATCCTTTTGCCAGTTCCTC-3′; IL-17 forward: 5′-TGAAGGCAGCAGCGATCA-3′; IL-17 reverse: 5′-GGAAGTCCTTGGCCTCAGTGT-3′; FASL forward: 5′-CAGCTTCAGATGCAAGTGAGTGG-3′; FASL reverse: 5′-CAAGGACAGAACTCTGACGCTGAC-3′.
Western blot
After 24 h of treatment, all mice were killed, liver tissues were homogenized, and the total protein was extracted. Then 40 μg of protein per sample was subjected to 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis, and the proteins were transferred onto polyvinylidene fluoride membranes (Millipore, Billerica, MA). After overnight incubation at 4°C with anti-phospho-STAT1 (Tyr701, D4A7), anti-STAT1 (D1K9Y), anti-phospho-STAT4 (Tyr693,D2E4), anti-STAT4 (C46B10), anti-phospho-JAK1 (Tyr1022/1023), anti-JAK1 (6G4), anti-phospho-JAK2 (Tyr1007/1008,C80C3), anti-JAK2 (D2E12) antibodies (all Cell Signaling Technology, Beverly, MA), and anti-IFN-γ (R&D, Minneapolis, MN), the membranes with blotted proteins were incubated with horseradish peroxidase–conjugated goat anti-rabbit secondary antibodies (Millipore; 1:3,000 dilution) or rabbit anti-rat secondary antibodies (Boster, Wuhan, China; 1:1,000 dilution) for 2 h at room temperature. The targeted proteins were detected and quantified using the G:BOX F3 gel doc system. The membranes were stripped and reblotted with β-actin antibody to verify the equal loading of protein in each lane.
Statistics
GraphPad Prism v6.01 (GraphPad Software, Inc., La Jolla, CA) was used to perform all statistical analyses. Data were expressed as mean ± standard deviation, and one-way analysis of variance was used for group comparisons. Throughout the text, figures, and legends, statistical significance is denoted as *p < 0.05 and **p < 0.01.
Results
IL-35-MSCs play a protective role in Con A–induced fulminant hepatitis
To define the role of IL-35-MSCs in Con A–induced hepatitis, sex- and age-matched C57BL/6J mice were transplanted with IL-35-MSCs, MSCs, or PBS, and 2 h later, a high dose of Con A (20 mg/kg body weight) was administrated by intravenous injection. All the mice transplanted with IL-35-MSCs survived throughout the whole period of observation, while the PBS pretreated mice died within 24 h. The survival rate of MSC-transplanted mice was 40% (Fig. 1). In order to determine the protective role of IL-35-MSCs in this model further, different doses of Con A were administered into different treated mice. A lower dose of Con A (15 mg/kg body weight) was found to be the best to demonstrate significant differences among these three groups (Fig. 2A). Thereafter, all of the following experiments were performed using this lower dose. The serum ALT level peaked at 24 h after treatment with Con A (Fig. 2A and B), and so further mechanisms researched mainly focused on this point.
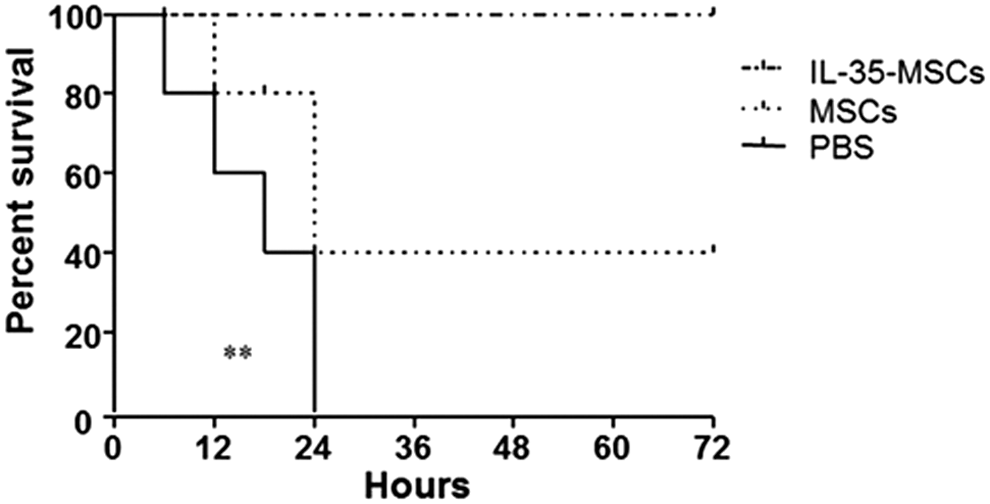
Interleukin (IL)-35 mesenchymal stem cells (MSCs) exerted a protective role in Concanavalin A (Con A)–induced hepatitis. Sex- and age-matched C57BL6/J mice were divided into three groups (n = 15 in each group) and were intravenously injected with IL-35-MSCs, MSCs, or phosphate-buffered saline (PBS). Two hours later, Con A (20 mg/kg body weight) was administrated by intravenous injection, and the rates of death were observed every 30 min and recorded. One representative experiment out of three repeated experiments is shown (*p < 0.05).
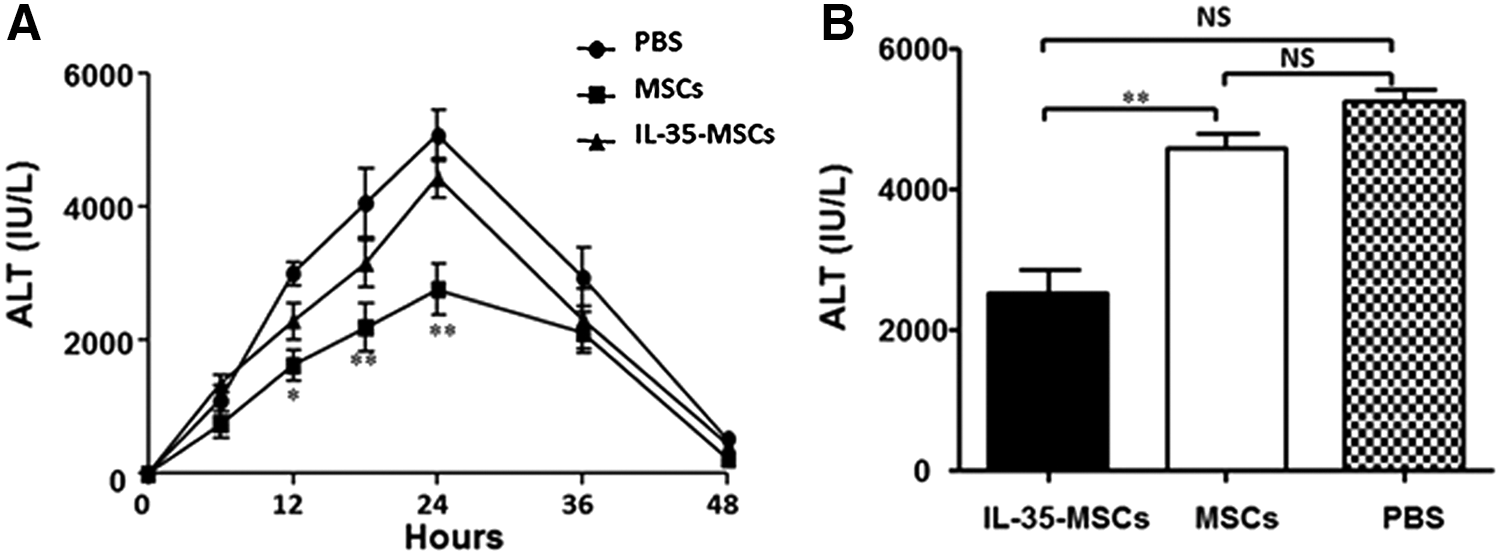
Transplantation of IL-35-MSCs decrease the alanine aminotransferase (ALT) level in Con A–induced hepatitis. Sex- and age-matched C57BL6/J mice were divided into three groups (n = 6 in each group) and were intravenously injected with IL-35-MSCs, MSCs, or PBS. Two hours later, Con A (15 mg/kg body weight) was administrated by intravenous injection.
Transplantation of IL-35-MSCs decreases hepatocyte necrosis and apoptosis
To define the role of IL-35-MSCs in Con A–induced hepatitis further, sex- and age-matched C57BL6/J mice were intravenously injected with IL-35-MSCs, MSCs, or PBS. Two hours later, Con A (15 mg/kg body weight) was administrated by intravenous injection. Liver necrosis was measured by H&E staining, and the cleaved caspase3 protein positive cell numbers were calculated (Fig. 3A and B). Although both IL-35-MSCs and MSCs narrowed the necrosis areas and decreased the caspase3 positive cell numbers significantly compared to the PBS control, pretreatment with IL-35-MSCs showed a better therapeutic effect than pure MSCs did (Fig. 3C and D). Furthermore, the changes of FASL mRNA expression levels of liver MNCs were detected, and the results showed that treatment with IL-35-MSCs significantly lowered the FASL expression level (Fig. 3E).

Transplantation of IL-35-MSCs prevents liver necrosis and hepatocyte apoptosis in Con A–induced liver injury. Sex- and age-matched C57BL6/J mice were divided into three groups (n = 6 in each group) and were intravenously injected with IL-35-MSCs, MSCs, or PBS. Two hours later, Con A (15 mg/kg body weight) was administrated by intravenous injection, and all the mice were euthanized 24 h after Con A treatment. Liver tissues were fixed by formalin.
Tracing of transplanted IL-35-MSCs
To investigate whether IL-35-MSCs transplanted to Con A injured the liver through the intravenous injection, liver tissues were fixed by OCT for frozen section to detect the GFP distribution. As shown in Fig. 4, GFP-positive cells were easily detected in the livers of the IL-35-MSC pretreated group rather than the MSC and PBS groups by means of fluorescence microscopy. Although the lung, spleen, and kidney were also examined in the IL-35-MSC pretreated group, there were no GFP-positive cells detected in these organs (data not shown).
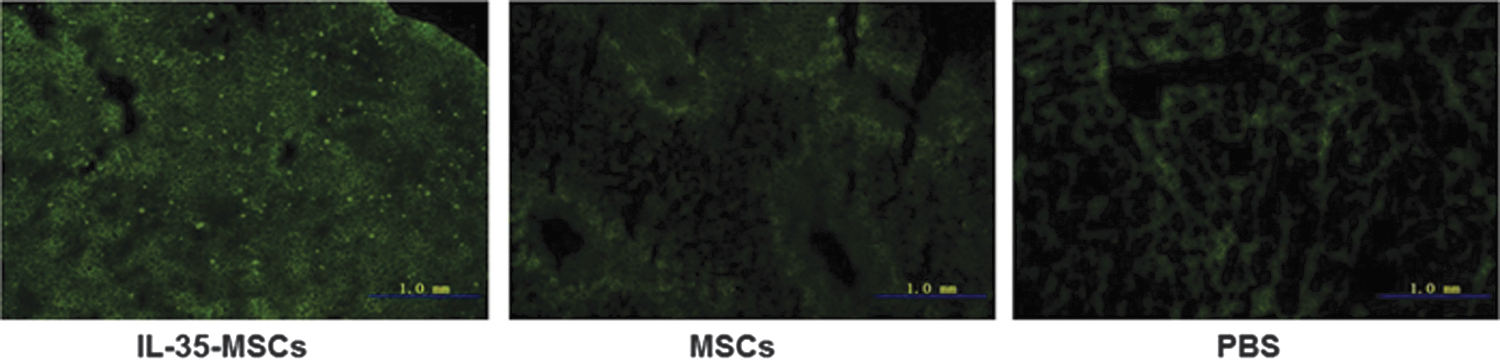
Tracing of transplanted IL-35-MSCs. Sex- and age-matched C57BL6/J mice were divided into three groups (n = 6 in each group) and received different treatment, as described above. Two hours later, Con A (15 mg/kg body weight) was administrated by intravenous injection. All the mice were euthanized 24 h after Con A treatment. Liver tissues were fixed by OCT for frozen section to detect the green fluorescent protein distribution.
IFN-γ was a critical cytokine for protection against Con A–induced hepatitis played by IL-35-MSCs
To determine whether treatment with IL-35-MSCs could affect cytokine profiles, the cytokine expression level of MNCs was analyzed and compared among different groups. The IFN-γ expression level decreased significantly in the IL-35-MSC transplanted group compared to the pure MSC- and PBS-treated control groups (Fig. 5A). The IL-17 expression level was significantly lowered in both IL-35-MSCs and MSCs compared to the PBS-treated group. However, there was no significant difference in the IL-17 expression between the IL-35-MSC and MSC groups (Fig. 5B). The expression of other cytokines involved in the Con A–involved liver injury were also detected, including the tumor necrosis factor (TNF)-α, IL-4, IL-2, and IL-12. Unfortunately, no significant difference was found (data not shown). Furthermore, the IL-17 and IFN-γ contents in the liver tissue was examined by Western blot analysis, and the variation tendency was consistent with the liver MNCs (Fig. 5C and D).
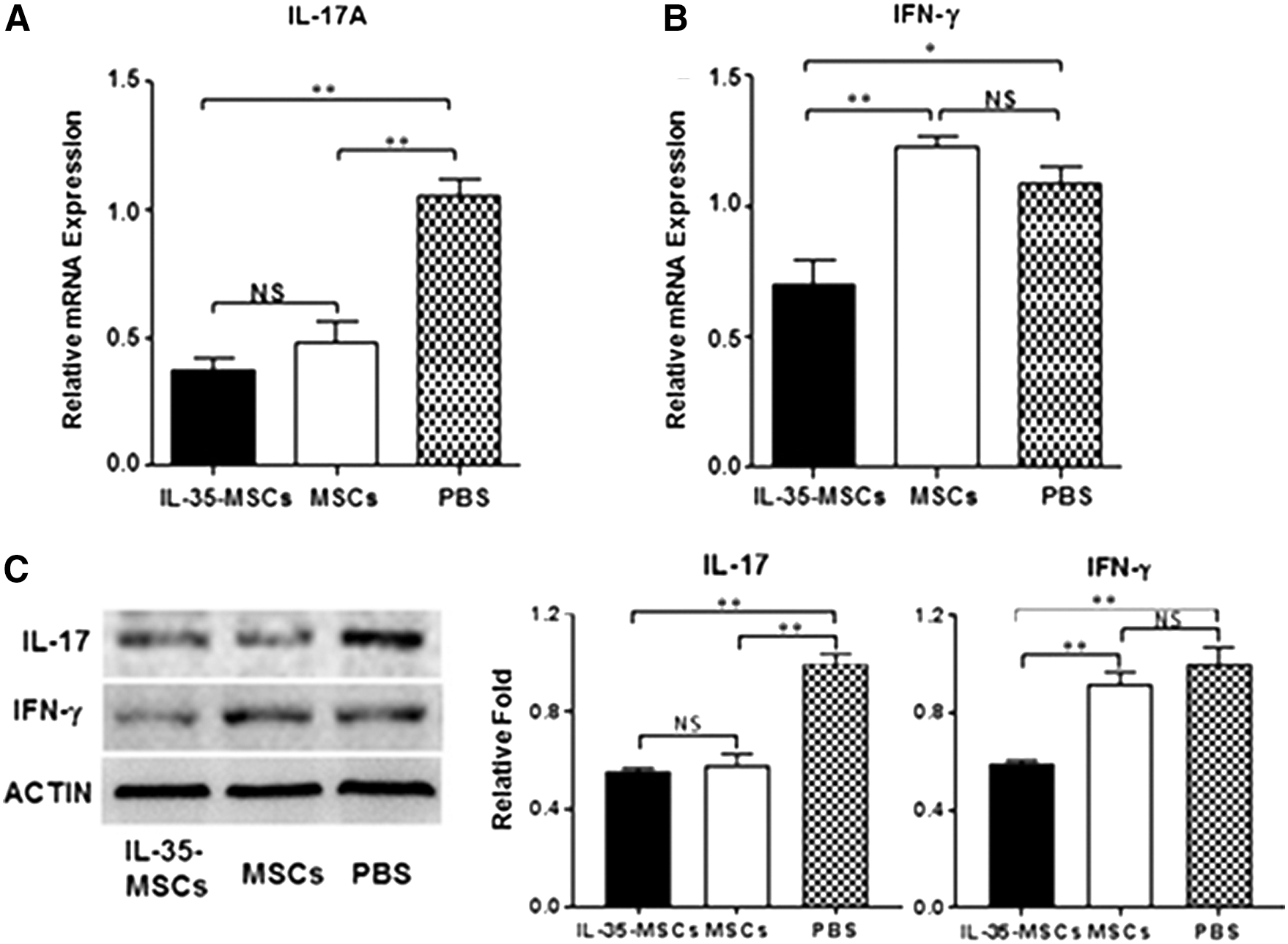
IL-35-MSCs regulated the expression of IL-17 and interferon (IFN)-γ. Sex- and age-matched C57BL6/J mice were grouped and treated as described above. Liver MNCs were prepared, and total RNA was extracted using TRIzol Reagent.
IL-35-MSCs regulated the expression of inflammation factors through the JAK1-STAT1/4 pathway
To investigate the mechanism of the cell therapies, the proteins involved in the IL-35 and IL-35R signal pathway were detected. The results showed that the protein expression of JAK1, STAT1, and STAT4 increased significantly in the group treated with IL-35-MSCs compared to the pure MSCs and the PBS control groups. Furthermore, the protein expression changes were amplified significantly after phosphorylation. Meanwhile, there was no significant difference in the expression of JAK2 and no obvious detection of pJAK2 among the three groups (Fig. 6).

IL-35-MSCs play effects through the JAK1-STAT signal pathway. The mice were grouped and treated as above, and the liver tissues were collected. Total protein was extracted from the harvested liver tissues and was analyzed to determine the levels of IL-35-related transcription factors using Western blot analysis.
Discussion
Con A–induced fulminant hepatitis is a well-known animal model for studying the pathophysiological mechanisms of AIH, a chronic inflammatory liver disease with significant mortality worldwide and without effective therapeutic approaches. IL-35, a new member of the IL-12 family, is a heterodimeric cytokine composed of the p35 subunit of IL-12 and Ebi3 subunit of IL-27 and is preferentially secreted by Tregs. 13 IL-35 is highly expressed in non-stimulated mouse Treg cells and stimulated human Treg cells, but it is not detected in non-stimulated human Treg cells. 14 IL-35 also can be produced by other cell types such as the dendritic cell and γδ T cells. 15 In contrast to the other IL-12 family members (IL-12 and IL-23), IL-35 appears to have a strictly regulatory function. The primary physiological role of IL-35 is to regulate T-cell homeostasis, such as inhibiting T helper (Th) 2 and Th17 polarization. 16 Furthermore, IL-35 exerts anti-inflammatory effects in autoimmune diseases such as EAE, EAU, CIA, inflammatory bowel disease, asthma, and so on. 17 –19 IL-35 also induces the conversion of conventional T cells into a suppressive, IL-35-producing CD4+Foxp3−-induced regulatory T-cell population (iTr35 cells). 20 However, the physiological impact of IL-35 on Con A–induced hepatitis is unknown. The idea of introducing genes into cells has been practiced for many years. MSCs are good candidates as gene-delivery vehicles for targeted therapy, as these cells possess the ability to home to sites of injury and damage. 21 During the past decades, remarkable progress has been made in the area of gene-engineered MSCs-based therapy. 22 –25 The main purpose of this study was to investigate the effect of IL-35 gene programmed MSCs in preventing Con A–induced liver injury.
It has been proven that IL-35-MSCs can effectively inhibit the proliferation and function of CD4 + T cells in vitro. 10 The present study observed that the transplanted IL-35-MSCs could specifically migrate to the damaged liver tissues, significantly prolong the survival rate, and ameliorate the extent of the liver injury compared to the MSCs- and PBS-treated groups. Similarly, MSCs transplantation could narrow the liver necrosis area, decrease the expression level of active caspase3, and reduce the FASL mRNA level of MNCs, consist with previous studies showing that MSCs can attenuate liver injury. 26 IL-35-MSCs exhibit a stronger protective effect through a combination of their individual characteristics.
Activated T cells have been defined to play a critical role in Con A–induced liver damage through producing cytokines, such as IFN-γ, IL-4, IL-2, IL-6, IL-10, IL-12, and TNF-α. 27 This study analyzed the mRNA and protein expression level of several principal inflammatory cytokines in the hepatic MNCs. The results show that IFN-γ expression was significantly decreased in the IL-35-MSCs-transplanted group compared to the MSCs and PBS groups. While the IL-17 expression level of MNCs was significantly lowered in both the IL-35-MSCs and MSCs treatment groups, there was no significance between them. These results suggest that MSCs could alleviate liver injury by reducing IL-17 secretion of liver MNCs but not IFN-γ. IL-35-MSCs could exert stronger protection through regulating the secretion of both IL-17 and IFN-γ, which might be the result of a combined action.
Since IL-35 appears to utilize receptor chains, IL-12Rβ2 or gp130 homodimers, as well as a heterodimer, the complete IL-12Rβ2:gp130 receptor was required for maximal suppression, IL-35 induction and iTr35 conversion. IL-35 signaled through a STAT1:STAT4 heterodimer, promoting a distinct mode of Il12a and Ebi3 promoter binding and gene activation. 28 In order to determine the potential involvement of transcription factors in liver injury prevention, the expression levels of the transcription factors that might be involved in the IL-35/IL35R signal pathway, including JAK1, pJAK1, JAK2, pJAK2, STAT1, pSTAT1, STAT4, and pSTAT4, were all determined by Western blot. The results show that both JAK1 and phosphorylated JAK1 expression were increased significantly in IL-35-MSCs-treated group, but there was no change in the JAK2 expression and no phosphorylated JAK2 was detected, which means that JAK1 mediated the signal transduction of IL-35. Furthermore, both the dephosphorylated and phosphorylated forms of STAT1 and STAT4 expression level were elevated significantly after IL-35-MSCs transplantation compared to the other two groups. Meanwhile, the protein levels of dephosphorylated and phosphorylated forms of JAK1, STAT1, and STAT4 in the MSCs-transplanted group were also higher than in the PBS control, though their increased amplitude was smaller compared to IL-35-MSCs group. The results prove that both IL-35-MSCs and MSCs played a protective role in the Con A induced fulminant hepatitis, but IL-35-MSCs exerted stronger therapeutic effects than MSCs.
In conclusion, the transplanted IL-35-MSCs exerted obvious therapeutic effects on Con A–induced liver injury than MSCs, though they shared some similar functions. IL-35-MSCs prevented hepatocyte apoptosis by reducing the FASL expression by MNCs and decreased the IFN-γ expression level through the JAK1-STAT1/STAT4 signal pathway.
Footnotes
Acknowledgments
This work was supported by the National Natural Science Foundation of China for Young Scholars (Grant No. 31400764) and Tianjin City High School Science and Technology Fund Planning Project (Grant No. 20130113).
Author Disclosure
The authors have no financial conflicts of interest.