
Abstract
Postprandial glucose-induced insulin secretion from the islets of Langerhans is facilitated by glucagon-like peptide-1 (GLP-1)—a metabolic hormone with insulinotropic properties. Among the variety of effects it mediates, GLP-1 induces delta cell secretion of somatostatin, inhibits alpha cell release of glucagon, reduces gastric emptying, and slows food intake. These events collectively contribute to weight loss over time. During type 2 diabetes (T2DM), however, the incretin response to glucose is reduced and accompanied by a moderate reduction in GLP-1 secretion. To compensate for the reduced incretin effect, a human immunodeficiency virus–based lentiviral vector was generated to deliver DNA encoding human GLP-1 (LentiGLP-1), and the anti-diabetic efficacy of LentiGLP-1 was tested in a high-fat diet/
Introduction
T
Pancreatic beta cell mass is mainly determined by islet cell replication, apoptosis, and new islet formation from exocrine pancreatic ducts, and agents that interfere with beta cell formation or new islet generation can decrease beta cell mass. 6 Unfortunately, disease progression in obese diabetic individuals cannot be reversed by medications that only induce insulin sensitivity and/or weight loss. Because of the diabetes pandemic seen in recent years, new therapeutic strategies are needed to prevent or reverse disease progression. 7 Agents that increase beta cell mass are considered useful in managing T2DM, 8 where the ideal beta cell preserving agent should reduce insulin resistance, increase glucose-induced insulin secretion, promote beta cell replication and/or islet neogenesis, and protect islet cells from apoptosis.
Glucagon-like peptide-1 (GLP-1) can increase beta cell mass in experimental animal models of diabetes. 9 –11 The peptide is initially synthesized as part of the larger prohormone proglucagon expressed in the α-cells of the pancreas, L-cells of the intestine, and neurons located in the caudal brainstem and hypothalamus. In addition to GLP-1, glucagon, GLP-2, glicentin, and oxyntomodulin are also encoded within proglucagon fragment. Intriguingly, GLP-1 and glucagon are produced as a result of the differential post-translational processing of preproglucagon fragment in the intestine and pancreas, respectively. 12 Moreover, GLP-1 is an incretin hormone capable of stimulating glucose-induced insulin secretion from pancreatic beta cells. 13,14 It can activate somatostatin release from pancreatic delta cells, which inhibits glucagon secretion from alpha cells. 15 Stimulation of beta cell proliferation, differentiation, and inhibition of islet cell apoptosis are among the many beneficial effects of GLP-1 relevant to diabetes therapy. 16 GLP-1 analogs (e.g., exenatide and liraglutide) are currently used in the treatment of T2DM. 17,18 However, incretin-based treatments manifest some concerning side-effects. 19 Exenatide and liraglutide have been suspected to cause pancreatitis and pancreas cancer in addition to their observed gastrointestinal side effects. 20 –22 Apart from these concerns, daily injections of exenatide (twice) and liraglutide (once) are needed to observe any therapeutic benefit. Since an insufficient incretin response to meals is one of the fundamental defects in T2DM patients, 23,24 restoring the incretin response by gene therapy represents an attractive way of treating the disease. 25
To provide constant GLP-1 synthesis and secretion in vivo, a lentiviral vector carrying the GLP-1 gene (LentiGLP-1) was constructed, and its therapeutic efficacy was tested in obese diabetic rats. LentiGLP-1 administration significantly reduced blood glucose and concurrently improved insulin sensitivity and glucose tolerance. Importantly, normoglycemia correlated with an increase in blood GLP-1 and pancreatic beta cell regeneration following LentiGLP-1 delivery. The data obtained suggest the clinical potential of GLP-1 gene transfer therapy for the treatment of T2DM. The implications of these findings will be discussed.
Materials and Methods
Animal handling and care
Male Sprague Dawley rats (SDW) were obtained at 4 weeks of age from the Animal Care Laboratory of Akdeniz University Hospitals and handled in accordance with the regulations of the Institutional Animal Care and Use Committee of the Akdeniz University School of Medicine. Rats were housed in a humidity and temperature-controlled room under a 12 h light/12 h dark cycle on sterilized bedding, with four rats per cage. Water and food were provided ad libitum. Rats were fed a standard (SD) or high-fat diet (HFD) where 60% of the calories came from fat. After being on the respective diets for 3 months, rats were given an intraperitoneal injection of streptozotocin (STZ; 40 mg/kg; Sigma–Aldrich, St. Louis, MO) to induce diabetes. Prior to injection, STZ was freshly dissolved in citrate buffer (0.01 M, pH 4.5) and kept on ice prior to use. Cages were changed twice a week after STZ injection to provide dry bedding for polyuric animals. Age-matched nondiabetic rats were used as controls. All rats remained on their respective diets for the duration of the study. Blood samples were collected via intracardiac puncture, and serum was obtained using a Serum Separator Vaccutainer (Vacuaette, cat. no. 454343; Greiner Bio-One, Kremsmünster, Austria) by centrifugation for 15 min at 1750 × g. Serum samples were stored at −20°C until use.
Construction of human GLP-1 encoding lentiviral vectors
The human GLP-1 coding sequence (193 bases in length), including insulin signal peptide and a furin cleavage site, was synthesized by Integrated DNA Technology Company (IDT, Coralville, IA) as a synthetic double-strand DNA molecule. The cloning strategy required addition of CACC nucleotides to the 5′ end of GLP-1 coding sequence. Directional TOPO cloning was used to generate a human GLP-1 coding entry vector using pENTR/D-TOPO Cloning Kit (K2400-20; Invitrogen, Carlsbad, CA). Subsequently, a GLP-1 encoding lentiviral expression clone (pLentiGLP-1) was generated using Gateway Technology. For this purpose, the GLP-1 transgene was cloned into the Gateway destination vector (pLEnti6.3/V5-DEST) using Virapower Hiperform Lentiviral Gateway Expression Kit (K5330-00; Invitrogen). Restriction enzyme analysis and DNA sequencing using a Big Dye Terminator v3.1 Cycle Sequencing Kit (cat. no. 4336917; Applied Biosystems, Foster City, CA) were performed to confirm the orientation and the DNA sequence.
Lentivirus production and titration
The following plasmids were used to obtain the necessary transacting factors for packaging: pMDLg/pRRE (HIV-1 pGag-Pol 12251), Rev-Plazmid (pRSV-Rev 12253), and pMD2.G (pVSV-G 12259; Addgene). The LV-RFP plasmid (Plasmid 26001) was used for the optimization of lentivirus production. In addition, a β-galactosidase encoding LentiLacZ vector was generated using LV-Lac plasmid (Plasmid 12108). For lentivirus production, 293T cells were cultured in roller bottles (8 × 107 cells/bottle) with 1,700 cm2 of growth area (Cell Master™ roller bottles; cat. no. 681 062; Greiner Bio-One GMbH, Frickenhausen, Germany) containing 10% fetal bovine serum (FBS), 1% penicillin-streptomycin (Pen + Strep) containing Dulbecco's modified Eagle's medium (DMEM; 4.5 g/L of D-glucose, 4 mM of L-glutamine, sodium pyruvate). Bottles were rotated at 0.3 rpm for 24 h and then the speed was raised to 1 rpm for the following 12–16 h in a HERAcell 240i CO2 incubator (Thermo Fisher Scientific, Waltham, MA). Cell culture medium was replaced with transfection medium (IMDM; cat. no. I7633; Sigma–Aldrich) containing 10% FBS, 1% Pen + Strep, and 25 μM of chloroquine (cat. no. C6628; Sigma–Aldrich) 1 h prior to transfection. Roller bottles were incubated in a 5% CO2 incubator at 37°C. For transient transfection, 176 μg of pGag-Pol, 68 μg of pRSV-Rev, 95 μg of pVSV-G, and 270 μg of pLentiGLP-1 plasmids were mixed in a conical tube. Next, 10.5 mL of 0.1 × TE:dH2O (2:1 v/v) and 1.5 mL of 2 M CaCl2 was added to the mixture. The precipitate was formed by adding the DNA:CaCl2 mixture drop wise into 12 mL of 2 × HEPES buffered saline (pH 7.0). After 5 min of incubation at room temperature, the mixture was added to the cell cultures. Subsequent to 6–8 h of incubation of the roller bottles at 0.3 rpm, the medium was replaced with harvest media (OPTI-MEM; 22600134; Gibco, Gaithersburg, MD) containing 10% FBS, 1% Pen + Strep, and then incubated at 37°C in a 5% CO2 incubator (1 rpm) for 2.5 days to allow lentiviral vector production. Viral supernatants were then cleared by low-speed centrifugation (2,000 g for 15 min) and filtered through 0.45 μm cellulose acetate filters. Unconcentrated viral supernatants were added to ultracentrifuge tubes (pollyallomer ultracentrifuge tubes; cat. no. 326823; Beckman Coulter, Brea, CA), and a sucrose cushion was created by layering 30 mL of viral supernatants onto 5 mL of 10% sucrose (in Hank's balanced salt solution [HBSS]). Tubes were ultracentrifuged at ∼82000 × g for 2.5 h with a Beckman SW28 rotor. Supernatants were poured off, and inverted tubes were rested on paper towels to allow any remaining liquid to drain. Virus pellets were re-suspended in 100 μL of HBSS and incubated at 4°C overnight. Viruses were collected the following day by pipetting, and then they were kept at −80°C prior to titration analysis. Physical titration of lentiviral vectors was performed using a QuickTiterTM HIV Lentiviral Quantitation Kit (HIV p24 ELISA; cat. no. VPK-108-H; Cell Biolabs, Inc., San Diego, CA). Functional titration of lentiviral vectors was determined by real-time polymerase chain reaction (PCR) using a Quantitect SYBR Green PCR Kit (cat. no. 204143; Qiagen, Valencia, CA) in an ABI 7500 Fast Real-Time PCR instrument. Woodchuck posttranscriptional regulatory element primers (Fwd: 5′-CCGTTGTCAGGCAACGTG-3′; Rev: 5′-AGCTGACAGGTGGTGGCAAT-3′) were used to determine viral genome amount, and albumin primers (Fwd: 5′-GCTGTCATCTCTTGTGGGCTGT-3′; Rev: 5′-ACTCATGGGAGCTGCTGGTTC-3′) were used as an internal control for real-time PCR. A standard curve was generated by using serial dilutions of albumin plasmid (Addgene). Relative levels of integrated vector copy numbers were determined using the 2–ΔΔCT method for each group. Data are given as normalized integrated copy numbers relative to the control rat tissues.
Blood glucose measurement
Blood glucose levels were measured using an AccuCheck Compact Glucometer (Roche Diagnostics, Indianapolis, IN). Rats exhibiting blood glucose levels >250 mg/dL were considered diabetic. All blood glucose measurements were performed early in the afternoon to minimize any variables due to feeding pattern. Insulin sensitivity and glucose tolerance tests were performed in accordance with the protocols of Animal Models of Diabetic Complications Consortium (AMDCC Protocols).
Histological analysis
Rats were sacrificed 28–30 days after vector injection, and pancreatic tissue was fixed with formalin by intraductal perfusion. Pancreas and intraabdominal tissues (the liver, spleen, and kidney) were embedded in paraffin cassettes. Hematoxylin and eosin (H&E) staining and immunostaining with rabbit polyclonal anti-insulin antiserum (cat. no. ab63820; Abcam, Cambridge, United Kingdom), rabbit polyclonal anti-glucagon antiserum (cat. no. ab18461; Abcam), or rabbit polyclonal anti-GLP-1 (cat. no. ab22625; Abcam) were performed on pancreatic sections. Subsequently, beta cell area was calculated by comparing the insulin-positive area to the total pancreas area using Image J software (National Institutes of Health, Bethesda, MD;
GLP-1 enzyme-linked immunosorbent assay
GLP-1 measurements from cell culture supernatants and blood serum samples were carried out using a GLP-1 (Active) enzyme-linked immunosorbent assay (ELISA) kit (cat. no. EGLP-35K; Millipore, Darmstadt, Germany). For in vitro analysis, 5 × 104 HT1080 cells were seeded into each well of a 24-well plate. Cells were transduced with the increasing doses of lentiviruses in the presence of polybrene. Supernatants were collected 72 h after transduction, and dipeptidyl peptidase IV inhibitor was added to the samples immediately after collection. Blood for serum was obtained from sacrificed rats via intracardiac puncture. All samples were stored at −20°C prior to ELISA.
Cyclic adenosine monophosphate measurements
MIN6 cells were transduced with GLP-1 encoding lentiviral vectors at increasing doses (5, 25, or 100 multiplicities of infection [MOI]). Following transduction, cyclic adenosine monophosphate (cAMP) levels were measured with a cAMP Direct Immunoassay Kit (ab65355) according to the manufacturer's protocol. LentiLacZ transduced cells were used as a control.
Glucose-stimulated insulin secretion test
MIN6 cells (provided by Prof. Jun-Ichi Miyazaki, Osaka University, Osaka, Japan) were added to 96-well flat-bottom plates (105 cells/well). The cells were transduced the next day with LentiGLP-1 (5, 25, or 100 MOI) or LentiLacZ (100 MOI) vectors in the presence of polybrene. One day after transduction, cell culture media was replaced with fresh DMEM, and cells were further incubated for 3 days at 37°C in a 5% CO2 incubator. Supernatants were collected, and the cells were washed with Kreb's Ringer Buffer (KRB) with no glucose. KRB containing different glucose concentrations (no glucose, 2.8 mM of glucose, and 25 mM of glucose) were then added to the wells to analyze the effect of glucose on insulin secretion. Supernatants were collected 1 h later, and an insulin ELISA was performed using an ultra-sensitive mouse/rat insulin ELISA kit (cat. no. 90080; Crystal Chem, Inc., Elk Grove Village, IL).
Analysis of blood lipid profile
Triglyceride levels in blood samples were analyzed by enzymatic/cholorimetric method using EnzyChrom Triglyceride Assay Kit (cat. no. ETGA-200; BioAssay Systems, Hayward, CA). Cholesterol levels were determined using a cholesterol/cholesteryl ester quantitation kit (#ab65359; Abcam).
Statistical analysis
Data are presented as the mean ± standard error of the mean. Statistical analysis for differences between groups was analyzed using GraphPad Prism v5 (GraphPad Software, Inc., La Jolla, CA) software. Differences were considered significant when p < 0.05.
Results
Generation of diet-induced obesity SDW rat model of T2DM
SDW rats were fed either a HFD where 60% of the calories came from fat or SD. Weight gain and blood glucose levels of animals were monitored periodically for the following 27 weeks (Fig. 1A). As little as 5 weeks of HFD feeding was sufficient to generate a significant increase in body weight in the HFD rats compared to the SD rats. Laparoscopic surgical examination conducted 2 months after the feeding also confirmed abdominal fat deposition in HFD rats (Fig. 1B). Surprisingly, blood glucose levels between the two groups were not statistically different (data not shown). Since insulin resistance and glucose intolerance are considered prediabetic findings in obese patients, HFD rats were tested for insulin sensitivity and glucose intolerance after 2 months of feeding. HFD rats clearly displayed insulin resistance (Fig. 1C) and glucose intolerance (Fig. 1D) compared to SD rats. These results led to the conclusion that a HFD induces obesity but not hyperglycemia or diabetes (>250 mg/dL) in SDW rats.

Next, the extent to which different doses of streptozotocin (STZ; 30 mg/kg, 40 mg/kg, and 50 mg/kg) could induce diabetes in obese SDW rats was tested. Intraperitoneal (i.p.) administration of 40 or 50 mg/kg of STZ successfully induced hyperglycemia in obese SDW rats, while 30 mg/kg of STZ was not sufficient to do so (Fig. 2A). Considering the high mortality rate seen after giving the 50 mg/kg dose of STZ, the decision was made to use 40 mg/kg of STZ to induce diabetes, with an acceptable survival rate in obese SDW rats (Fig. 2B). Collectively, these data suggested HFD feeding of SDW rats following STZ administration is a suitable T2DM animal model for subsequent experiments.
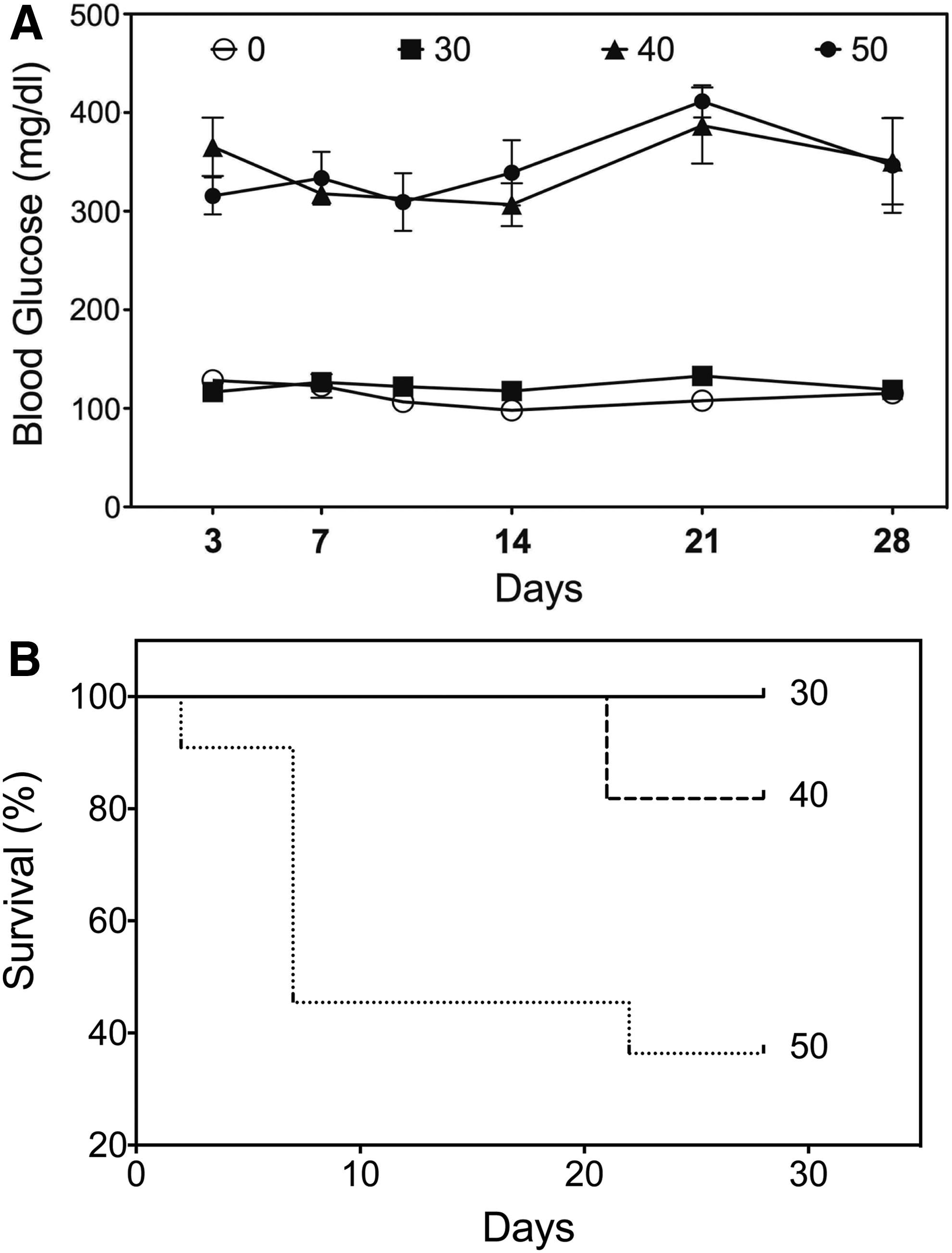
Streptozotocin (STZ) dose testing and induction of diabetes. Obese SDW rats injected intraperitoneally (i.p.) with 0 (n = 14), 30 (n = 10), 40 (n = 11), and 50 (n = 13) mg/kg STZ to induce hyperglycemia. Blood glucose levels
Construction and functional analysis of lentivirus vector expressing GLP-1
The prohormone proglucagon consists of 180 aa that when fragmented become glucagon, GLP-1, GLP-2, glicentin, and oxyntomodulin. 26 Since the use of the preproglucagon transgene in a vector would generate all of these bioactive fragments, gene therapy studies involving GLP-1 are restricted to GLP-17–37. For the production of the GLP-1 encoding lentivirus, an oligonucleotide containing the sequence for a translation start codon (methionine) upstream of the GLP-17–37 coding sequence was synthesized. In addition, the sequence for a secretory signal peptide was included to target GLP-1 to the constitutive secretory pathway (CSP) to provide post-translational processing by a signal peptidase. Lastly, a furin peptidase recognition site (RGRR) was included in the GLP-1 cDNA to permit the removal of the preceding amino acids by furin endopeptidases to generate the active form of the peptide capable of receptor binding before secretion. The Gateway Hiperform lentiviral expression system was used to construct a lentivirus vector carrying the GLP-1 encoding gene (Fig. 3).
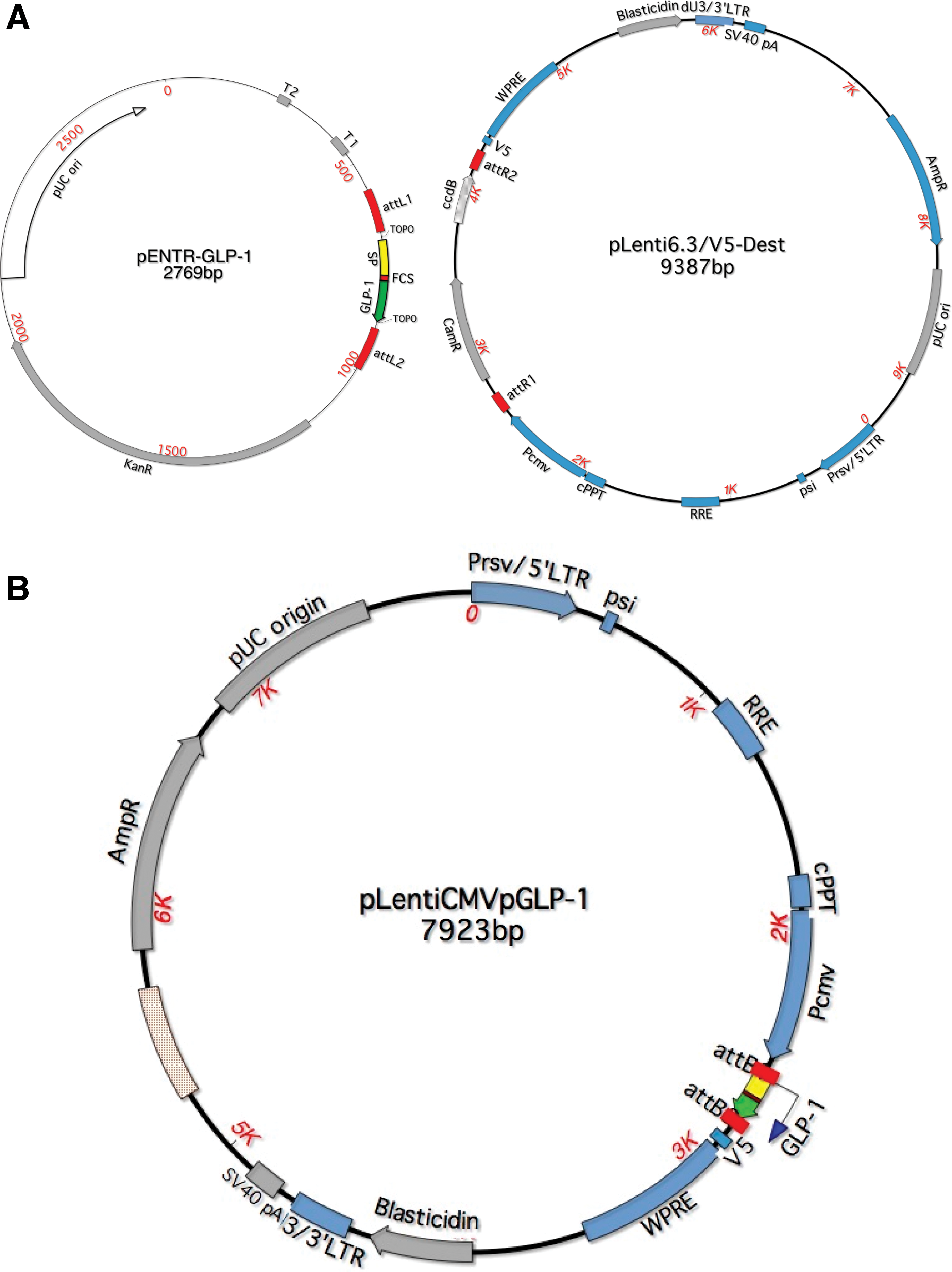
L × R recombination reaction. A recombination reaction between attL1/attL2 sites on glucagon-like peptide-1 (GLP-1) entry vector (pENTR-GLP-1) and attR1/attR2 sites on a destination vector (pLenti6.3/V5-Dest)
GLP-1 expression was first confirmed using HT1080 cells infected with the LentiGLP-1 vector. GLP-1 peptide synthesis and secretion were indeed detected by ELISA in LentiGLP-1-infected cells but not LentiLacZ-infected cells (Fig. 4A). GLP-1 binding to GLP-1 receptor on pancreatic beta cells activates adenylyl cyclase, resulting in the production of cAMP from ATP. Thus, next, a cAMP production assay was performed using the Min6 pancreatic beta cell line. LentiGLP-1 infection at MOIs of 25 and 100, but not LentiLacZ (100 MOI), produced significant amounts of cAMP in the pancreatic beta cell line (Fig. 4B). Furthermore, the insulinotropic effects of LentiGLP-1 vector were revealed via a glucose stimulated insulin secretion test (Fig. 4C). Specifically, only LentiGLP-1 led to a significant increase in insulin secretion from Min6 cells. Collectively, data from these in vitro assays suggest LentiGLP-1 encodes functional GLP-1 peptide with incretin properties. Moreover, the total insulin content of pancreatic beta cells transduced with LentiGLP-1 vector was also increased, indicating the positive effect of GLP-1 expression on insulin gene synthesis (data not shown).
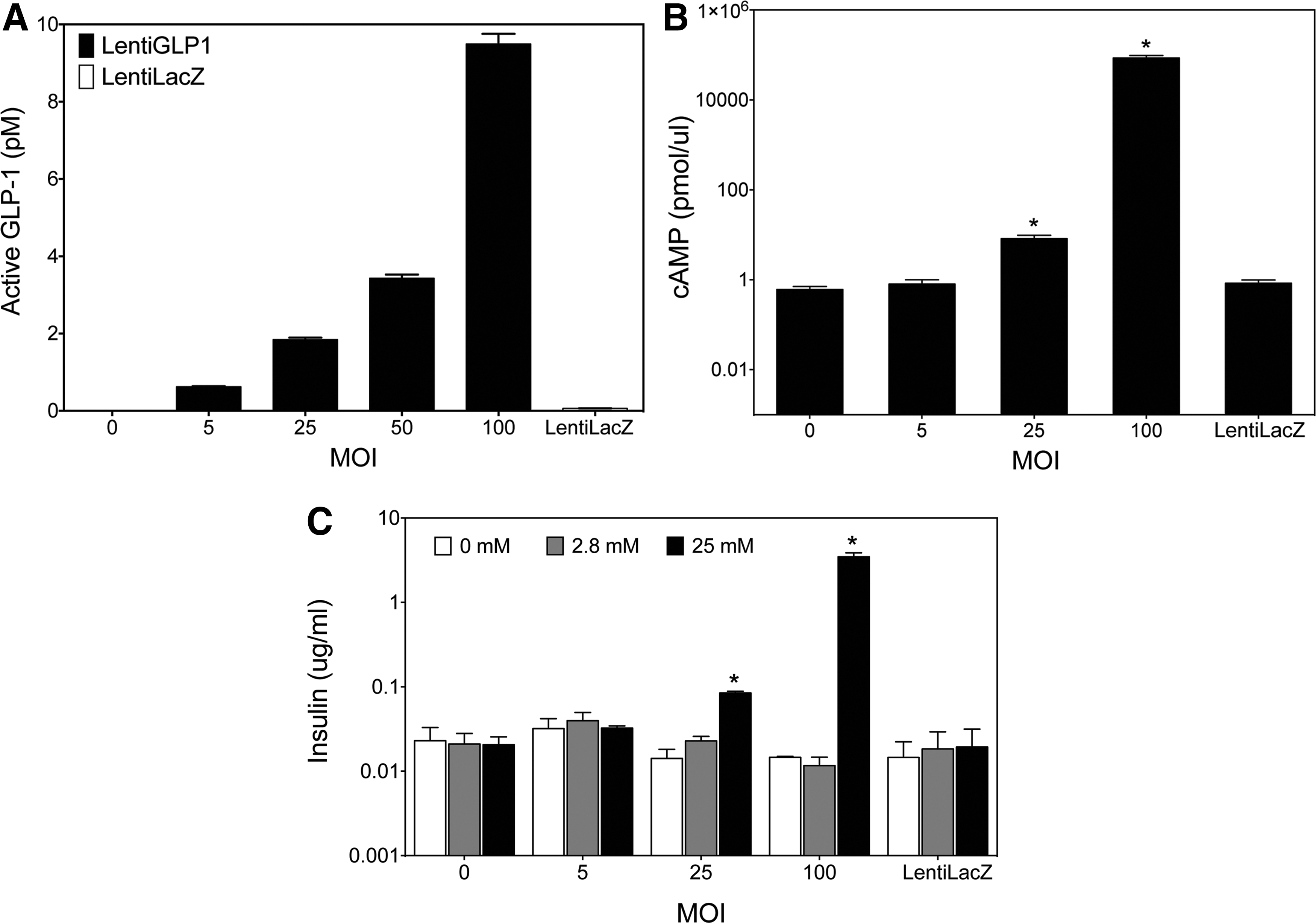
LentiGLP-1 transduction of HT1080 cells results in GLP-1 production and secretion from HT1080 cell line.
LentiGLP-1 injection decreased blood glucose levels of obese diabetic SDW rats
Having tested the LentiGLP-1 vector in vitro, next, the in vivo characteristics of this vector were examined. For these studies, a third-generation lentivirus vector pseudo-typed with vesicular stomatitis virus G glycoprotein (VSV-G) was used for increased stability and broad tissue tropism. Since GLP-1 is mainly secreted from gastrointestinal cells, the vectors were injected i.p. to transduce abdominal organs. After confirming diabetes on days 3 and 5 after STZ injection, the obese diabetic SDW rats were divided into two groups on day 7. The rats received either 1011 TU LentiGLP-1 or LentiLacZ i.p., and then blood glucose levels were followed over time. LentiGLP-1-injected obese diabetic SDW rats became normoglycemic 3 days after vector delivery, while LentiLacZ-injected obese diabetic SDW rats remained diabetic (Fig. 5A). The LentiLacZ-injected obese diabetic SDW rats were sacrificed after 1 month, while LentiGLP-1-injected SDW rats were monitored for 6 months, as they remained normoglycemic. Blood samples taken 1 month after STZ injection indicated the LentiGLP-1-injected rats displayed threefold higher levels of GLP-1 in the blood compared to control or LentiLacZ-injected rats (Fig. 5B). To reveal the effect of GLP-1 transgene expression on insulin resistance and glucose tolerance, obese diabetic SDW rats were tested for insulin sensitivity and glucose intolerance 5 weeks after gene delivery. As expected, LentiLacZ-injected obese diabetic SDW rats were completely resistant to insulin injection and remained glucose intolerant, while LentiGLP-1 injection clearly reversed insulin resistance (Fig. 5C) and improved glucose tolerance in diabetic SDW rats (Fig. 5D and E). Together, these data indicate the in vivo activity of the LentiGLP-1 vector in obese diabetic SDW rats.

LentiGLP-1 alters blood lipid profile in obese diabetic SDW rats
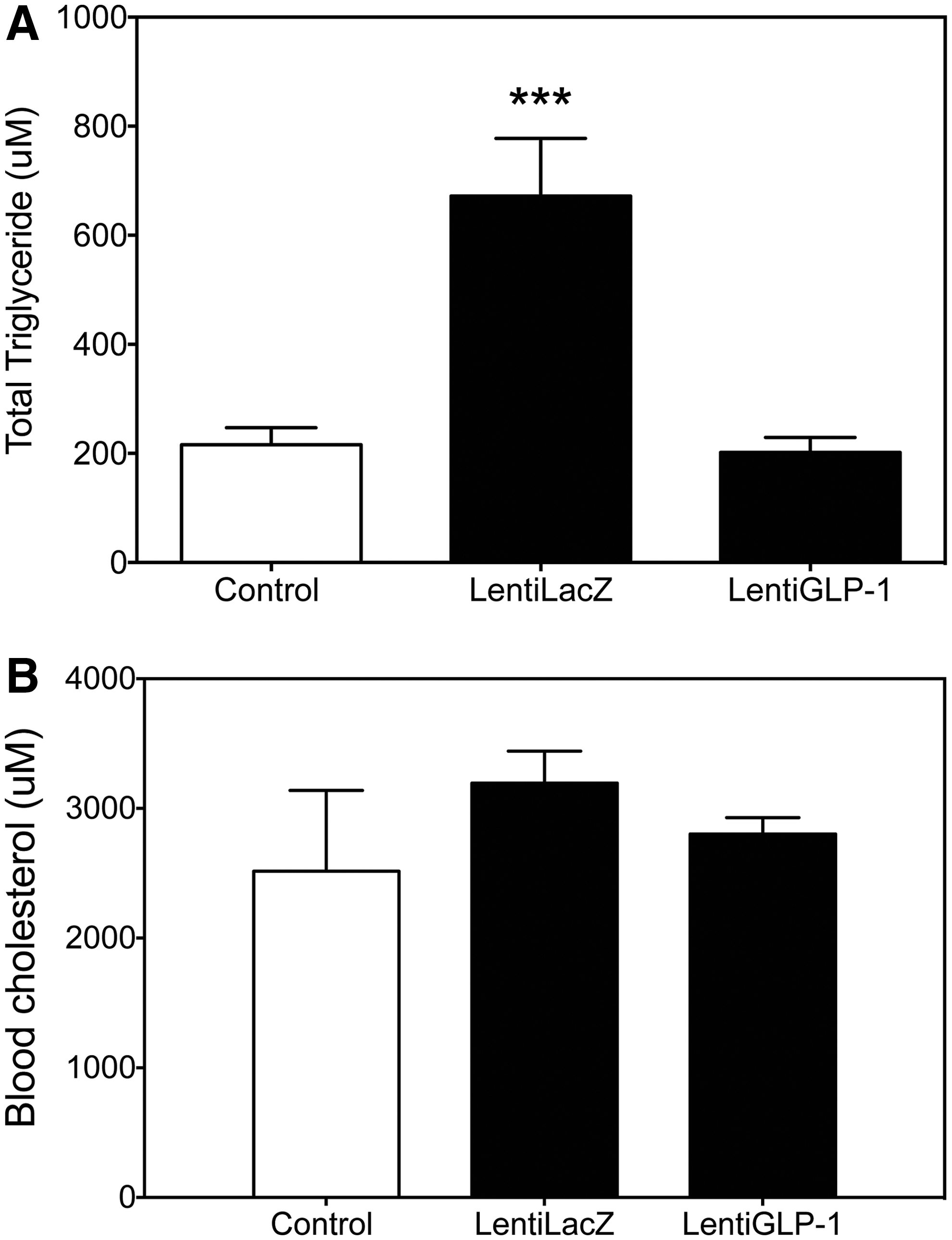
The most common causes of high triglycerides are obesity and poorly controlled diabetes. In addition to the high triglycerides, low-density lipoprotein tends to be high in obese individuals. Consequently, people with high cholesterol often have a raised level of triglycerides, predisposing them to heart disease. To determine to what extent GLP-1 treatment altered blood lipid profile, obese diabetic SDW rats were injected with either LentiLacZ or LentiGLP-1, and the blood lipid profile was analyzed 5 weeks after gene delivery. LentiGLP-1 treatment significantly reduced plasma triglyceride levels (Fig. 6A) without altering blood cholesterol in obese diabetic SDW rats (Fig. 6B). The mechanism of a differential GLP-1 effect on blood lipid profile remains to be clarified.
LentiGLP-1-infected obese diabetic SDW rats regained pancreatic beta cell mass correlating with normoglycemia
The three most common features of T2DM are insulin resistance, impaired glucose tolerance, and beta cell loss. 27 Insulin resistance is considered a major risk factor for cardiovascular disease. 28 Under the influence of environmental and/or genetic factors, insulin resistance can cause pancreatic beta cell failure, leading to glucose intolerance. During the development of T2DM, glucose tolerance is generally lost long before actual appearance of hyperglycemia. Although hyperglycemia may not be evident until significant portions of beta cells are lost, it is the end result of pancreatic beta cell loss in diabetic patients. To reveal the potential GLP-1 effect on pancreatic beta cell regeneration, rats were sacrificed 5 weeks after gene delivery, and their pancreata were examined histologically. No difference was observed between the SD-fed control and HFD-fed obese SDW rats in terms of insulin (+) area on pancreas. Immunostaining of pancreatic sections for insulin indicated LentiLacZ-infected obese diabetic SDW rats lost >90% of their pancreatic beta cells, correlating with hyperglycemia (Fig. 7A and B). In contrast, LentiGLP-1-infected rats reacquired 50% of their pancreatic islets, yielding normoglycemia. Accordingly, as LentiLacZ-treated rats exhibited low insulin scores due to a significant loss of pancreatic beta cells, LentiGLP-1-treated rats displayed insulin scores similar to those of the SD-fed control or HFD-fed obese SDW rats (Fig. 7C). These results suggest LentiGLP-1 treatment partially restores not only pancreatic beta cell mass, but also insulin gene expression in pancreatic islets.

LentiGLP-1 reduces body weight gain in obese diabetic rats
GLP-1, which is a potent inhibitor of food intake, plays a physiological role in the regulation of body weight. To reveal any potential anorexic effects of GLP-1 peptide treatment in obese diabetic rats, HFD-fed SDW rats were injected with STZ on week 14 to induce diabetes. After the confirmation of diabetes on days 3 and 5 after STZ injection, rats were divided into two groups and received either LentiGLP-1 or LentiLacZ vector IP on day 7. Body weight measurements were recorded over the following 6 weeks. LentiGLP-1-injected rats displayed a significant reduction in body weight compared to LentiLacZ-injected obese diabetic or obese control animals (Fig. 8).
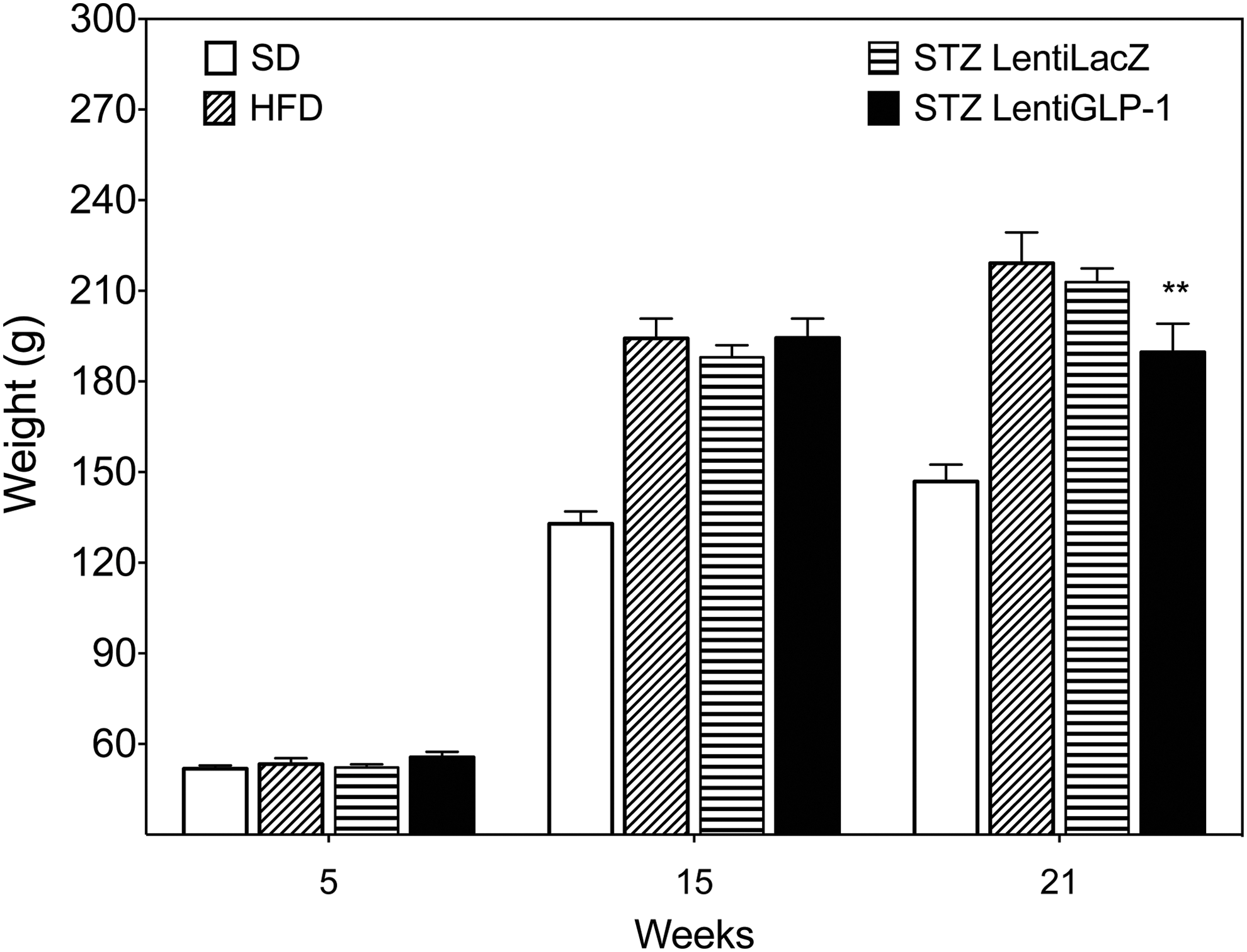
Anorexic effect of GLP-1 on diabetic obese SDW rats. Body weight in LentiGLP-1 (n = 10) versus LentiLacZ (n = 10) injected obese diabetic SDW rats is shown. Animals fed SD (n = 10) or HFD (n = 10) are also shown on the graph for comparison purposes. Lentivirus injections were performed on week 15 (STZLentiLacZ vs. STZLentiGLP-1, p = 0.0032). **p < 0.005.
In vivo abdominal tissue distribution profile of lentiviral vectors
Being a gastrointestinal hormone with major antidiabetic effects on internal organs (e.g., pancreas, GI track, stomach, heart, and kidneys), all procedures involving lentivirus delivery were done via i.p. injection. Real-time PCR analysis was performed to determine the organs infected with lentivirus vectors. Expectedly, LentiGLP-1 injection displayed a similar distribution profile compared to LentiLacZ (Table 1). To correlate gene expression to lentivirus integration, immunohistochemistry analyses of tissue sections of various intra-abdominal organs dissected from sacrificed animals following gene delivery were performed using rabbit polyclonal anti-GLP-1. GLP-1 expression was clearly detectable in the pancreas, liver, spleen, and kidney of LentiGLP-1-injected animals, leading to high levels of circulating GLP-1 in the blood (Fig. 9).

Immunohistochemistry analysis of intra-abdominal organs following lentivirus gene delivery (n = 9/group, 5 sections/animal). Control represents tissues dissected from PBS-injected HFD-fed animals treated in the absence of primary antibody (40 × magnification). Staining of alpha cells on pancreatic sections is due to cross-reactivity of GLP-1 antibody to major proglucagon fragment. Color images available online at
Intraabdominal tissue distribution profile of lentiviral vectors (n = 9/group)
Genomic DNA was isolated from intraabdominal organs, and quantitative polymerase chain reaction was conducted to determine integrated lentiviral vectors, as described in Materials and Methods. Data represent integrated vector copy numbers. Data are reported as means ± standard error of the mean.
GLP-1, glucagon-like peptide-1; HFD, high-fat diet; STZ,
Discussion
The intestinal mucosa releases incretins to promote the glucose-induced insulin secretion (insulinotropic effect) from pancreatic beta cells. Incretins are essential to suppress postprandial glucose excursions facilitating glucose entry into peripheral tissues. 29 GLP-1 and GIP are the two gastrointestinal hormones that mediate 70% of the glucose-dependent insulin secretion in humans. 30 GLP-1 released into the bloodstream following meals activates insulin biosynthesis and secretion in pancreatic beta cells via G protein coupled receptors. 31 In addition to its insulinotropic activity, GLP-1 suppresses glucagon release and improves age-dependent glucose tolerance. 32 GLP-1 also possesses mitogenic activity, causing cellular differentiation 16 and an increase in beta cell mass. 11
T2DM patients have a 50% decrease in incretin effect, as determined from isoglycemic glucose tolerance tests, compared to healthy controls. 33 Because GLP-1 infusions restored an insufficient beta cell response, it is considered a therapeutic agent for T2DM. 34 GLP-1 can effectively stimulate glucose-induced insulin secretion without causing hypoglycemia. 35 However, due to its short half-life as a result of quick inactivation by dipeptidyl peptidase-4, larger quantities or frequent injections are needed to compensate for this deficit. 30 Thus, gene delivery systems were developed to provide constant GLP-1 production and secretion in vivo. 25,36 Non-viral gene delivery techniques are limited by low transfection efficiency. Adenovirus vectors can only provide short-term therapeutic benefit due to their antigenic nature. 37,38 Adeno-associated viruses (AAVs) have limited cargo capacity and transduction efficiency. 39,40 Compared to other viral vectors, lentiviruses are the vector of choice considering longevity of transgene expression, safety, and efficacy. 41 –43 Thus, lentivirus vectors pseudo-typed with VSV-G were generated to perform GLP-1 gene transfer in diabetic rats.
A number of gene transfer protocols developed for diabetes have mainly focused on the ex vivo modification of pancreatic islets for transplantation. 44,45 The administration of viral vectors via the common bile duct 46,47 or a distal blood vessel from the pancreas 48 for the purpose of in vivo gene transfer to pancreatic islets, however, has been inefficient, prompting an investigation of other routes (i.p., intraductal, and intravenous [i.v.]) to achieve robust and prolonged expression of genes delivered by dsAAV vectors in the pancreatic islets of mice. 49 Delivery i.p. led to 5- to 10-fold more efficient pancreatic transduction versus i.v. injection, since the liver filters out a significant amount of the dsAAV administered. Interestingly, the limited diffusion capacity of the viral particles resulted in the cells of the peripheral zone of the islet, in contrast to cells within the central zone, to be preferentially transduced after i.p. delivery. These data highlight the difficulty of using viral vectors to deliver genes efficiently to pancreatic islets, largely because of the clustered architecture of endocrine cells. 50 By comparison, islet cell transduction rates can be further improved by using retrograde pancreatic intraductal delivery, an approach similar to the clinical technique of endoscopic retrograde cholangiopancreatography, compared to traditional i.p. or i.v. delivery.
Ectopic expression of GLP-1 outside the pancreas represents another therapeutic option due to the destruction of pancreatic beta cells during disease progression. GLP-1 is mainly produced and secreted by intestinal entero-endocrine L-cells located in the lower intestine (ileum) upon food consumption. The i.p. route of gene delivery was chosen for LentiGLP-1-mediated gene transfer in diabetic obese rats. Most of the previous GLP-1 gene delivery studies involved genetically modified obese rodent species, such as ob/ob or db/db mice and Zucker diabetic rats. 51 –54 Unfortunately, these models carry mutations affecting appetite (leptin [ob/ob] or leptin receptors [db/db, Zucker diabetic rats [fa/fa]), and mutation-caused diabetes is extremely rare in humans. 55 Since diabetes mostly results from the interaction between genes (predisposition) and environment (sedentary life-style), a diet-induced obesity model combined with low-dose STZ injection was used in this study to mimic better the T2DM seen in humans. 56 –58 Expectedly, prediabetic HFD rats exhibited insulin resistance and glucose intolerance, in addition to being obese. Low-dose STZ injection resulted in pancreatic beta cell loss, leading to hyperglycemia. Delivery i.p. of LentiGLP-1 into obese diabetic rats resulted in a decrease in hyperglycemia and eventually led to normoglycemia. LentiGLP-1 gene delivery also reversed insulin resistance and improved glucose tolerance in obese diabetic rats. Decreased blood glucose correlated with an increase in insulin-positive beta cells. While no alteration in blood cholesterol was detectable, serum triglyceride levels were reduced only in LentiGLP-1-injected rats. Genome integration studies conducted by quantitative PCR revealed i.p. injection of lentivirus vectors (LentiGLP-1 or LentiLacZ) resulted in successful transduction of major abdominal organs such as the liver, spleen, and kidney, including the pancreas. By doing so, GLP-1 levels were raised threefold in the blood, which might be essential to observe beneficial effects of GLP-1 in diabetic obese animals.
Several gastrointestinal peptides such as cholecystokinin, peptide tyrosine-tyrosine, and nesfatin-1 including GLP-1 have been implicated in the regulation of hunger and satiety. 59 Furthermore, continuous subcutaneous infusion of GLP-1 for 6 weeks into T2DM patients resulted in a reduction in body weight and appetite, in addition to the inhibition of gastric emptying. 60 Thus, GLP-1 peptide has a strong influence on appetite and weight maintenance due to its proposed actions on the gastrointestinal track involving reduction in gastric emptying and gut motility. As shown in this study, LentiGLP-1-treated obese diabetic rats displayed reduced body weight gain compared to LentiLacZ-treated rats—presumably due to decreased appetite. Although the synergistic actions of GLP-1 in the gut and brain, acting on both central and peripheral receptors, were suggested for the effects of GLP-1 on satiety, 61 the main mechanism of hormone action resulting in weight loss remains to be studied.
In conclusion, technical difficulties in gene transfer to the pancreas initially undermined the successful application of GLP-1 gene delivery targeting this organ. Technical improvements in gene delivery methods and generation of different vectors in recent years have allowed for a better assessment of the therapeutic efficacy of GLP-1 gene delivery in preclinical models of T2DM. Prolonged GLP-1 synthesis and secretion is achievable with current gene therapy vectors, but the most efficient gene delivery method has yet to be developed. The therapeutic effects of GLP-1 in diabetic obese rats in this study suggest GLP-1 gene delivery might be a good alternative to GLP-1 infusion or multiple daily injections. Despite all these circumstances, larger-animal models of GLP-1 gene delivery are needed to accelerate the transition from preclinical to a clinical trial in the near future.
Footnotes
Acknowledgments
This study is supported by grants from Akdeniz University Scientific Research Administration Division and the Scientific and Technological Research Council of Turkey (TUBITAK-112S114).
Author Disclosure
The authors declare no conflicts of interest that could be perceived as prejudicing the impartiality of the research reported.