
Abstract
Cross-reacting material 197 (CRM197) is a mutant form of the diphtheria toxin. Recent studies have found that CRM197 exerts an experimental antitumor effect on several types of tumors. This study applied a novel treatment of adenovirus-mediated CRM197 (AdCRM197) to human ovarian cancer cells. Interestingly, it was found that A2780 cells were sensitive to AdCRM197, but SKOV3 cells were resistant to it. Since SKOV3 cells are p53 deletion cells, while A2780 cells are p53 wild-type cells, it was postulated that p53 might play a key role in AdCRM197-induced apoptosis. This presumption was demonstrated by means of knockdown of p53 of the A2780 cells through lentivirus-mediated RNA interference. This knockdown resulted in the A2780 cells becoming resistant to AdCRM197. To verify this presumption further, the wild-type p53 gene in the SKOV3 cells was replaced with adenovirus-mediated p53 (Adp53). As expected, AdCRM197 plus Adp53 resulted in apoptosis of the SKOV3 cells. The combined treatment of AdCRM197 plus Adp53 also showed a good antitumor effect in the in vivo experiment on nude mice with xenograft tumors. Taking these results together, it is concluded that AdCRM197 induces apoptosis of human ovarian cancer cells via the p53 pathway. Moreover, it was found that Adp53 can reverse the resistance of p53-deletion human ovarian cancer cells to AdCRM197. The combination of AdCRM197 and Adp53 may be a potentially effective method for overcoming the resistance of p53-deficient human ovarian cancer to AdCRM197.
Introduction
C
Although CRM197 is often thought to be a nontoxic mutation of the diphtheria toxin, recent studies have found that CRM197 is not a nontoxic agent. CRM197 shows a weak toxicity and can inhibit cells' protein synthesis. 6 The toxicity of CRM197 is about 106 times less than that of the diphtheria toxin. 7 The protein of CRM197 was used in the therapy of several types of tumors, including oral cancer, ovarian cancer, T-cell acute lymphoblastic leukemia, colon cancer, breast cancer, and human U251 glioma cells. 8 –16 In addition, the protein of CRM197 was used in the treatment of ovarian cancer in a human Phase I study. 17
This study made a new attempt to use adenovirus-mediated CRM197 (AdCRM197) in the treatment of human ovarian cancer cells. Interestingly, A2780 cells but not SKOV3 cells died under treatment with AdCRM197. Since A2780 cells are p53 wild-type cells, while SKOV3 cells are p53-deletion cells, it was postulated that p53 might play a key role in the AdCRM197-induced apoptosis of human ovarian cancer cells. To demonstrate this presumption, p53 was knocked down in A2780 cells using lentivirus-mediated RNA interference, and wild-type p53 were restored in SKOV3 cells by means of infection with Adp53. Knockdown of p53 in the A2780 cells resulted in the cells becoming resistant to AdCRM197, and restoring p53 in the SKOV3 cells resulted in these cells becoming sensitive to AdCRM197. These findings demonstrated that p53 plays a key role in AdCRM197-induced apoptosis of human ovarian cancer cells.
Materials and Methods
Treatment of the cells
HEK 293 cells, 293T cells and two kinds of human ovarian cancer cells (A2780 and SKOV3 cells) were grown in Dulbecco's modified Eagle's medium that contained 10% fetal bovine serum. These cells were incubated with 5% CO2 at 37°C. The A2780 and SKOV3 cells received the following treatments: control group—the cells were treated with the culture medium only; EV group—the cells were infected with AdEmpty; AdCRM197 group—the cells were infected with AdCRM197 in a similar way to the EV group; Adp53 group—the cells were infected with Adp53; combination group—the cells were infected with both AdCRM197 and Adp53. The cells were infected with AdCRM197, Adp53, or AdEmpty for 2 h at a multiplicity of infection of 10. These cells were then collected for the subsequent experiments 48 h after the various treatments.
Adenovirus production
Fragment A of the CRM197 gene in the plasmid pCite-CRM197 was kindly provided by Dr. Manuel Caruso. AdCRM197 was produced using the AdEasy System. Briefly, Fragment A of the CRM197 gene was amplified with the following pairs of primers: the forward primer, which contains a Kpn I site—5′-GGAGGTACCATGGGCGCTGATGATGTTGTTG-3′; and the reverse primer, which contains a Xho I site—5′-CTTCTCGAGTTATGATCGCCTGACACGATTTC-3′. The polymerase chain reaction product of Fragment A of the CRM197 gene was then ligated into a pShuttle-CMV plasmid to create a recombinant plasmid pShuttle-CRM197. The pShuttle-p53 plasmid containing the wild-type p53 gene was made by chemosynthesis. Then, the manufacturer's instructions were followed to produce adenovirus-mediated CRM197 (AdCRM197) and adenovirus-mediated p53 (Adp53). The empty adenoviruses (AdEmpty) were produced using a similar method. The titer of the virus was calculated by counting the infected AD-293 cells in various dilutions.
Knockdown of p53
The p53 of the A2780 cells was knocked down by means of lentivirus-mediated RNA interference. The following two DNA oligonucleotides were synthesized using the method of a previous study 18 : p53-RNAi-F: 5′-GATCGGACTCCAGTGGTAATCTACTTCAAGAGAGTAGATTACCACTGGAGTCTTTTT and p53-RNAi-R: 5′-AATTAAAAAGACTCCAGTGGTAATCTACTCTCTTGAAGTAGATTACCACTGGAGTCC-3′. After annealing, the double-stranded DNA was linked into the shRNA lentivirus vector pGreenPuro (System Biosciences) to create the pGreenPuro-P53-RNAi plasmid. The pGreenPuro-P53-RNAi combined with the two other skeletal plasmids (pCMVdeltaR8.91 and pMD.G) were co-transfected into the 293T cells to produce lentivirus-P53-RNAi. The A2780 cells were then infected with the lentivirus-P53-RNAi. In addition, the p53-knockdown A2780 cells (A2780-P53-RNAi) were screened by puromycin and were checked under a microscope using green fluorescence.
Generation of A2780 cells expressing the mutant EF2 gene
To determine whether the p53 upregulation activity by AdCRM197 takes place via EF2-ADP-ribosyl activity or via inhibiting of HB-EGF, A2780 cells expressing the mutant elongation factor 2 gene (A2780-mEF2 cells) were generated. Cells expressing this mutant gene will not be killed by ADP-ribosylation of the diphtheria toxin. 19
The mutant EF2 gene with a G-to-R mutant at position 717 was synthesized, and then the mutant EF2 gene to pCDH-CMV-MCS-EF1-copGFP-T2A-Puro (a lentivirus vector plasmid containing a puromycin-resistant gene and a GFP gene) was cloned at the BamHI site and the EcoRI site. The recombinant pCDH-mEF2 plasmid, pCMVdeltaR8.91, and pMD.G were co-transfected to the 293T cells to yield the lentivirus-mEF2. The mutant EF2 A2780 cells (A2780-mEF2) were screened by puromycin and were confirmed by green fluorescence under a microscope.
Western blot analysis
Western blot analysis was performed, as previously described. 20 The mouse polyclonal antibody of CRM197 was made by the authors' laboratory (data not shown). The antibody of caspase-9 and that of cleaved caspase-3 were purchased from Cell Signaling Technology. Anti-p53 was purchased from Beyotime Biotechnology. Anti-HB-EGF and anti-Bax were purchased from Santa Cruz Biotechnology. Anti-cleaved PARP was purchased from BD Biosciences.
MTT assay, Hoechst staining, and flow cytometric analysis
Cell viability was estimated by MTT assay, as previously described. 20 Hoechst staining and flow cytometric analysis with propidium iodine (PI) staining were carried out, as described previously. 20
Experiments using xenograft mice
The xenograft nude mice were treated, as described previously with a few modifications. 21 Briefly, A2780 cells or SKOV3 cells (5 × 106cells per mouse) were injected into the flank of female nude mice. Seven days after inoculation, the mice were treated as follows: (1) phosphate-buffered saline (PBS) group: these mice were treated with an intratumoral injection of 100 μL of PBS; (2) EV group: these mice were treated with an intratumoral injection of empty viruses at a titer of 5 × 108 PFU in 100 μL of PBS twice a week; (3) AdCRM197 group: these mice were treated with an intratumoral injection of AdCRM197 at a titer of 5 × 108 PFU in 100 μL of PBS twice a week; (4) Adp53 group: these mice were treated with an intratumoral injection of Adp53 at a titer of 5 × 108 PFU in 100 μL of PBS twice a week; (5) AdCRM197 plus Adp53 group: a combination of the AdCRM197 group and the Adp53 group. Tumor size was calculated as A × B 2 × 0.52, where A represents the length and B represents the width. 20
TUNEL staining
TUNEL staining was carried out using the In Situ Cell Death Detection Kit (Roche) according to the manufacturer's instructions. The TUNEL-positive cells were counted in a blind manner. Twenty fields were randomly chosen. The apoptotic index represents the percentage of TUNEL-positive cells.
Data analysis
All values in the figures are represented as means ± standard deviation. GraphPad Prism v5 (GraphPad Software, Inc.) was used to create all the drawings, and SPSS Statistics for Windows v17.0 (SPSS, Inc.) was used to analyze the statistics. Differences between groups were analyzed by one-way analysis of variance, and then the Tukey–Kramer multiple comparison test was used to analyze the differences in the values of the two groups. A repeated-measures analysis of variance was used to analyze the differences in the tumor volume, and then Tukey's HSD test was used to analyze the differences between the two groups. The log-rank test was used to analyze the statistical significance of the survival curves. A p-value of <0.05 was considered a significant difference.
Results
A2780 cells were sensitive to AdCRM197, but SKOV3 cells were resistant to AdCRM197
The A2780 cells and the SKOV3 cells that were infected with AdCRM197 showed different sensitivity to AdCRM197. The A2780 cells were sensitive to AdCRM197, but the SKOV3 cells were resistant to AdCRM197.
The cells were also stained with Hoechst 33258 to show the changes in the nuclei. Four days after infection with AdCRM197, the cells were stained with Hoechst 33258 and were examined under a microscope. The A2780 cells infected with AdCRM197 showed cracked or condensed nuclei, but the SKOV3 cells showed no obvious change in the nuclei (Fig. 1A and B).
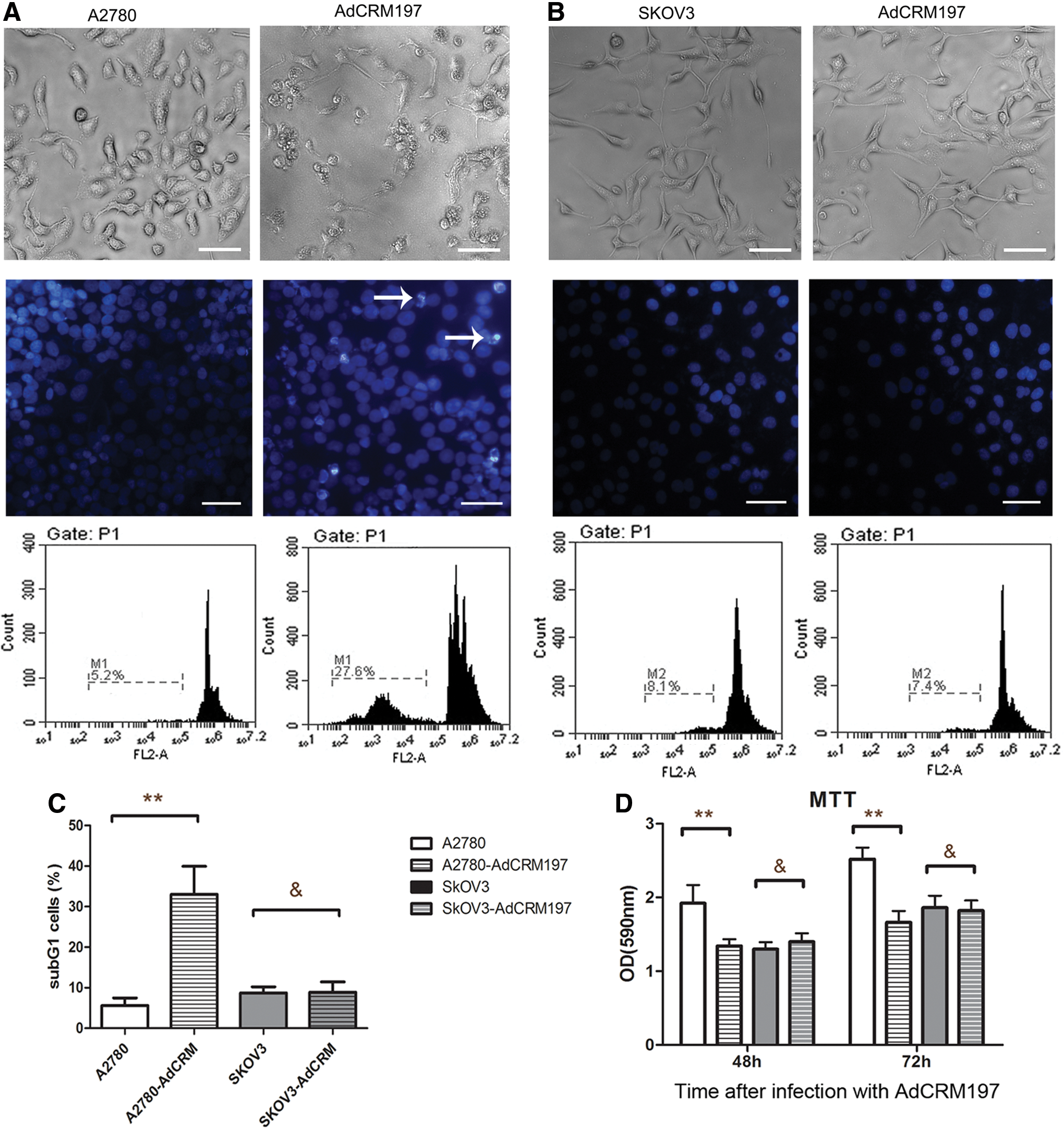
The A2780 cells were sensitive to adenovirus-mediated cross-reacting material 197 (AdCRM197), but the SKOV3 cells were not.
In the flow cytometric analysis with PI staining, the number of sub-G1 A2780 cells was significantly enhanced by infection with AdCRM197. However, the SKOV3 cells showed no obvious change after infection with AdCRM197 (Fig. 1A–C).
Cell viability was tested by MTT assay 48 or 72 h after infection with AdCRM197. The cell viability of the A2780 cells was significantly lower after infection with AdCRM197 (p < 0.01). However, the cell viability of the SKOV3 cells was not obviously changed after infection with AdCRM197 (p > 0.05; Fig. 1D).
Knockdown of p53 in the A2780 cells resulted in resistance to AdCRM197
Knockdown of p53 in the A2780 cells was performed by lentivirus-mediated RNA interference. After screening by puromycin, A2780-p53-RNAi cells were obtained. The A2780-p53-RNAi cells were green under fluorescence microscopy, as the lentivirus vector contained green fluorescent protein (Fig. 2A). Western blot analysis of the p53 was carried out in order to verify the p53 knockdown. A2780-p53-RNAi cells were successfully obtained, since they hardly expressed any p53 (Fig. 2B).

p53 knockdown of the A2780 cells by lentivirus-mediated RNA interference.
The A2780-p53-RNAi cells were then infected with AdCRM197. MTT assay and Hoechst 33258 staining were used to test the effect of AdCRM197 on the A2780-p53-RNAi cells. Unlike the A2780 cells, the A2780-p53-RNAi cells showed resistance to AdCRM197 (Fig. 3A).
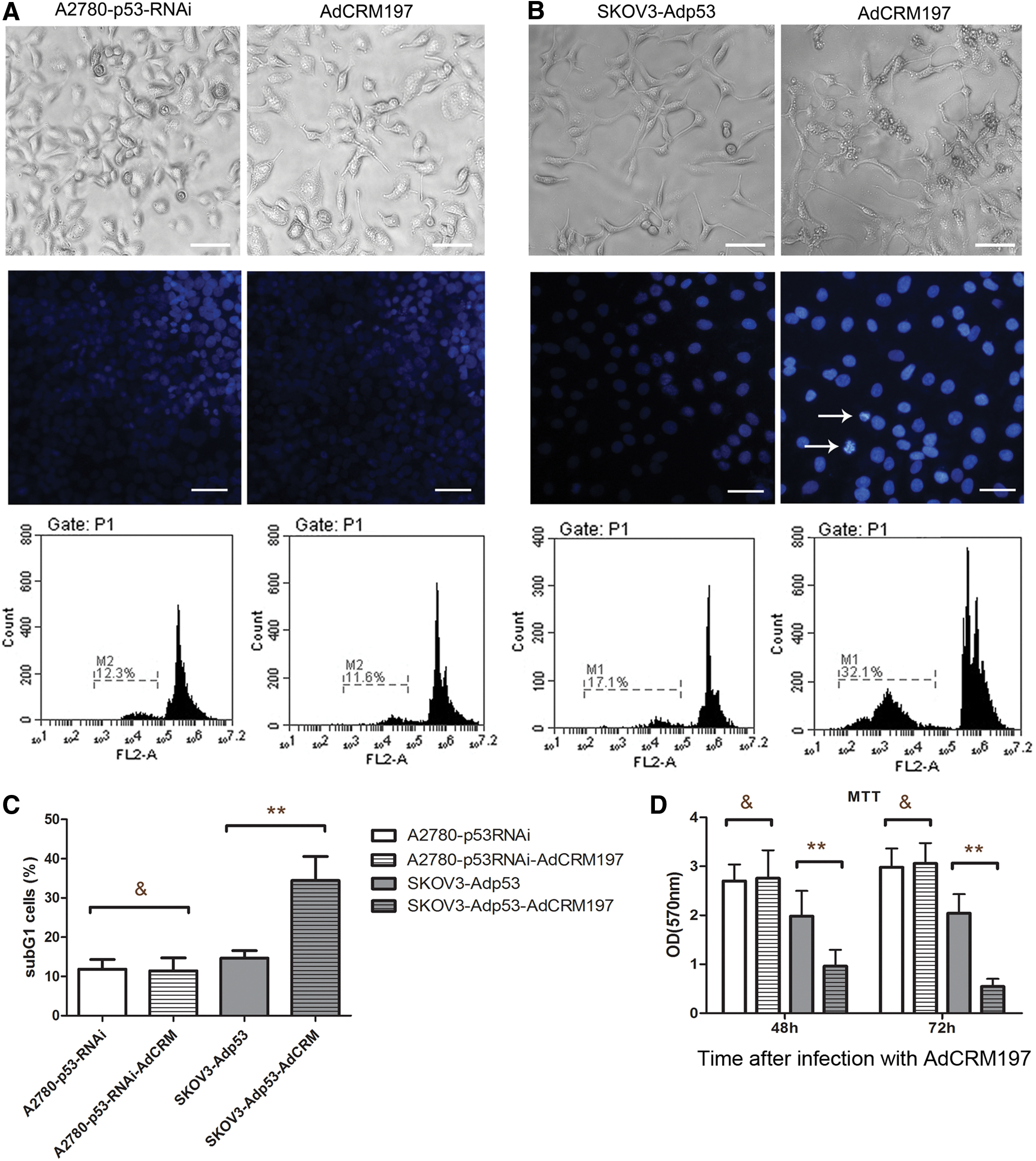
Knockdown of p53 in A2780 cells resulted in these cells' resistance to AdCRM197, restoring the p53 sensitized SKOV3 cells to AdCRM197.
The A2780-p53-RNAi cells infected with AdCRM197 showed no obvious change in the nuclei (Fig. 3A). These results were further demonstrated by flow cytometric analysis with PI staining and by MTT assay. The number of sub-G1 cells was not obviously changed by infection with AdCRM197 (p > 0.05; Fig. 3C). The viability of the A2780-p53-RNAi cells was not affected by their infection with AdCRM197 in the MTT assay 48 or 72 h after infection with AdCRM197 (p > 0.05; Fig. 3D).
Restoring the p53 sensitized SKOV3 cells to AdCRM197
p53 was restored in SKOV3 cells with Adp53. Unlike the SKOV3 cells, the SKOV3-p53 cells were sensitive to AdCRM197. After infection with AdCRM197, the SKOV3-p53 cells showed cracked nuclei or condensed nuclei (Fig. 3B), and the number of sub-G1 cells was significantly higher (p > 0.05; Fig. 3C). The viability of the SKOV3-p53 cells was significantly decreased by infection with AdCRM197 in the MTT assay 48 or 72 h after infection with AdCRM197 (p < 0.01; Fig. 3D).
p53 plays a key role in AdCRM197-induced apoptosis of human ovarian cancer cells
Expression of Bax, caspase 9, and cleaved caspase 3 was tested by Western blot analysis to show the key role of p53 in CRM197-induced apoptosis. Bax and the downstream caspases were activated by AdCRM197 in A2780 cells. However, when p53 was knocked down, Bax and the downstream caspases were not activated by AdCRM197 (Fig. 4A).

p53 was activated by EF2-ADP-ribosyl activity of AdCRM197.
Bax and the downstream caspases were not activated by AdCRM197 in SKOV3 cells. However, when p53 was restored with Adp53, Bax and the downstream caspases were activated by AdCRM197 (Fig. 4B).
p53 was activated by the EF2-ADP-ribosyl activity of AdCRM197
In order to determine the means by which AdCRM197 activated p53, A2780 cells expressing the mutant elongation factor 2 (mEF2) were generated. Unlike A2780 cells, the A2780-mEF2 cells were resistant to AdCRM197 (Fig. 4C).
Expression of p53 and of the heparin-binding epidermal growth factor-like growth factor (HB-EGF) was tested by Western blot analysis. In the A2780 cells, the expression of p53 was enhanced by AdCRM197, and the expression of HB-EGF was also elevated by AdCRM197 (Fig. 4D). However, in the A2780-mEF2 cells, the expression of p53 and of HB-EGF was not elevated by AdCRM197. These results demonstrated that p53 was activated by EF2-ADP-ribosyl activity but not by the inhibiting of HB-EGF.
Adp53 enhanced the antitumor effect of AdCRM197 in the A2780 xenograft mice
The combined therapy of AdCRM197 plus Adp53 was used on A2780 xenograft tumors to observe the antitumor effect. It was found that the antitumor effect of AdCRM197 was enhanced by Adp53.
The tumor volume of the AdCRM197-treated mice was decreased significantly in comparison to that of the two control groups (PBS group and AdEmpty group; p < 0.01). The tumor volume of the combination group (the combination of AdCRM197 and Adp53) was decreased significantly in comparison to that of the AdCRM197 group (p < 0.01; Fig. 5A). The survival time of the combination group was also clearly prolonged in comparison to that of the AdCRM197 group (p < 0.01; Fig. 5B). TUNEL assay was carried out to show the apoptotic phenomenon of the A2780 tumor cells (Fig. 5C; 200 × magnification). More apoptotic cells were detected in the mice with the combination treatment of AdCRM197 plus Adp53 than in the mice with the AdCRM197 treatment alone (p < 0.01; Fig. 5D). These results indicated that Ap53 enhanced the antitumor effect of AdCRM197 on A2780 xenograft tumors.
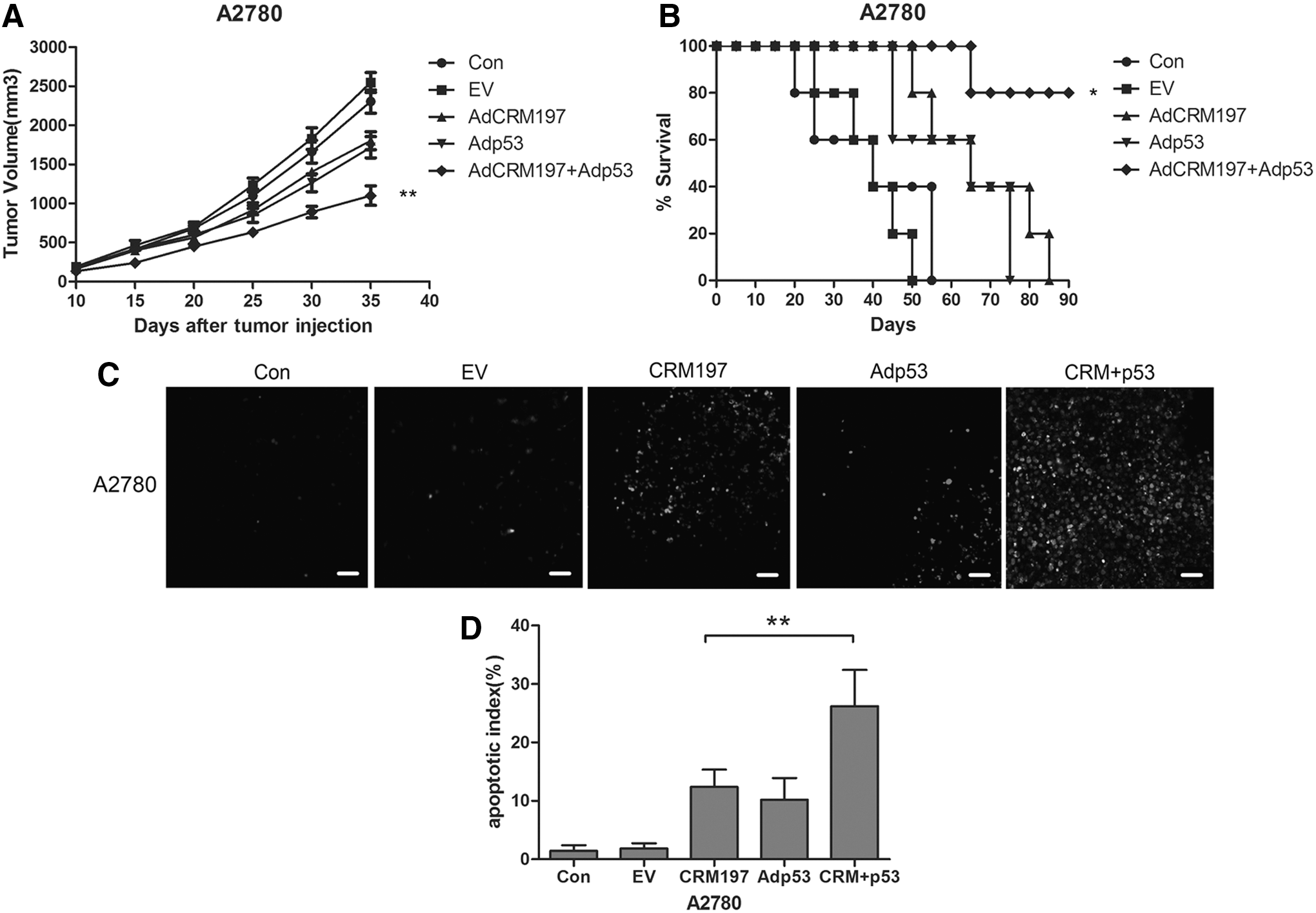
Adp53 enhanced the antitumor effect of AdCRM197 in the A2780 xenograft mice.
Combination of AdCRM197 and Adp53 is an effective way to overcome resistance of SKOV3 xenograft tumors to AdCRM197 in vivo
Based on the in vitro phenomenon whereby restoring p53 reverses the resistance of SKOV3 cells to AdCRM197, the presumption was further demonstrated in an in vivo experiment on nude mice with SKOV3 xenograft tumors.
No significant change was found in the tumor volume of the AdCRM197-treated mice in comparison to that of the two control groups (PBS group and AdEmpty group; p > 0.05). However, the combination therapy of AdCRM197 plus Adp53 resulted in a significantly decreased tumor volume in comparison to AdCRM197 therapy or Adp53 therapy (p < 0.01; Fig. 6A). The survival time of the combination group was also prolonged in comparison to that of the AdCRM197 group or the Adp53 group (p < 0.05; Fig. 6B). TUNEL assay was carried out to demonstrate the apoptotic phenomenon of the SKOV3 tumor cells (Fig. 6C; 200 × magnification). More apoptotic cells could be detected in mice that had been treated with the combination therapy of AdCRM197 plus Adp53 than in the mice treated with the AdCRM197 therapy or the Adp53 therapy (p < 0.01; Fig. 6D). These results indicated that SKOV3 xenograft tumors were resistant to AdCRM197 therapy, but the combination of AdCRM197 and Adp53 could overcome the resistance of the SKOV3 xenograft tumors to AdCRM197 in vivo.
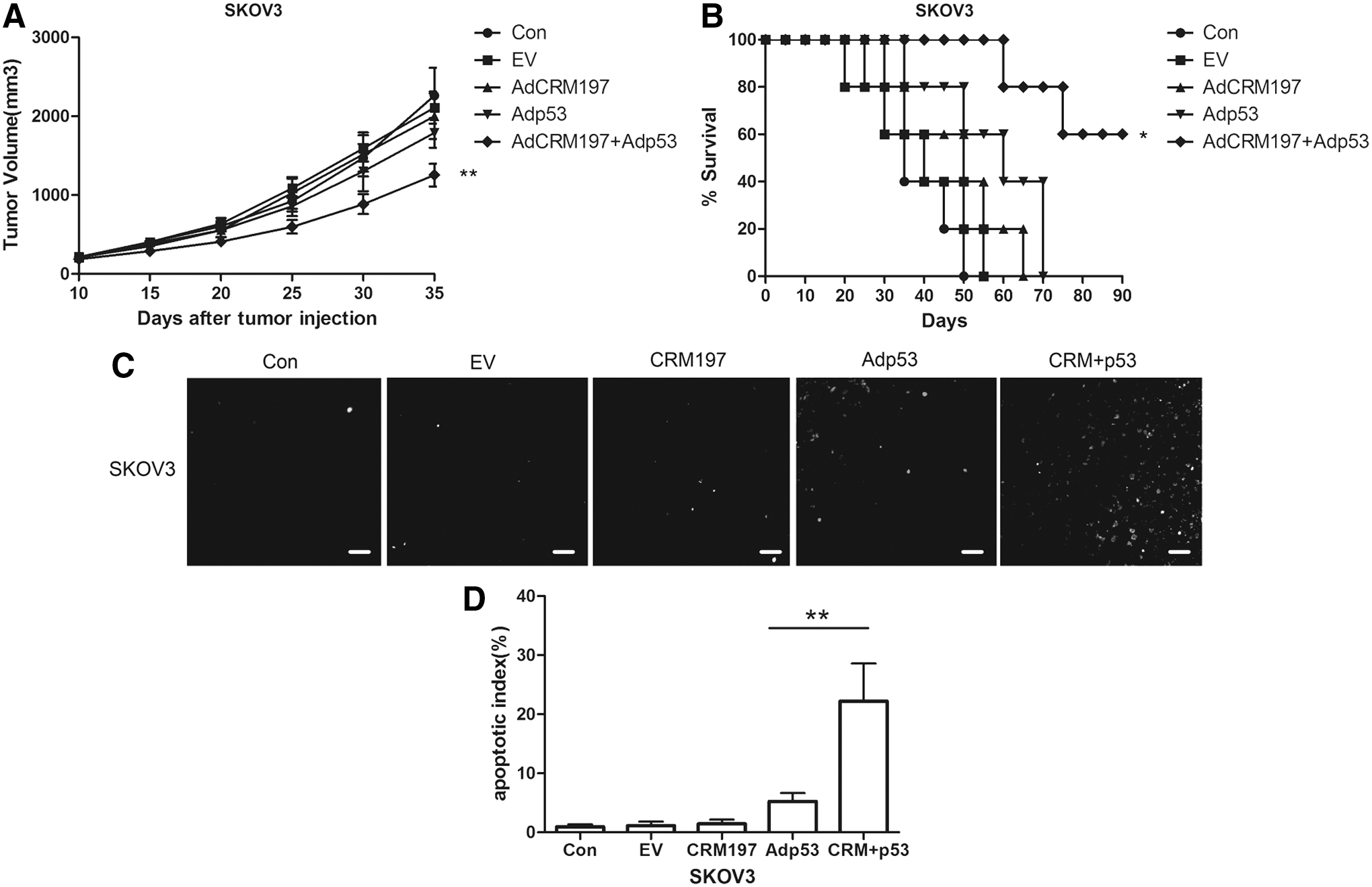
The combination of AdCRM197 and Adp53 is an effective way to overcome the resistance of SKOV3 xenograft tumors to AdCRM197.
Discussion
For a very long time, CRM197 was thought to be a nontoxic variant of the diphtheria toxin. However, recent research has revealed that CRM197 has a weak toxicity. Previous studies found that the protein of CRM197 inhibited cell growth in two ways. One way is by the protein of CRM197 binding to HB-EGF and inhibiting the expression of HB-EGF. 22 This process could result in inhibiting the mitogenic activity of the cells through its ability to inhibit EGFR binding. 21,22 The other way is through the weak EF2-ADP-ribosyl activity of CRM1977. CRM197 induces ADP-ribosylation of elongation factor 2 (EF2). The weak EF2-ADP-ribosyl activity results in the inhibiting of protein synthesis in the cells. 6
In order to determine the means by which AdCRM197 induces the death of human ovarian cancer cells and activates p53, A2780 cells expressing the mutant EF2 (A2780-mEF2) were generated. The expression of the mutant EF2 in the A2780 cells abolished the elevated expression of p53 by AdCRM197, and the A2780-mEF2 cells were resistant to AdCRM197. In addition, it was found that HB-EGF was not inhibited in the AdCRM197-infected A2780 cells. A previous study demonstrated that HB-EGF is a downstream factor of p53 and that HB-EGF will be upregulated by the activation of p53. 23 Taking these findings together, it is concluded that unlike the protein of CRM197, AdCRM197, which expresses CRM197 in cells, does not bind to HB-EGF in the suffice of the cells, and AdCRM197 does not inhibit the expression of HB-EGF. The toxicity of AdCRM197 depends on the EF2-ADP-ribosyl activity. p53 is activated by EF2-ADP-ribosyl activity but not by the inhibiting of HB-EGF.
As a tumor suppressor, p53 plays a very important role in the apoptosis of cells. p53 is a very important mediator in cell growth and apoptosis in response to many cellular stresses, including DNA damage, hypoxia, and growth factor withdrawal. 24 On the one hand, when p53 is activated, it will upregulate Bax. Bax changes its conformation and translocates from the cytoplasm to the mitochondria, and this results in the formation of pores on the mitochondrial outer membrane. 25 Then, the mitochondria releases cytochrome c and Smac, and caspase 9 is activated. On the other hand, p53 activates fas/apo1, and then caspases 8 is activated. 24 When caspase 9 or caspase 8 is activated, this will activate the executioner caspase 3.
For an efficient and high rate of gene transfer, virus vectors are widely used in gene therapy. In this study, adenoviral vectors and lentivirus vectors were both used to deliver the genes. Adenoviral vectors are very common vectors in human gene therapy. 26 –31 Adenovirus-mediated p53 is the most widely used adenovirus in cancer gene therapy. 32,33 Adenoviruses do not integrate their genes into the genome of the host cells, so adenoviral vectors are very suitable for gene therapy because of their safety. In comparison with adenoviruses, lentiviruses are relatively easy to package, and they need a relatively shorter time to be packaged. Lentivirus vectors integrate their genes into the genome of the host cells, so they are very suitable for use in generating cells that stably express the target genes.
Targeted therapy is a promising way forward in cancer therapy. 33 –47 p53 is a very popular target for cancer therapy. 48 –54 p53 deletion and p53 mutation are very common in cancer cells, especially in ovarian cancer cells. p53 deletion or mutation often results in resistance to chemotherapies. 55 In order to overcome drug resistance due to p53 mutation or deletion, p53 restoration is a viable strategy. This study demonstrated that p53 plays a key role in AdCRM197-induced apoptosis of human ovarian cancer cells by p53 knockdown in the A2780 cells and by restoring p53 in the SKOV3 cells. When p53 is knocked down, A2780 cells become resistant to AdCRM197, and when p53 is restored, SKOV3 cells become sensitive to AdCRM197. Moreover, the study demonstrated that p53 is activated by the EF2-ADP-ribosyl activity of AdCRM197. Once AdCRM197 induces the ADP-ribosylation of EF-2, p53 is activated, and then it activates Bax and the downstream caspases.
Taking these findings together, it is concluded that AdCRM197 induces the apoptosis of human ovarian cancer cells via the p53 pathway, which is a novel finding in research concerned with the antitumor effect of AdCRM197. It was also found that Adp53 can reverse the resistance of p53-deletion ovarian cancer cells to AdCRM197. The combination of AdCRM197 and Adp53 may be a potentially effective method for overcoming the resistance of p53-deficient ovarian cancer to AdCRM197.
Footnotes
Acknowledgments
This work was supported by the Foundation of the special funds for construction of high-level universities of Guangzhou University of Traditional Chinese Medicine (NO.A1-AFD018171Z11072), National Natural Science Foundation of China (Grant No. 31170007, 81773432, 81573154, 31201092, 31470080, 31201092, 31201045), Education Department of Sichuan Province Natural Science Research Program (Grant No.15ZB0250, 18ZA0148), and Program of Chengdu Medical College (Grant No. CYCG15-02).
Author Disclosure
No competing financial interests exist.