
Abstract
Congenital adrenal hyperplasia (CAH) is an autosomal recessive disorder occurring in 1:10,000 to 1:20,000 live births. In >95% of the cases, CAH results from mutations in the CYP21A2 gene, encoding the adrenal steroid enzyme 21-hydroxylase (21OH). Cardinal phenotypic features of CAH include genital ambiguity and sexual precocity, and in severe cases, neonatal salt loss and death. Current standard of care consists of lifelong oral steroid replacement to reverse the cortisol deficiency. Although significant advances in the treatment of CAH have been made, the burden of a lifelong therapeutic intervention is not ideal for quality of life. Gene therapy for CAH by adeno-associated virus (AAV) vectors has been shown to efficiently transduce the adrenal cortex, restoring normal steroidogenesis in the short term. However, adrenocortical cells are continuously renewed by stem cells located at the adrenal capsule, which differentiate as they centripetally migrate towards the adrenal medulla where they undergo apoptosis. In this context, we hypothesized that AAV-mediated genetic correction of the adrenal cortex will work short term but will eventually lead to a loss of correction. To test this hypothesis, we administered intravenously an AAV serotype rh.10 gene transfer vector (AAVrh.10-21OH-HA) to 21-hydroxylase deficient mice (21OH−/−). The data demonstrates that a single intravenous administration efficiently transduces adrenocortical cells leading to 21OH-HA expression and restoration of normal steroidogenesis. However, the duration of therapeutic efficacy lasted for only 8 weeks, accompanied by loss of 21OH-HA expression in the adrenal gland. Analysis in immunodeficient mice confirmed that the disappearance of transgene expression was not due to an antiviral/transgene immune response. Taken together, these results demonstrate that a single treatment with an adeno-associated viral vector expressing a functional copy of the mutated gene can only transiently treat adrenocortical hereditary disorders and that strategies to genetically modify the adrenocortical stem cells population will likely be required.
Introduction
C
We and others 18 –20 have demonstrated that, in the short term, adeno-associated virus (AAV) vectors are capable of genetically modifying adrenal gland cortex cells, the site of expression of steroids and thus the critical target for genetic modification to correct CAH. However, while AAV-mediated gene transfer to the adrenal cortex can work in the short term, the biology of the adrenal gland may prevent using AAV vectors for long-term correction of disorders such as CAH. In this regard, the adrenal gland cortex has a high regenerative capacity, with persistent turnover. In mice, all cortical cells are replaced within 200 days. 21 The mechanisms underlying the cortex cell turnover are not fully understood, but the standard model based on studies in mice, has identified a proliferating stem cell population located at the outermost rim at the adrenal capsule, which gives rise to adrenocortical precursor cells which become differentiated as they centripetally repopulate the adrenal gland until they reach the juxtamedullary region where they undergo apoptosis (Fig. 1). 22 –28

Schematic of the adrenal zonation and cell renewal. The adrenal gland consists of two glandular zones, an outer cortex and an inner medulla. The human and mouse adrenal cortex is subdivided into three histologically and functionally distinct layers, zona glomerulosa (mouse and human), zona fasciculata (mouse and human), and zona reticularis (human)/X-Zone (mouse). Steroids produced in the adrenal cortex inhibit the pituitary gland, leading to reduced secretion of adrenocorticotropic hormone (ACTH). Adrenocortical stem cells residing within the capsule give rise to undifferentiated progenitor and differentiated adrenal cells as they centripetally repopulate the adrenal gland until they reach the medulla boundary where they undergo apoptosis. AAV, adeno-associated virus; CAG, chicken ß-actin promoter; ITR, inverted terminal repeat.
In the context that the adrenal cortex cells are constantly replaced, we hypothesized that AAV-mediated genetic correction of the adrenal cortex will work short term but will eventually lead to a loss of correction because of the progressive turnover of cortical cells. To test this hypothesis we used the CAH H-2aw18 (21OH−/−) mouse model, containing a deletion of the 21OH genome region. 29 –31 This model comprises several phenotypic characteristics of CAH, including elevated progesterone levels, a deficiency in mineralocorticoid synthesis, and overproduction of ACTH. Unlike prior reports demonstrating AAV-mediated correction of a murine model of CAH in the short term, 18 –20 the present study was carried for a time sufficient for adrenocortical cells to be replaced as part of the normal biology of the adrenal cortex. 21 The data supports the hypothesis. AAV constructs were produced with a transgene encoding an hemagglutinin (HA)-tagged 21OH to facilitate a sensitive assay for the detection of the 21OH protein; we and others have shown that HA tags do not affect function. 20,32 While a single intravenous administration of an AAV serotype rh.10 gene transfer vector coding for human 21OH-HA efficiently transduces adrenocortical cells leading to high levels of 21OH-HA expression and restoration of normal steroidogenesis in the short term, consistent with the biology of the adrenocortical cell turnover, the transgene expression eventually waned and therapeutic efficacy lasted only up to 8 weeks, a time period when all zona glomerulosa cells and half of the zona fasciculata cells are known to be replaced due to cellular turnover. 21 The data further demonstrates that the loss of expression was not related to immunity against the vector and/or transgene, as a similar loss of transgene expression was also observed in NOD scid gamma (NSG) immunodeficient mice. Based on these observations, we conclude that AAV vectors are not suitable for persistent genetic modification of the adrenal cortex and will not be useful for disorders such as CAH.
Methods
Vector production
For the initial studies, two AAV capsids (AAV9 and rh.10) were compared for efficiency of adrenal cortex transduction (Fig. 2A). The vectors were identical except for the capsids. The genomes of both were comprised 5′ to 3′: the AAV2 5′ inverted terminal repeat, the AAV2 packaging signal (Ψ), cytomegalovirus enhancer, chicken β-actin promoter with splice donor and rabbit β-globin intron with splice acceptor, the human 21OH cDNA with a HA tag for detection, rabbit β-globin polyA signal, and the AAV2 3′ inverted terminal repeat packaged into rhesus serotype rh.10 capsid. The two vectors were produced as described previously. 33 Briefly, the expression plasmid pAAV2-21OH-HA (600 μg) and the AAVrh.10 packaging-Ad helper hybrid plasmids (pPAKMArh.10 for the AAVrh.10 vector or pPAKMA9 for the AAV9 vector; both 1,200 μg) were co-transfected using polyethylenimine (PEI) transfection reagent (Polysciences, Warrington, PA) into 293T cells containing an integrated copy of the AdE1 gene. At 72 h posttransfection, cells were harvested and crude viral lysate was prepared using five freeze–thaw cycles, and the lysate was clarified by centrifugation at 3,500 g for 15 min. The vectors were purified from the crude viral lysate by iodixanol gradient followed by Q Sepharose® High Performance (QHP) anion exchange chromatography (GE Healthcare, Piscataway, NJ). The purified vectors were concentrated using a BioMax 100K membrane concentrator (Millipore, Billerica, MA) and stored in phosphate-buffered saline (PBS), pH 7.4 at −80°C.
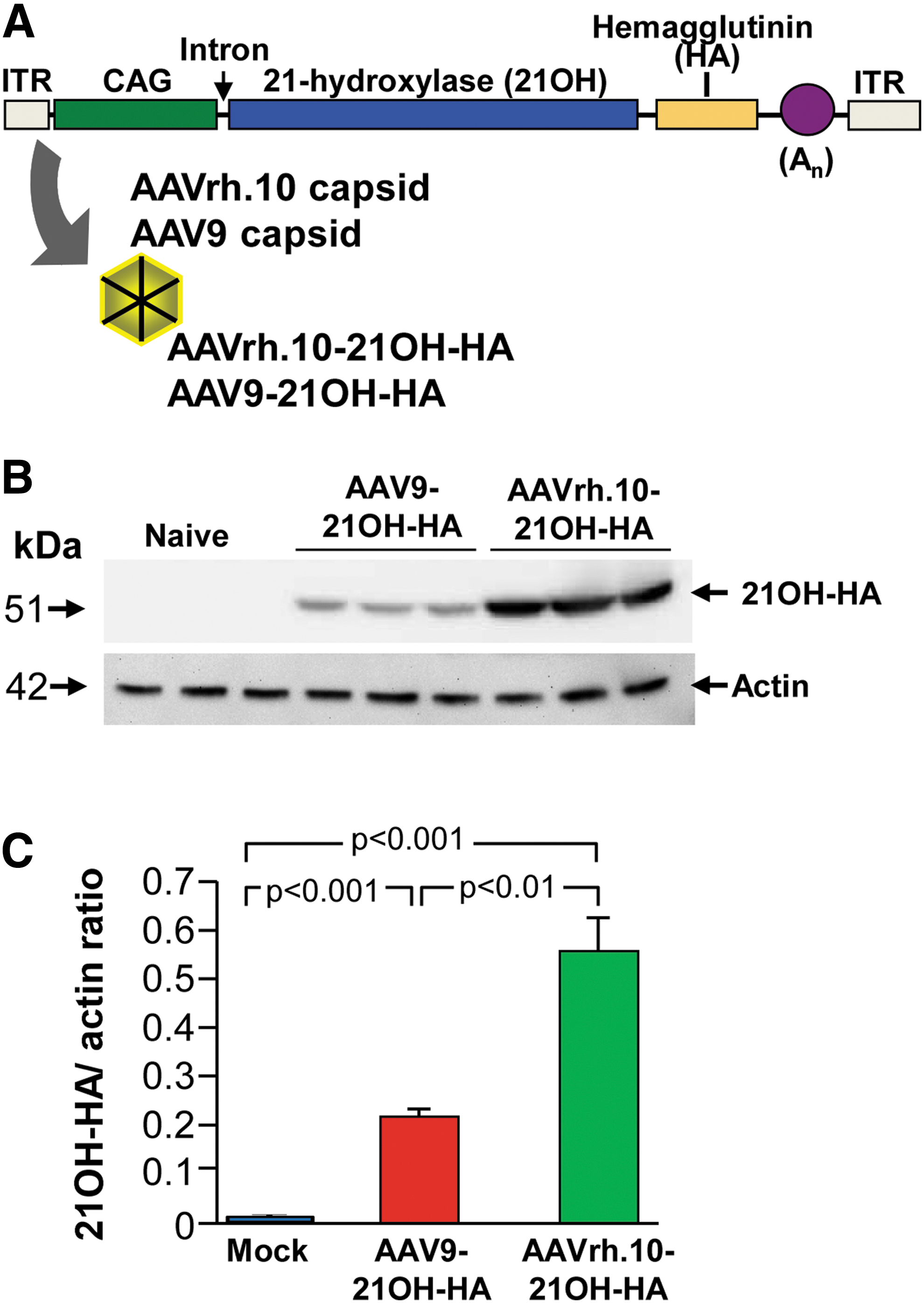
Expression of AAVrh.10-21OH-HA and AAV9-21OH-HA vectors in the adrenal gland. The two vectors are identical except for the capsid.
Vector genome titers were determined by TaqMan real-time PCR using a cytomegalovirus–chicken ß-actin promoter (CAG)-specific primer–probe set (forward primer: GTCAATGGGTGGAGTATTTACGG and reverse primer: AGGTCATGTACTGGGCATAATGC), designed using Primer Express software (Applied Biosystems, Foster City, CA). The purified vectors were digested with proteinase K in the presence of 0.5% sodium dodecyl sulfate and 25 mM ethylenediaminetetraacetate at 70°C for 1 h followed by inactivation of the protease at 95°C for 15 min. This material was used as a template for TaqMan analysis using an AAV-CAG-hCLN2 plasmid DNA standard of known copy number to generate a standard curve.
In vivo expression
To evaluate the most efficient vector for adrenal gland transduction and to analyze the distribution of transgene expression in the adrenal gland over time, 7-week-old wild-type mice were administered with 6.5 × 1011 gc of AAVrh.10-21OH-HA or AAV9-21OH-HA by intravenous injection via the tail vein in 100 μL volume. Mice were euthanized 2, 6, 16, and 32 weeks postadministration using isoflurane followed by cardiac perfusion with 40 mL cold PBS, pH 7.4. Adrenal glands were collected and immediately frozen on dry ice or fixed overnight with 4% paraformaldehyde. Frozen left adrenal glands were homogenized in 50 μL lysis buffer (10 mM HEPES-KOH, pH 7.4, 5 mM mannitol, and 1% Triton X-100 in water) using a pestle mixer (VWR International). The homogenate was centrifuged at 10,000 g for 5 min, and supernatant was collected and stored in aliquots at −80°C until use. Thirty μg protein was loaded on a 4–12% Bis-Tris Mini-SDS gel (Novex) and transferred to a polyvinylidene difluoride membrane. Expression of 21OH-HA was evaluated by incubation of the membrane with mouse-anti-HA (Sigma) and peroxidase-conjugated goat-anti-mouse (Santa Cruz) antibody and enhanced chemiluminescence Western blot detection reagent (GE Healthcare). Paraformaldehyde fixed adrenal glands were embedded in paraffin and cut into 5 μm sections. Detection of 21OH-HA expression in the adrenal gland was evaluated with an anti-HA antibody (Histowiz, Brooklyn, NY). Following sectioning, anti-HA immunohistochemical staining, and counterstaining with hematoxylin, digital images of adrenal cross sections were acquired using a 40 × objective. A lattice containing rectangular counting areas corresponding to 50 μm wide columns was digitally applied to the sections such that the counting columns were perpendicular to the adrenal cortical layer spanning the cortex from medulla to capsule. The percentage of HA-positive cells was determined by counting the number of hematoxylin-positive nuclei that were present in cells that were either negative or positive for HA staining. Six randomly selected 50 μm columns were quantified per histological section.
Homozygous 21OH−/− mice
To assess the therapeutic efficacy of viral vector for CAH, 21OH-deficient mice were used as the prototype hereditary adrenocortical disease. Heterozygous 21OH-deficient mice C57BL/10snSlc-H-2aw18/H-2b were kindly provided by Dr. Toshihiko Shiroishi (Institute of Genetics, Japan). Heterozygous mice were crossed with Balb/c and C57BL/6 wild-type mice (Jackson Labs, Bar Harbor, ME) to receive heterozygous mice in a mixed C57Bl/6-Balb/c background. In order to produce homozygous mice for this study, mice were kept in a mixed background and bred as heterozygous pairs with genotyping of pups at weaning via PCR. Because most homozygous (21OH −/−) pups died soon after birth, all dams received 5 μg dexamethasone subcutaneously every other day.
Assessment of serum progesterone and ACTH levels
To analyze the therapeutic efficacy of the AAVrh.10-21OH-HA vector over time, progesterone and adrenocorticotropic hormone (ACTH) levels were measured before and after treatment of 21OH−/− mice and compared with age-matched wild-type and untreated 21OH−/− mice. At 7 weeks of age, 21OH−/− mice were administered with 6.5 × 1011 gc of AAVrh.10-21OH-HA by intravenous injection via the tail vein in 100 μL volume. Blood (100 μL) was collected in Microtainer blood collection tubes (Fisher Scientific, Waltham, MA) from the tail vein of the mice at various time points and centrifuged at 10,000 g for 10 min to collect serum. Progesterone and ACTH levels were evaluated in 5 μL mouse sera by using a rat/mouse progesterone enzyme-linked immunosorbent assay (ELISA) kit (Alpco, Salem, NH) and a rat/mouse ACTH radioimmunoassay kit (Phoenix pharmaceuticals, Burlingame, CA) according to manufacturer's protocol.
Assessment of anti-vector/anti-transgene immunity on transgene expression
To determine whether the loss of 21OH-HA expression was secondary to anti-capsid and/or transgene immunity, immunodeficient NSG mice were intravenously injected with 6.5 × 1011 gc of AAVrh.10-21OH-HA. At 2, 6, and 16 weeks after administration, mice were euthanized and adrenal glands were isolated as described above. Antibodies in serum of AAVrh.10-21OH-HA treated NSG, 21OH−/− and untreated wild-type mice were analyzed by Western blot analysis, ELISA, and neutralizing assay antibody titer.
Western analysis
Adrenal gland homogenate protein (20 μg) and 5 × 1010 genome copies of AAVrh.10-21OH-HA were loaded on a 4–12% Bis-Tris Mini-SDS gel (Novex, Carlsbad, CA). Serum (1:500 dilution) of an AAVrh.10-21OH-HA injected 21OH−/− mouse (6 weeks after administration), serum (1:500 dilution) of an AAVrh.10-21OH-HA–injected NSG mouse (12 weeks after administration) and anti-HA antibody (Sigma-Aldrich) were used as primary antibodies. Horseradish peroxidase–conjugated goat anti-mouse antibody (Santa Cruz) and enhanced chemiluminescence Western blot detection reagent (GE Healthcare) were used for detection.
Anti-AAVrh.10 total antibodies analysis
Anti-AAVrh.10 antibody titers were measured using ELISA. Flat-bottomed 96-well ELISA plates (Corning, New York, NY) were coated with 1 × 109 genome copies per well AAVrh.10–human alpha-1 antitrypsin in carbonate buffer, pH 9.4, overnight at 4°C. The wells were blocked with 5% milk in PBS for 30 min at 23°C. The plates were then incubated with diluted sera (1:1,000 to 1:128,000) from AAVrh.10-21OH-HA administered or naive mice for 90 min at 23°C. The plates were washed with PBS plus 0.05% Tween 20 (PBS-Tween) and then incubated with horseradish peroxidase–conjugated goat anti-mouse immunoglobulin G (Santa Cruz) in 1% milk in PBS for 90 min, 23°C. The plates were washed with PBS-Tween followed by PBS and were treated with peroxidase substrate (BioRad, Hercules, CA). The reaction was stopped by addition of 2% oxalic acid 20 min later and absorbance was measured at 415 nm. Anti-AAVrh.10 antibody titers were calculated by interpolation of the log(OD)−log(dilution) with a cutoff value equal to twofold the absorbance of background.
AAVrh.10 neutralizing antibody titer analysis
Serum AAVrh.10 neutralizing antibody titers were determined by an in vitro assay with 293-ORF6 cells in 96-well plates. AAVrh.10-Luciferase (3,000 genome copies/cell) was incubated with diluted sera (1:20 to 1:5,760) from AAVrh.10-21OH-HA administered or naive mice at 37°C for 45 min before adding the mixture to the 293-ORF6 cells. At 48 h postinfection, luciferase activity was assessed in cell lysate using Luciferase Assay System (Promega, Madison, WI). The neutralizing antibody titer was expressed as the reciprocal of serum dilution at which 50% inhibition of AAVrh.10-Luciferase was observed. 33
Statistics
All data are presented as means ± standard deviation, and comparisons between groups were conducted by a two-tailed, unpaired t-test.
Results
Comparison of in vivo expression of the AAV9-21OH-HA and AAVrh.10-21OH-HA vectors
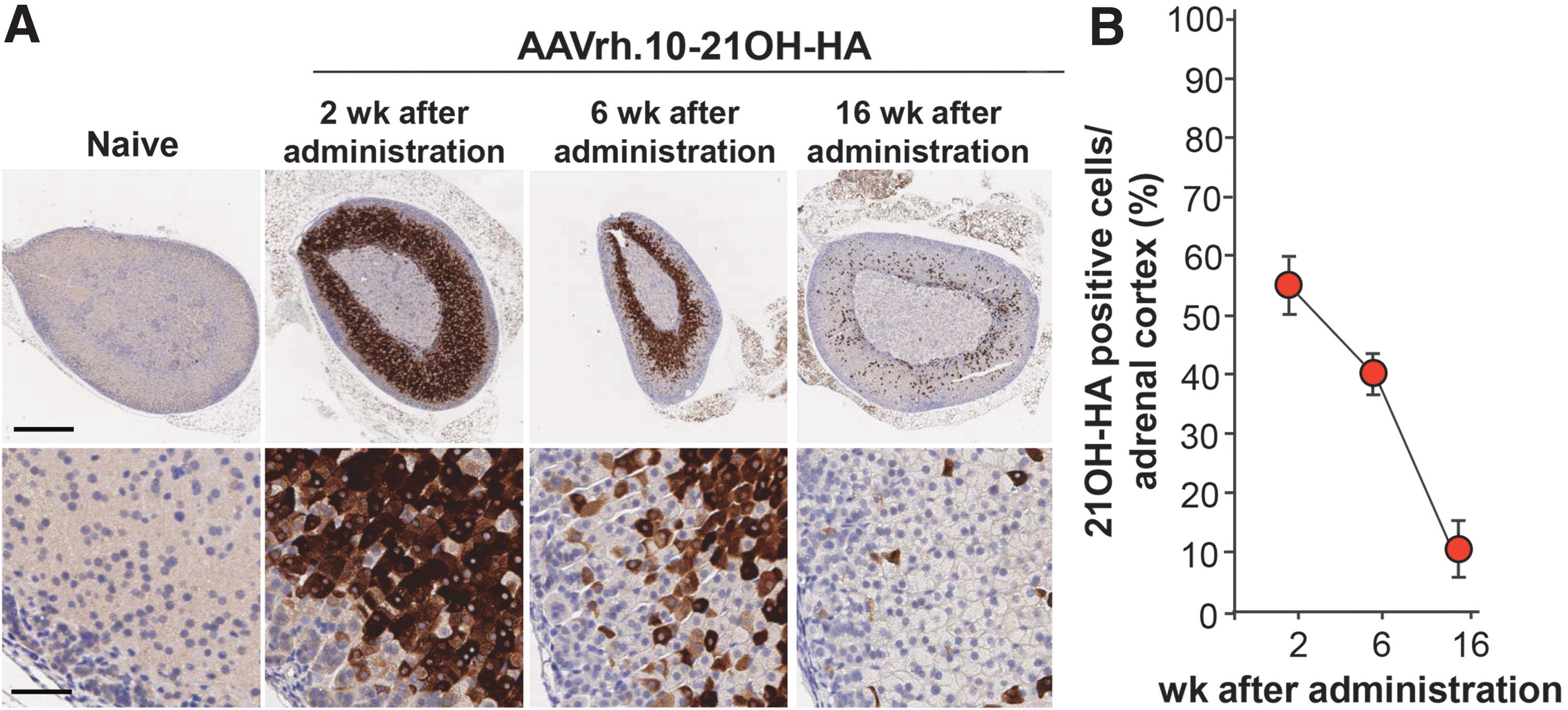
To analyze which of the two vectors were most efficient for transducing the adrenal gland cortex, serotype 9 and rh.10 vectors were intravenously administered to wild-type mice (n = 3) with the same dose (6.5 × 1011 genome copies; Fig. 2B). Two weeks after administration, the left adrenal glands were analyzed for 21OH-HA expression. The AAVrh.10-21OH-HA vector mediated significantly higher transgene expression compared with AAV9-21OH-HA (Fig. 1B and C). At 2 weeks, transgene expression was detected in zona fasciculata and the X-zone, but not in the capsule, zona glomerulosa, or medulla (Fig. 3A), with 64.8 ± 4.8% of 21OH-HA–positive adrenocortical cells (Fig. 3B). The amount of transgene expressing adrenal cells waned over time. At 6, 16, and 32 weeks after administration, only 41.0 ± 14.3%, 8.3 ± 3.9%, and 2.1 ± 1.2% of the adrenocortical cells expressed 21OH-HA, respectively (Fig. 2A and B). By comparison, the liver of the AAVrh.10-21OH-HA treated mouse had substantial levels of 21OH-HA at 32 weeks, whereas no detectable expression was observed in the adrenal (Fig. 3A and Supplementary Fig. S1; Supplementary Data are available online at

21OH-HA expression in the adrenal gland at different time-points after intravenous AAVrh.10-21OH-HA delivery. Wild-type mice received 6.5 × 1011 genome copies of AAVrh.10-21OH-HA by intravenous administration.
Similar expression patterns were seen in AAV9-21OH-HA transduced adrenal glands (Supplementary Fig. S2A and B). Based on the higher 21OH-HA expression levels generated by the rh.10 vector, this vector was used for all subsequent studies.
Progesterone and ACTH levels in 21OH−/− mice after AAVrh.10-21OH-HA delivery
The conversion of progesterone to deoxycorticosterone in the mouse adrenal gland is catalyzed by 21OH. 34 Thus, 21OH deficiency leads to accumulation of progesterone, which can be measured in blood. 30 To analyze the therapeutic efficacy of AAVrh.10-21OH-HA, progesterone levels were measured before and after treatment of 21OH−/− mice and compared with age-matched wild-type and untreated 21OH−/− mice (Fig. 4A). The 21OH−/− mice, treated at 7 weeks of age, had progesterone levels in the serum before treatment from 487 to 143 ng/mL with an average of 243 ng/mL (0 weeks after administration). Two weeks after treatment, progesterone levels decreased significantly (21OH−/− mice vs. AAVrh.10-21OH-HA treated 21OH−/− mice at 2 weeks after administration: p < 0.02) to 22.2 ng/mL ±14.8 ng/mL, similar to wild-type levels (2.26 ± 1.03 ng/mL) (wild-type mice vs. AAVrh.10-21OH-HA treated 21OH−/− mice at 2 weeks after administration: p > 0.06), whereas the levels of untreated, age-matched 21OH−/− mice did not change (162 ± 57 ng/mL). Progesterone levels of AAVrh.10-21OH-HA treated 21OH−/− mice remained similar to wild-type levels until 6 weeks after administration but increased thereafter. The duration of therapeutic efficacy lasted until 8 weeks after administration. This is the last time point at which significant differences could be detected in progesterone levels between age-matched AAVrh.10-21OH-HA treated and untreated 21OH−/− mice (21OH−/− mice vs. AAVrh.10-21OH-HA treated 21OH−/− mice at 8 weeks after administration, p < 0.005). This loss of efficacy did not change up to 32 weeks after administration, the last time point measured.
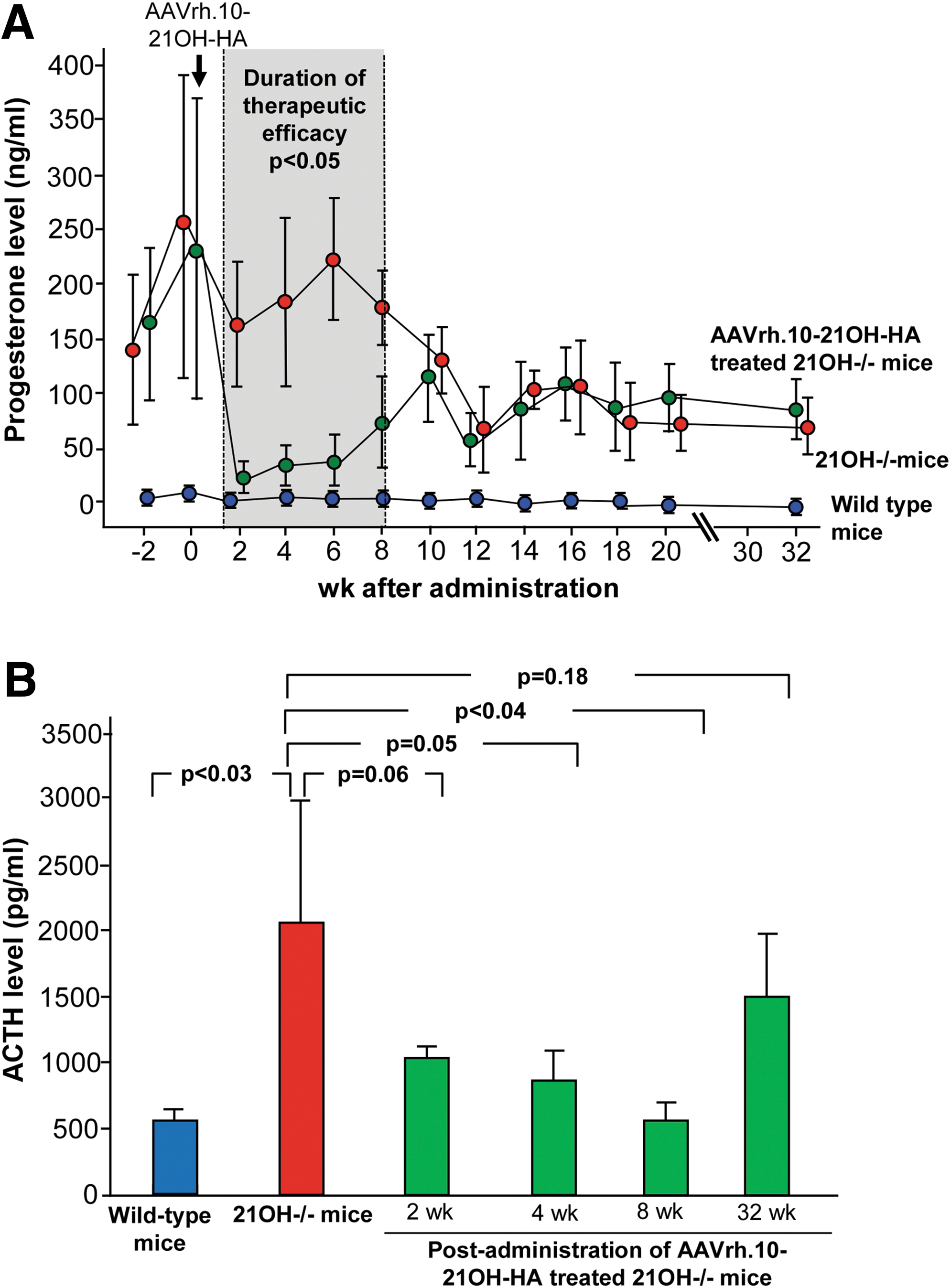
Progesterone and ACTH levels in 21OH−/− mice treated with AAVrh.10-21OH-HA. Seven weeks old 21OH−/− female mice received 6.5 × 1011 genome copies AAVrh.10-21OH-HA by intravenous administration.
In wild-type mice, mineralocorticoids secreted from the adrenal gland inhibit the pituitary gland resulting in decreased release of adrenocorticotropin (ACTH; Fig. 1). In 21OH-deficient mice, the defect in mineralocorticoid biosynthesis leads to hypersecretion of ACTH, and thus, the impact of AAVrh.10-21OH-HA administration on ACTH levels in the blood would provide an additional assessment of the efficacy of the treatment. ACTH levels of untreated 21OH−/− mice varied between 1,898 to 3,888 pg/mL and significantly decreased 8 weeks after intravenous delivery of AAVrh.10-21OH-HA to 560 ± 136 pg/mL, similar to wild-type levels (389 ± 178 pg/mL; Fig. 4B). Consistent with the progesterone levels, 32 weeks after administration ACTH levels increased to the pre-administration level of 1,505 pg/mL ±468 pg/mL, consistent with the loss of therapeutic efficacy.
Impact of immunity against vector/transgene on transgene expression
To analyze whether waning of transgene expression was due to an anti-vector and/or anti-transgene immune response, 6.5 × 1011 gc AAVrh.10-21OH-HA were administered to immunodeficient NSG mice. Two weeks after administration, 21OH-HA expression could be detected in the zona fasciculata and X-zone (Fig. 5A) with transduction efficacy of 54.4 ± 5.2% of adrenocortical cells (Fig. 5B). At 6 weeks and 16 weeks after administration, transgene expression decreased to 39.8 ± 2.9% and 10.1 ± 4.8%, respectively (Fig. 5B), similar to the expression levels in AAVrh.10-21OH-HA injected wild-type mice at this time points (wild-type mice vs. NSG mice 6 weeks postadministration: p = 0.9; wild-type mice vs. NSG mice 16 weeks postadministration: p = 0.3; Fig. 3B).
Expression of 21OH-HA in immunodeficient NOD scid gamma (NSG) mice. NSG mice received 6.5 × 1011 genome copies AAVrh.10-21OH-HA by intravenous administration.
Western analysis using AAVrh.10-21OH-HA treated 21OH−/− mouse serum as first antibody revealed the presence of antibodies against the transgene and all capsid proteins (VP1, VP2, VP3) of the vector, whereas AAVrh.10-21OH-HA treated NSG mouse serum did not contain these antibodies (Fig. 6A). All 21OH−/− mice that received AAVrh.10-21OH-HA developed anti-AAVrh.10 antibodies and anti-AAVrh.10 neutralizing antibodies in serum (Fig. 6B, C). At 2 weeks after administration, anti-AAVrh.10 antibodies titers increased significantly and remained high for the length of the study (Fig. 6B). Anti-AAVrh.10 neutralizing antibody titer peaked at 6 weeks after administration (Fig. 6C). As expected, these antibodies could not be detected in serum of AAVrh.10-21OH-HA treated NSG mice or untreated wild-type mice. Together, these data indicate that the loss of therapeutic efficacy was not due to immune responses against the vector/transgene, most likely due to the turnover of adrenal cortex cells.
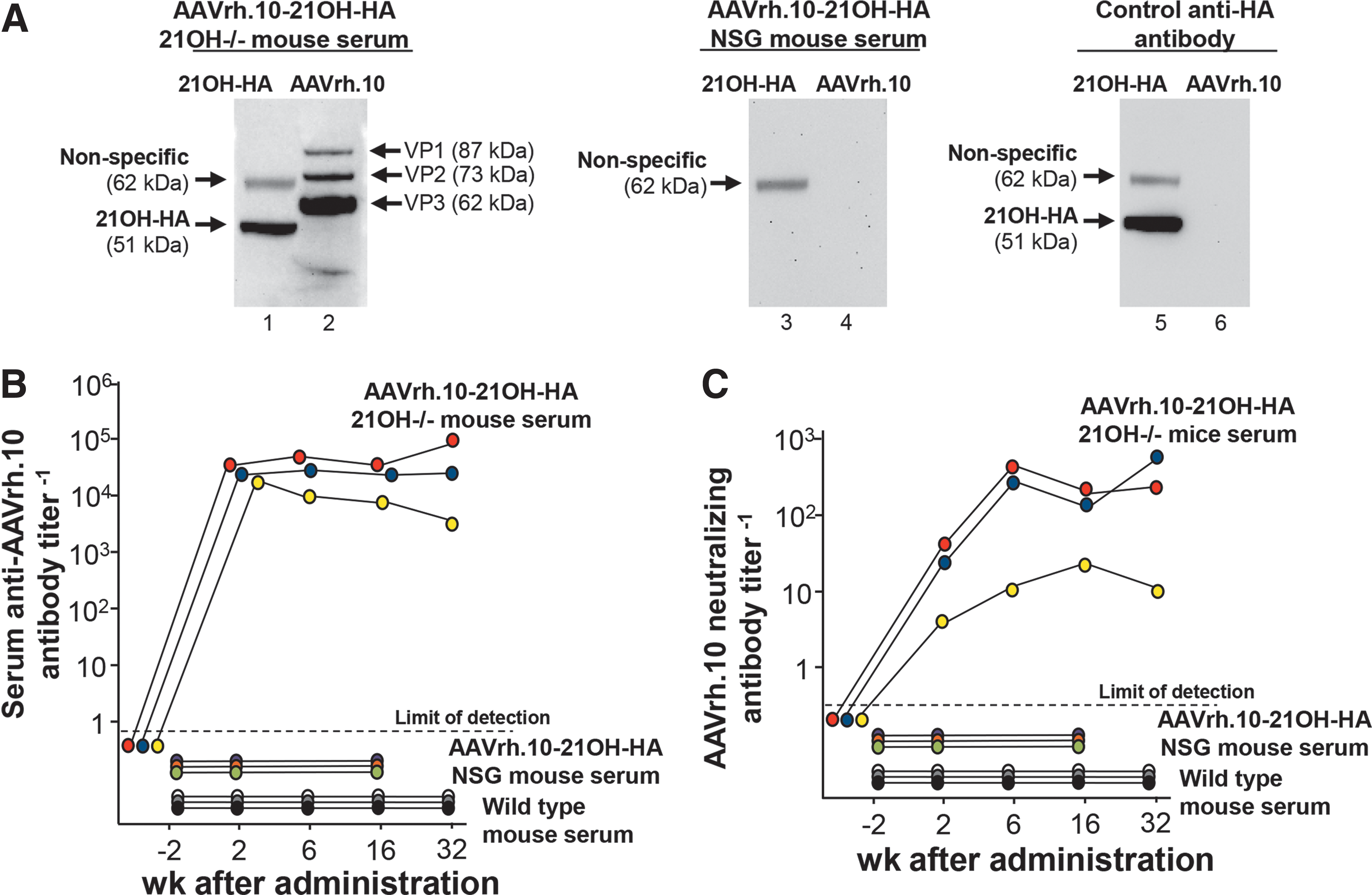
Lack of impact of anti-vector or anti-transgene immunity on transgene expression.
Discussion
Previous reports have shown the efficient transduction of adrenocortical cells by AAV vectors, leading to high levels of transgene expression and short term therapeutic efficacy. Based on these results, we and others have proposed AAV-based gene therapy to treat CAH. 18 –20 However, the biology of the adrenocortical cell turnover may prevent AAV-mediated long-term gene therapy of diseases involving genetic defects in enzymes involved in steroid biosynthesis. Using a murine model of 21OH deficiency as the prototype adrenocortical disease, we have investigated persistence of AAVrh.10 vector mediated 21OH-HA expression and efficacy in 21OH deficient (21OH−/−) mice. The data demonstrate that a single intravenous administration of AAVrh.10-21OH-HA efficiently transduces adrenocortical cells leading to high levels of 21OH-HA expression and restoration of normal steroidogenesis. Progesterone and ACTH normalized to wild-type levels within 2–8 weeks after AAV administration. However, transgene expression and therapeutic efficacy rapidly declined, with complete loss of the therapeutic efficacy by 10 weeks. This loss of efficacy was not due to immune response against the vector and/or transgene, since parallel studies in immunodeficient NSG mice showed the same decline in transgene expression. These observations are consistent with pulse radiolabeling experiments in mice demonstrating that in a time period of 8 weeks, all zona glomerulosa cells and half of the zona fasciculata cells are known to be replaced due to cellular turnover in mice. 21 Based on these data, we concluded that while AAV vectors are effective in the short term, AAV vectors are not suitable for persistent adrenal gland cortex genetic modification, and will not be useful for hereditary disorders such as CAH.
Transduction of adrenocortical zone by AAVrh.10-21OH-HA
The adrenal gland is comprised of two distinct layers: the outer cortex and the inner medulla. The cortex has three zones—the outer zona glomerulosa, the inner zona fasciculata, and the innermost zona reticularis—that together produce endogenous mineralocorticoids, glucocorticoids, and androgens. 34 –37 In mice, the adrenal gland only expresses mineralocorticoids due to the constitutive absence of 17α-hydroxylase in that organ. 34,38 Our study focused on AAV-mediated delivery of the 21OH coding sequence to the zona fasciculata and X-zone (the anatomical term of the zona reticularis in mice) 34,39 Previous studies have shown that murine 21OH is normally expressed in the zona fasciculata. 24,36 As shown in the immunohistology, AAVrh.10-21OH-HA has tropism for this region and can provide a functional mineralocorticoid pathway that was able to reduce progesterone levels to normal in the short term in 21OH−/− mice. The cells of the three layers express specific subsets of cell surface receptors, which are up- and down-regulated as the cells migrate centripetally toward the medulla. 22 –28,40,41 In the mouse, expression of the cell surface receptor for the binding of AAVrh.10 is likely highest in the zona fasciculata and X-zone.
Adrenocortical cells are continuously renewed throughout life. In mice, pulse radiolabeling of adrenal cortex cells has shown that all cortical cells are replenished within 200 days. 21 The stem cell population is located at the outermost rim at the adrenal capsular/subcapsular region. 42 These cells give rise to adrenocortical precursor cells, which differentiate to glomerulosa cells. The differentiated glomerulosa cells undergo lineage conversion to fasciculata cells and migrate centripetally until they reach the juxtamedullary region where they undergo apoptosis. 22 –25,27,28 Expression of 21OH-HA after AAVrh.10-21OH-HA administration was restricted to the zona fasciculata and X-zone, and the turnover of the cells in the zona fasciculata occurred within 70 days after vector administration, with concomitant loss of expression of 21OH-HA.
Loss of transgene expression due to cortex cell turnover in immune deficient mice
AAV-mediated transgene expression can be lost due to humoral and cellular immune responses against the vector and/or transgene. 43 –45 To analyze whether loss of 21OH-HA expression was due to an immune response, we assessed 21OH-HA gene expression in AAVrh.10-21OH-HA-treated immunodeficient NSG mice. Analysis of serum from AAVrh.10-21OH-HA treated 21OH−/− mice, identified antibodies against the 21OH-HA protein and the capsid proteins (VP1, VP2, VP3) of the vector, whereas these antibodies could not be detected in AAVrh.10-21OH-HA treated NSG mice. Similar waning of transgene expression in the adrenal gland was observed in wild-type and NSG mice, consistent with the conclusion that immune responses against the transgene or the vector were not involved.
Together, the data in the present study indicate that the loss of AAV-mediated transgene expression and therapeutic efficacy is most likely due to the turnover of adrenocortical cells. Thus, while AAV vectors can efficiently transfer genes to the adrenal cortex, the biology of the adrenal gland with its persistent turnover of cortical cells result in ineffective correction of hereditary adrenal cortical disorder on a persistent basis using these vectors. Since the turnover of the adrenocortical cells is derived from cortical stem cells in the outer rim of the cortex, long-term correction of hereditary adrenal cortical diseases, such as CAH, will likely require vectors that insert the corrective transgene into the genome of the cortical stem cells.
Footnotes
Acknowledgments
We thank T. Shiroishi, Institute of Genetics, Japan for access to the 21OH−/− mouse model and P. Aubourg and P. Bougnères (Bicetre Hospital, Le Kremlin Bicetre, Paris, France) for helpful discussions. These studies were supported, in part, by the Department of Genetic Medicine, Weill Cornell Medicine (New York). O.P. was supported, in part, by the Parker B. Francis Foundation.
This study was designed by S.M., B.P.D., J.B.R., and R.G.C. and supervised and coordinated by S.M.K., D.S., O.P. and R.G.C. Experiments were performed by S.M., B.P.D., J.R., and C.L.J.; data analysis was conducted by S.M., B.P.D., J.B.R., and P.L.L. The manuscript was written by S.M., B.P.D., O.P., and R.G.C. and edited for content by S.M., B.P.D., and R.G.C.
Author Disclosure
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.