
Abstract
Recombinant adeno-associated virus vectors (rAAVs) represent a reliable tool for basic and translational research, while rAAVs are also making strides in early clinical trials as vehicles for gene transfer. Their low immunogenicity, tissue tropism, and relative safety due to their low rate of genomic integration represent key features, making rAAVs promising instruments as vectors for future gene therapy approaches. Specifically, for cardiovascular gene therapy, rAAVs appear superior to other vector systems such as lenti- and adenoviral vectors due to the ease of accomplishing long-term cardiac expression of target genes and the reduced risk of provoking immune responses or triggering malignancies through genomic integration. However, major obstacles remain to be resolved if rAAVs are to achieve their full potential as gene therapy vectors in clinical trials. The main hurdles prohibiting their sustained success are their limited capacity to carry transgenes of larger sizes, the prevalence of neutralizing antibodies in the general population, and their tissue specificity, which leaves room for improvement. This review discusses the properties of rAAV that make them useful tools in experimental studies and the treatment of cardiovascular disease in patients.
Introduction
C
Gene therapy approaches for CVDs are currently under investigation in preclinical animal models and in clinical trials. In gene therapy, genetic material is introduced into an organism with the aim of regulating protein expression ultimately to improve the function of a target organ. Available vectors used to carry genetic information display dramatic differences in the efficacy of transgene expression, the targeting of the desired tissue/cells, and the duration of expression.
Initially, gene transfer was achieved by using naked plasmid DNA. In animal models, this approach was suitable for the transient cardiac expression of reporter genes, albeit with low transfection efficacy, 4 which can be somewhat enhanced by incorporating DNA into liposomes. 5 Lasting and efficient transgene expression, however, was only achieved by the use of viral vectors. Today, integrating γ-retroviruses and lentiviruses as well as non-integrating adenoviruses (AdVs) and adeno-associated virus vectors (AAV) are in frequent use in preclinical animal models and clinical trials.
While retro- and lentiviral vectors have cemented their place in basic research and are making large strides in first clinical applications, integrating viruses have prompted multiple reports of secondary malignancies after virus-based gene therapy. 6,7 AdVs, on the other hand, are of limited clinical use due to their high immunogenicity and only short-term transgene expression. Therefore, AdVs are mostly used to boost vaccination efficacy by administration of AdVs expressing antigens specific to a variety of infectious disease, such as malaria, rabies, and hepatitis, 8 –10 and in cancer therapy as oncolytic agents. 11
Gene therapy of heart failure specifically demands an efficient transduction of cardiac tissue, a low immunogenicity, and a reliable long-term expression of a therapeutic transgene. For this reason, recombinant AAVs (rAAVs) represent a promising treatment option for gene therapy, albeit with specific pitfalls of their own. This review highlights key features of AAV biology, their use in preclinical animal models, their clinical application, and their potential further use in the treatment of CVD.
Wild-Type Aav
Wild-type (wt) AAV, first discovered in 1965 as contaminations in AdV preparations, are single-strand DNA viruses of the family of parvoviridae, with a size of 20 nm and a packaging capacity of 4.6–4.8 kb. 12,13 AAV serotype 2 represents the most studied and best understood serotype of the AAV family. While there are slight differences between serotypes, the basic structure remains the same, which is why here the fundamental characteristics of AAVs are described on the basis of AAV2. AAV2 consists of the viral genome embedded in a capsid consisting of the three Cap-Proteins VP1, VP2, and VP3, which organize into an icosahedral structure with a mixture of the three proteins at a ratio of 1:1:10. 14 The entry of wtAAV2 into cells is dependent on chlathrin-mediated endocytosis, 15 followed by the formation of stable double-strand viral genomes, which is organized in circular episomes or higher-order concatemers 16 for the most part, although genomic integration of the viral genome is reported with varying frequencies. Genomic integration of AAV2 has been reported at a frequency of 60–70%, with a preferred integration site termed “adeno-associated virus integration site 1” at q13.4-ter of chromosome 19. 17 –19 The viral genome of AAV2 consists of three features: two open reading frames for replication proteins (Rep) and the Capsid gene (Cap) together with inverted terminal repeats (ITRs), which form hairpin structures required for replication and packaging of the viral genome. 20 The three VP proteins are generated from the Cap gene via alternative splicing with the p40 region acting as the promoter, while the Rep gene is transcribed into the four replication regulating proteins Rep78, Rep68, Rep52, and Rep40 via the promoter regions p5 and p19 (Fig. 1A). Recently, an eight protein generated from the AAV2 genome has been identified and termed assembly-activating protein (AAP). This protein, with a calculated molecular weight of 23 kDa, is translated from the Cap gene via a nonconventional translation initiation site and has been demonstrated to act as a chaperon regulating capsid assembly by directing the capsid component proteins to the nucleolus of the host cell where capsid formation occurs. 21 In most AAV serotypes, AAP-mediated capsid assembly is essential for the generation of functional viral particles, with the absence of AAP leading to a proteasomal degradation of capsid proteins, while in others—particularly AAV4, AAV5, and AAV11—a baseline viral particle assembly remains in the absence of AAP. 22,23 Apart from the better studied serotypes (such as AAV2), a variety of serotypes exist, identified in both humans as well as nonhuman primates, each with a unique tissue tropism. For example, AAV2 and AAV9 display a rather diverse infection pattern, 24 while AAV1 favors skeletal muscle infection, and AAV5 shows high efficacy in infecting neurons. 25,26
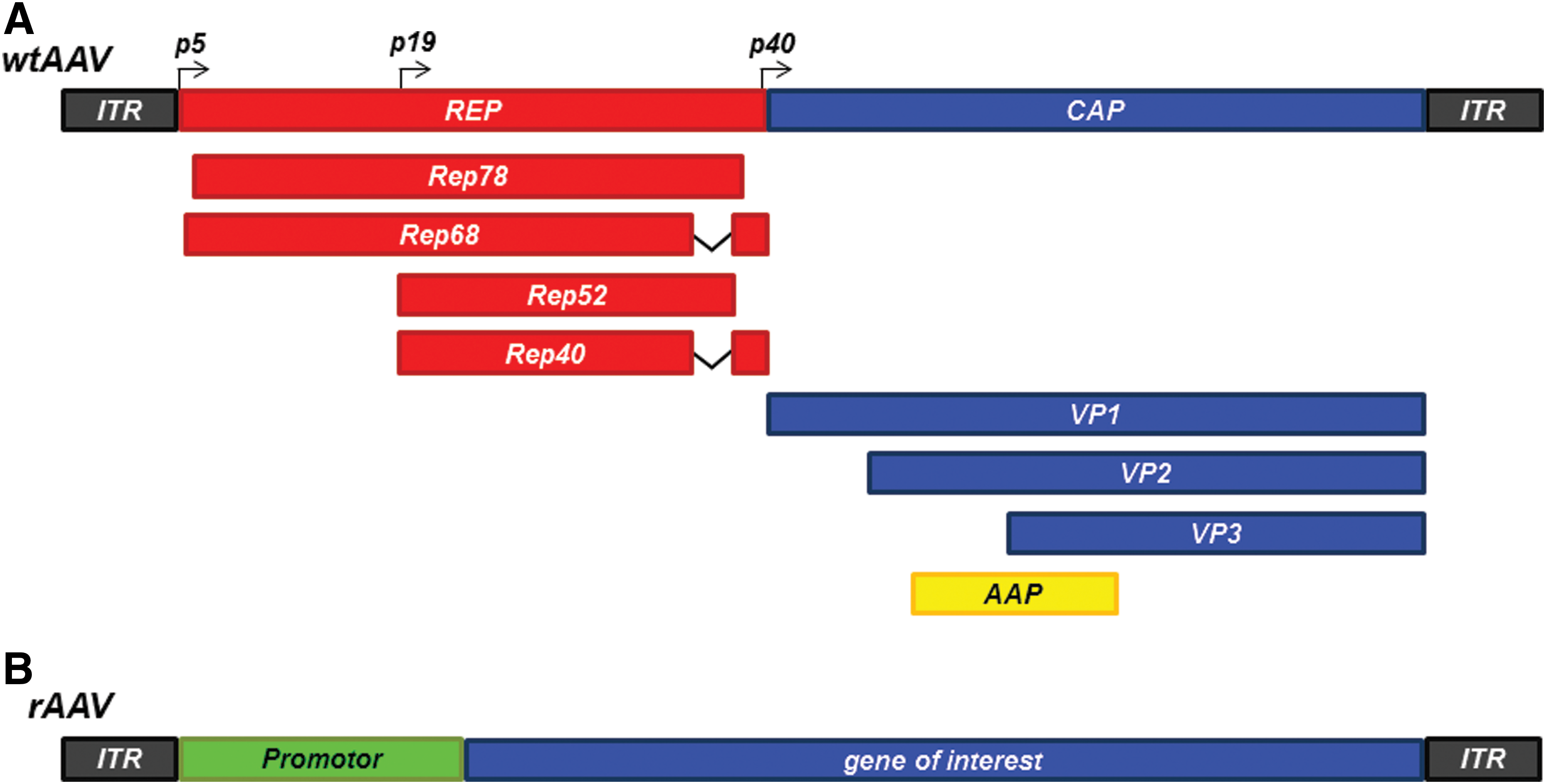
rAAV
Modifications to the wtAAVs result in the generation of rAAV. Since the viral genome (apart from the inverted terminal repeats) is dispensable for transduction and intracellular genome organization, the Rep and Cap sequences can be removed from the viral backbone, leaving room for promoter regions and genes of interest, with a packaging capacity of 4.5–5 kb (Fig. 1B). Furthermore, by dispensing with the Rep ORF present in wtAAVs, chromosomal integration is drastically reduced to a rate of 0.01–0.1%. 27
Another modification to rAAVs in contrast to their wtAAV counterparts is achieved by pseudopackaging. In pseudopackaging, a gene of interest is inserted in a viral genome of one serotype void of the Rep and Cap sequences (leaving only the viral ITRs) in combination with the capsid proteins of another, leading to the creation of novel sub-serotypes with favorable tropisms and expression profiles. Generally, rAAVs are produced by triple transfecting mammalian cells (usually HEK293 cells) with a plasmid carrying the gene of interest flanked by serotype specific ITRs, together with a plasmid encoding the capsid proteins and an adenoviral helper plasmid. The generated viral particles are then quantified via quantitative polymerase chain reaction (PCR) after DNA digestion to quantify the amount of intact viral vectors, whose DNA content is shielded from digestion via the capsid protein (termed DNAse resistant particles [DRP]). From the initially described AAV2/1 (an AAV2 ITR containing backbone in a AAV1 virion), which displays an improved myocardial expression profile, 28 multiple combinations of the AAV2 backbone with capsids from other serotypes have been produced, the tropisms of which have been extensively studied by Zincarelli et al. 29 From those, AAV2/5, AAV2/9, and AAV2/10 in particular have demonstrated a robust and long-lasting cardiac expression. 30,31 Apart from the recombination of capsids and viral genomes from various subtypes, additional modifications of AAVs are devised to improve transduction efficacy and transgene expression. One such example lies in the construction of diverse AAV libraries via error-prone PCR-mediated directed evolution or DNA shuffling, which in the past lead to the generation of the AAV strain AAVM41 that displayed a high cardiac muscle transduction capability while reducing off-target transduction specifically of the liver. 32,33 Further improvements in tissue tropism were also achieved by adding targeting peptides to the capsid structure for improved endothelial transduction efficacy. 34 This is of particular interest, since vascular diseases represent a main driver of cardiovascular mortality, and unmodified rAAV serotypes have proven unfit to target endothelial cells thus far.
AAVs in Preclinical Animal Models of CVD
Animal models of CVD are well established and can mimic central human disease such as arterial hypertension, atherosclerosis, hypertrophic cardiomyopathy, and chronic and acute myocardial ischemia. Furthermore, AAVs have been in frequent use in animal models, helping to elucidate pathophysiological processes and testing therapeutic strategies in translational animal models. Along with the variety of animal models, an abundance of administration routes of rAAVs have been conceived with variable degrees of efficacy.
Since rAAVs are well suited to target cardiomyocytes, 35,36 models of heart failure and myocardial ischemia represent the bulk of rAAV applications in preclinical animal models. Those studies can be grouped by mechanisms that are attempted to be improved. On the one hand, rAAVs can be used to improve cardiomyocyte function by altering the expression of key functional proteins. On the other hand, rAAVs may serve to induce overexpression of secreted factors mediating angiogenesis, arteriogenesis, or attenuating inflammation, in short by regulating vascular physiology.
One key feature in the failing heart is found in the dysregulation of Ca2+ cycling in cardiomyocytes, which leads to a loss in contractility. Here, failing cardiomyocytes show a reduction in Ca2+ transient amplitudes, which are generated by calcium transporting proteins, such as sarcoplasmic reticulum Ca2+-ATPase (SERCA2a), a pivotal protein in calcium handling in cardiomyocytes and ryanodine receptor (RyR2), which releases Ca2+ from the SR into the cytosol. 37 A regulatory protein central to the modulation of Ca2+ currents is S100A1, which mediates its action via regulating, among other proteins, SERCA2a and RyR2, 38 rendering S100A1 a worthwhile target in cardiac gene therapy. To this end, Pleger et al. transduced wt pigs with rAAV9-S100A1 after intermittent occlusion of the left circumflex artery, followed by an observation period of up to 14 weeks. 39 A retrograde vector delivery route via the anterior cardiac vein was chosen in this study, which allowed for robust myocardial expression, as demonstrated in their publication, and is especially useful for reducing the amount of viral vector needed as well as off-target tissue transduction. The antegrade infusion of viral vectors represents an alternative method of delivery. However, pressure-regulated retroinfusion has been proven to be of wider use in cardiac gene therapy in pigs, since it can be performed in porcine models of chronic myocardial ischemia where the antegrade delivery route is blocked by the permanent occlusion of the coronary artery supporting the ischemic area. 40 –43 In the case of retrograde S100A1 delivery, the drop in S100A1 expression seen in the ischemic area of the anterior wall could be ameliorated, leading to an increase in Ca2+ transient amplitude and sarcoplasmatic Ca2+ uptake and a decrease in diastolic Ca2+ levels and sarcoplasmatic Ca2+ leakage via a normalization in SERCA2a and RyR2 levels, ultimately improving cardiac function in the failing myocardium. Attempts have also been made in regulating the expression of SERCA2a itself via AAV systems by antegrade injection of rAAV1.SERCA2a (with saline as control) in a pig model of volume overload heart failure, which was achieved by rupturing the mitral valve chordae tendinae utilizing a catheter-based biopsy pincer. The overexpression of SERCA2a achieved reduced left ventricular diameters, increased ejection fraction, and increased left ventricular contractility (dP/dTmax) 2 months after gene transfer (4 months after induction of mitral regurgitation). 44 This last study, together with the work done on SERCA2a in rodents and in vitro, 45 –47 is of particular interest, since the positive results of these studies led to the initiation of the first-in-human clinical trials to utilize rAAV-based gene therapy in heart failure (the results of which are discussed in the next section).
Although major strides in the development of treatment strategies for heart failure have been made in recent years, patients with terminal heart failure still rely on heart transplantation as a final treatment option, a procedure evoking a whole new set of complications that might be targeted by AAV-based gene therapy. While acute organ rejection is manageable via immunosuppressive treatment regimes, chronic vascular rejection remains a common issue limiting graft and patient survival. 48 –50 In preclinical pig models of heart transplantation, Postrach et al. demonstrated that rAAV2/9-mediated myocardial overexpression of Thymosin β4, an anti-apoptotic and anti-inflammatory peptide, 51 can reduce vascular inflammation and promote graft capillarization in two porcine models of heterotopic heart transplantation. 52 One especially interesting fact in this study is the administration route of the rAAV. Here, rAAV2/9.Tβ4 or the control vector is administered via the perfusion of the donor organ in situ via Langendorff perfusion, a method that might drastically improve myocardial targeting while avoiding the host immune response to the viral particles. Thus, transduced donor organs revealed a decrease in necrosis, apoptosis, and vascular inflammation while improving regional myocardial function of the graft, highlighting an interesting application for rAAVs, especially considering the need for optimal graft performance in times of limited donor organ supply.
Apart from focusing on the improvement of myocardial function and the amelioration of arrhythmias, other approaches tackle coronary artery occlusion and chronically insufficient blood supply to the myocardium, which is the cause for myocardial deterioration in a large subset of heart failure cases. This chronic state of ischemia can trigger a metabolic switch in cardiomyocytes, leading to hibernating myocardium, a disease state that can be reproduced in animal models as well. In one such study, a reduction stent was implanted into the circumflex artery of pigs, leading to a complete occlusion of the vessel over a period of 4 weeks. This slow closure of the vessel induces the conversion of the downstream myocardium into a hibernating state. 53 After the initiation of this hibernating metabolic state, rAAV2/9 encoding for VEGF-A and/or platelet derived growth factor B (PDGF-B) were retrogradely infused. 38 Of note, although rAAV.VEGF-A induced an increase in capillary networks, it lacked the ability to attract pericytes for capillary maturation as well as the induction of collateral growth. As a novel angiogenic agent, the transcription factor MRTF-A was injected via pressure regulated retroinfusion into the great cardiac vein. MRTF-A was shown to regulate SRF target gene expression, like the proangiogenic CCN1 and CCN2, which may enhance recruitment of mural cells to newly formed endothelial sprouts. 54,55 This treatment resulted in the increase of mature capillaries, collaterals, and, ultimately, myocardial function in pigs. A similar effect is observed in the same pig model when overexpressing Thymosin β4, which drives MRTF-A nuclear translocation by competing with MRTF-A for G-actin binding. 55 Consequently, rAAV2/9.Tβ4 overexpression resulted in an increase in mature capillaries, collaterals, and cardiac function. 56
Protection of microvascular integrity is of utmost relevance not only in chronic ischemic instances as cited above, but also in acute myocardial ischemia. Particularly in scenarios of reopening of the infarct vessel, a standard in current clinical treatment, inflammatory events such as recruitment of neutrophils and pro-inflammatory macrophages in the early phase of post-ischemic sterile inflammation determine the expansion of infarct size. 57 Here, preemptive AAV treatment with antioxidant enzymes such as heme oxygenase 1 (HO-1) was capable of reducing infarct size and functional impairment in the early post-ischemic phase via a reduction in reactive oxygen species production and endothelial adhesion protein upregulation, where rAAV.HO-1 treatment showed similar efficacy compared to genetically modified pigs ubiquitously overexpressing HO-1, further highlighting the potency of rAAV-mediated gene transfer. 58
While the coronary vasculature is already the target of various rAAV-mediated gene therapy approaches, rAAVs capable of specifically transducing endothelial cells are currently missing, leading to difficulties in rAAV-based treatment strategies for systemic vessel disease such as atherosclerosis. One attempt to target a systemic vascular pathology was made in a study from the authors' group where rAAV2/9 was injected into the tail vein to overexpress the pericyte chemoattractant PDGF-B in mice before inducing a systemic inflammation via the intraperitoneal injection of lipopolysaccharides. This treatment led to a marked increase in pericyte recruitment to the microcirculation, which improved vascular barrier function, resulting in an attenuation of sepsis-induced systemic hypotension. This stabilization of vascular function was able to reduce lipopolysaccharide-associated mortality in rAAV.PDGF-B-treated mice as compared to rAAV.LacZ-treated control animals, 36 a series of effects that could later be replicated by the transduction of Thymosin β4 via a rAAV2/9. 59 While these interventions targeted the vascular system, the therapeutic effect was mediated by a secretable factor, which explains the efficiency in the modification of vascular biology without a meaningful transduction of vessel structures observed in this study.
Apart from the overexpression of individual important regulatory proteins or growth factors during heart failure, rAAVs have also been used to alter disease-related expression patterns via the overexpression of microRNAs (miRNAs). miRNAs regulate gene expression by binding to multiple mRNAs of target proteins, resulting in mRNA degradation or repression of mRNA translation, thus regulating whole cellular pathways instead of individual proteins. 60 Among a host of miRNAs dysregulated in the failing heart, miR-1—which suppresses pro-hypertrophic pathways via a downregulation of insulin-like growth factor-1, calmodulin, and MEF2A 61,62 —is among the earliest downregulated miRNAs during heart failure, 63 making it a worthwhile target in the treatment of cardiac hypertrophy. Indeed, restoration of miR-1 expression via AAV9 in rats undergoing ascending aortic stenosis-induced heart failure not only ameliorated miR-1 downregulation, but also drastically reduced myocardial hypertrophy and improved myocardial function. 64 Studies targeting microRNAs might be of particular interest, since heart failure, unlike monogenic disease discussed in the next section, is a multifactorial entity that might be hard to treat by regulating expression of individual proteins and rather responds to the regulation of whole signaling networks, which might be facilitated by rAAV-mediated overexpression of miRNAs.
This short insight into the use of rAAVs in preclinical animal models highlights the variability in the use of rAAVs. The work done in pigs in particular represents a close approximation of human disease states, which highlights the possibility of rAAV application in patients. In the next section, advances in rAAV-based gene transfer in clinical trials in general and in heart failure patients in particular will be discussed.
rAAVs in Clinical Trials
Initial clinical trials with rAAVs started in the 1990s, with work done in 12 patients suffering from cystic fibrosis, a disease elicited by defects in the cystic fibrosis transmembrane conductance regulator protein (CFTR) leading to increased viscosity of epithelially secreted mucus. 65 The initial test subjects received an AAV2-based rAAV carrying the complete human CFTR ORF via local administration (nasally and instillation into the right lower pulmonary lobe). 66 Since this initial treatment was tolerated well and target organ transgene expression was demonstrated, the treatment strategy was pursued through various clinical trials up to a Phase IIB clinical trial in 2007, in which aerosolized AAV-CFTR was administered repeatedly in >100 subjects. 67 Unfortunately, a significant improvement of pulmonary function did not materialize in this line of clinical studies.
Since these early attempts in human rAAV-based gene therapy, a plethora of clinical trials involving rAAVs—more than 180 to date
68
—have been conducted or are ongoing. While rAAVs, with a total of 183 reported trials, historically represent the fourth most used vector system—after adenoviral vectors, naked/plasmid DNA, and retroviral vectors—they have become the most widely used vector system in current clinical trials. Of all gene therapy approaches registered on
While rAAVs have been in use in clinical trials for approximately 25 years, they have yet to demonstrate their potency in the treatment of CVD. The initial calcium upregulation by percutaneous administration of gene therapy in patients with cardiac disease (CUPID) trial and the variations of this pioneering work represent the first-in-human approach to treat a CVD utilizing rAAVs as gene delivery systems. This study, delivering Mydicar (AAV1.SERCA2a) in patients with heart failure, demonstrates the feasibility of rAAV gene therapy in CVD as well as its limitations.
The CUPID trials are, apart from being the first-in-human rAAV approaches, the only gene therapy program that have tested rAAVs to treat heart failure. The vector of choice was AAV1 carrying the SERCA2a gene. The Phase I trial started in May 2007, after successful demonstration of improved LV systolic function in a pig model of volume overload heart failure.
44
Initially, nine patients with systolic heart failure (NYHA class III/IV) were enrolled in a dose escalating study applying 1.4 × 1011–3 × 1012 DRP of the rAAV in an open-label manner. An additional group of three patients receiving 1 × 1013 DRP was later added, and these patients fared well, establishing the safety of this vector dose in human application with potential therapeutic efficacy.
75
It is noteworthy that two patients who failed to improve clinically had higher titers of neutralizing antibody compared to other patients, and no vector genome was detected in the cardiac tissue from these patients at the time of their clinical events. These results became the basis for exclusion criteria (neutralizing antibody positive) in following Phase IIa and IIb studies. In the ensuing Phase IIa study, 39 patients were randomized to receive either low-dose 6 × 1011 DRP, middle-dose 3 × 1012 DRP, high-dose 1 × 1013 DRP, or placebo, at a ratio of 8:8:8:13. This double-blind, randomized, placebo-controlled study demonstrated significant improvement in pre-specified composite endpoints in patients treated with high-dose AAV1.SERCA2a at 12 months after gene therapy.
76
Longer-term follow-up data for these patients were reported later, showing a reduced risk of cardiovascular events by 82% in patients after high-dose AAV1.SERCA2a treatment versus the placebo group.
77
The Phase IIb, multicenter, international, double-blind, placebo-controlled study enrolling a total of 250 patients was subsequently initiated to confirm these exciting data in a broader cohort. In this study, NYHA class II patients were also included, and they received either placebo or 1 × 1013 DRP of AAV1.SERCA2a at a ratio of 1:1. The primary endpoint was time to recurrent clinical events after gene therapy. Despite the enthusiasm raised by the results of previous trials, this Phase IIb trial failed to demonstrate reduction in clinical events after AAV1.SERCA2a gene therapy compared to the placebo treatment (104 vs. 128 events; hazard ratio = 0.93, 95% confidence interval 0.53–1.65; p = 0 · 81). Another two AAV1.SERCA2a trials that ran in parallel, one in the United Kingdom (SERCA-LVAD
A variety of reasons can be considered for the lack of efficacy in the CUPID Phase IIb trial. It is likely that one or more of the following issues might have negatively affected rAAV1.SERCA2a potency: vector serotype (AAV1), vector dose, vector quality, delivery method (intracoronary slow infusion), target gene (SERCA2a), or promoter. It is also possible that the targeted patient population was not suited for testing this therapy for the following reasons: patients did not have reduced SERCA2a expression before treatment, comorbidities decreased the efficacy, or the amount of fibrosis in the heart was already too advanced to improve clinical outcomes. The clinical trial design, including endpoints and study length, may also have contributed to the ultimately disappointing trial outcome. The difficulty in clearly identifying the errors in the study design that are responsible for the inefficiency of the Phase IIb trials highlights the challenges in rAAV-mediated cardiac gene therapy that the field currently faces. However, available evidence from cardiac tissue analysis in patients receiving AAV1.SERCA2a from the Phase IIb trial suggests that low gene transduction is at least one of the key problems. This might be due to the host immune response to AAV particles upon intravenous or intracoronary rAAV administration. While successful gene therapy approaches currently rely heavily on local injection of viral vectors (e.g., in the case of Parkinson's or various retinal disease), local viral vector administration seems an unlikely strategy for the treatment of heart failure, which necessitates more scrutiny in terms of dealing with host immunogenicity for cardiovascular applications of rAAVs. Therefore, improvements in vector and/or delivery methods are likely required for future cardiac gene therapy trials. Potential options include employing different vector serotypes, increasing the dose, and modifying vector delivery methods. Recent large-animal cardiac gene therapy studies demonstrate that changes in these factors can increase transgene expression, prompting clinical examination. It remains unclear if these modifications are sufficient to improve transgene expression in humans, but these incremental attempts might promote the field's maturation and lay the groundwork for successful cardiac gene therapy in the future. In the next section, specific strategies to improve gene delivery and reduce rAAV immunogenicity are discussed.
Future Perspectives
Considering that a major breakthrough in AAV-based gene therapy for CVD has thus far failed to materialize, further optimization of rAAVs appears essential, with several relevant parameters in AAV biology having the potential for improvement. On the one hand, the generation of novel vector systems by directed evolution and DNA shuffling represents a promising strategy to generate more tissue-specific virus subtypes facilitating successful transgene delivery, reducing off-target organ transduction, and, potentially, facilitating the transduction of currently insufficiently transducible cell types, such as endothelial cells. 32,33 In order to evaluate these newly designed rAAVs and choose the most promising serotype, monitoring tissue distribution and transduction efficacy is of paramount importance. A useful tool in this endeavor is “Tomato” animal models that have proven capable in registering virus tropism. First generated as a mouse line (mTmG), these animals express a membrane-targeted tandem dimer Tomato (mT) under the control of a chicken β-actin core promoter with a CMV enhancer (pCA), which is surrounded by loxP-sites. After Cre-mediated excision of the mT-sequence, the pCA promoter drives expression of a membrane-targeted enhanced green fluorescent protein (mG). 79 In these animals, newly generated rAAVs carrying a Cre-recombinase can be tested for transduction efficacy and target tissue specificity. More recently, a first Tomato-Pig was described, with comparable features to the mTmG mouse line, which greatly enhances the possibility of observing tissue distribution of viral vectors in this large-animal model, 80 thus expanding the ability to test virus distribution in large-animal models.
Furthermore, optimization of rAAVs is necessary to improve immunogenicity of rAAVs in humans. Although rAAVs induce only a marginal cellular immune response, 81 humoral immune responses, specifically pre-existing neutralizing antibodies, represent a major challenge in the clinical use of rAAVs. 82 Here, the prevalence worldwide lies in the range of 20–30%, with sometimes dramatic differences between AAV serotypes and the prevalence in certain global regions. To this end, neutralizing antibodies against AAV2 have been shown to be the most common, with AAV1, AAV7, and AAV8 antibodies displaying a lower rate of abundance. Interestingly, the population of the United States has the lowest prevalence of neutralizing antibodies, while African populations seems to be the most burdened by neutralizing antibodies to AAV serotypes. 83 Alongside the modification of the viral backbone and capsid, modifications of the viral particle via targeting peptides and cationic dendrimers might also improve target gene delivery. Promising strategies to generate rAAVs capable of escaping antibody neutralization are currently underway, relying on error-prone PCR (insertion of point mutations into the capsid protein by DNA polymerases defective in proof-reading capability), capsid shuffling (nuclease fragmentation and random realignment of different AAV-serotypes), and the generation of rAAV peptide display libraries, summarized by Buning et al., 84 with the efficiency of these approaches awaiting confirmation.
An additional way of improving the control of transgene expression might lie in the development of inducible vector systems. Especially considering the expression of growth factors, a regulatory promoter might be superior to unregulated promoters. However, currently, no regulated expression systems for AAVs that can be used in humans are available. While doxycycline regulated promoters, for example, have been used in preclinical animal models, 56,85 they are unusable in primates due to their immunogenicity caused by their bacterial origin. 86
Apart from modifying rAAV structure, CRISPR-Cas9-mediated gene therapy offers an exciting new avenue for the use of AAVs. The availability of transgenic mouse lines expressing the endonuclease Cas987 allows for rapid gene knockout via the AAV-based delivery of suitable guide RNAs under the control of a U6 promoter. Assuming sufficient tissue specificity and transduction efficacy, this technique can greatly accelerate the study of tissue-specific gene knockouts. Furthermore, this technique promises to permit in vivo genome editing, possibly in humans. To this end, the whole CRISPR-Cas9 system with the Cas9 endonuclease, the guide RNAs, and their respective promoters would have to be delivered through AAVs, which currently exceeds the packaging limit of rAAVs. One workaround lies in the splitting of the Cas9 protein. Recently, a split Cas9 system was devised based on Intein-mediated trans-splicing. Co-expression of an N-Cas9 (amino acids 2–573) combined with an N-Intein and a C-Cas9 linked to the C-Intein in one cell leads to rapid trans-splicing, resulting in a full-length Cas9 and Intein. 88 This system has already been used in vitro where it demonstrated efficient genome editing in murine cell lines, which represents, presupposing sufficient efficacy in genome editing in vivo, an enticing new area of rAAV-mediated gene therapy. Concrete examples of possible applications can be found in genetic disorders causing cardiomyopathies, such as titin-based dilated cardiomyopathies or Duchenne muscular dystrophy. A variety of point mutations resulting in premature stop codons and expression of truncated protein version have been described in both conditions, and both have been demonstrated curable via exon-skipping. 89,90 An exon-skipping strategy via AAV-mediated delivery of CRISPR/Cas9 to affected tissue (skeletal muscle and/or heart, both tissues easily reachable via AAVs) has the potential to fix these genetic disorders permanently, which today still are associated with severe impairments and high mortality.
Concluding, rAAV represents a serviceable tool in basic and preclinical research and is potentially able to succeed in clinical trials if the pitfalls of this technique can be addressed. Emerging novel approaches described above are certainly promising and could be the game-changer in the near future. Importantly, application of these new technologies, new vector/serotype, higher dosing, and more varied delivery method need to be carefully and thoroughly evaluated for their safety profiles before their use in human subjects. The continued efforts of the community are essential and will promise successful clinical translation of this approach.
Footnotes
Acknowledgments
R.H. and C.K. were supported by the Deutsche Forschungsgemeinschaft, the German Ministry for Research and Education, and the Else-Kröner-Fresenius Stiftung.
Author Disclosure
There are no conflicts of interest.