
Abstract
To study whether ectopic human melanopsin (hMel) in retinal ganglion cells (RGCs) could restore the visual function in end-stage retinal degeneration, AAV2/8-CMV-hMel/FYP was injected into the intravitreal space of Royal College of Surgeons (RCS) rats. It was observed that ectopic hMel/yellow fluorescent protein (YFP) was dominantly expressed in the RGCs of the RCS rat retinae. At 30–45 days after administration of AAV2/8-CMV-hMel/FYP in RCS rats, the flash visual evoked potentials and behavioral results demonstrated that visual function was significantly improved compared to that in the control group, while no improvement in flash electroretinography was observed at this time point. To translate this potential therapeutic approach to the clinic, the safety of viral vectors in the retinae of normal macaques was then studied, and the expression profile of exogenous hMel with/without internal limiting membrane peeling was compared before viral vector administration. The data revealed that there was no significant difference in the number of RGCs containing exogenous hMel/YFP between the two groups. Whole-cell patch-clamp recordings demonstrated that the hMel/YFP-positive RGCs of the macaque retinae reacted to the intense light stimulation, generating inward currents and action potentials. This result confirms that the ectopic hMel expressed in RGCs is functional. Moreover, the introduction of AAV2/8-CMV-hMel/FYP does not cause detectable pathological effects. Thus, this study suggests that AAV2/8-CMV-hMel/FYP administration without internal limiting membrane peeling is safe and feasible for efficient transduction and provides therapeutic benefits to restore the visual function of patients suffering photoreceptor loss.
Introduction
Retinitis pigmentosa (RP) is the most common form of inherited retinal degenerative diseases that lead to progressive visual decline resulting from continuing loss of photoreceptor cells. 1,2 RP has been identified in at least 45 genes related to the phototransduction pathway, and >150 mutations have been reported to date in the opsin gene associated with RP since the Pro23 mutation was reported in 1990. Approximately 100 genes related to RP and up to 3,100 mutations have been reported. 3,4 At present, this type of degenerative retinal disease remains incurable. New drug development, 5 –11 stem-cell transplantation, 12 –14 and gene therapy 15 –20 have been conducted using animal models to find effective therapy. The above-mentioned therapeutic strategies aim to rescue visual function either by correcting the mutated gene or by slowing the degenerative process. The combination of multiple causative genes and a broad range of clinical severity has made diagnosis challenging. 3,21 Therefore, most patients miss the opportunity for early treatment and eventually suffer blindness due to loss of photoreceptor cells. Thus, novel treatments need to be developed to treat patients at the end stage of the disease.
Retinal prostheses 21 –24 can partially restore vision to people who have lost their photoreceptors due to retinal diseases such as RP. Various retinal implants including epiretinal 25 –30 (on the retina), subretinal (behind the retina), 31 –36 and suprachoroidal (between the choroid and the sclera) 37 –39 implants introduce visual information into the retina by electrically stimulating the surviving retinal neurons. Thus, elicited percepts have a rather low resolution and may be suitable for light perception and recognition of simple objects. 40 Given that inner retinal cells maintain their neural circuitry, 41,42 which can still transform the visual signal, although with some remodeling during degeneration, converting the surviving inner retinal cell into photosensitive cells by introducing a photosensitive probe, such as ChR2 43 –47 or melanopsin, 48 –51 has recently been attempted.
ChR2 was first described in green algae. It is a light-gated ion channel that can evoke ultrafast membrane depolarization and mediates action potentials. 52 Pioneering work from Pang's lab demonstrated that ChR2 can restore lost visual function due to degenerative retinal diseases. 43 However, xenotransplantation of ChR2 might cause immune responses, therefore decreasing the therapeutic benefit. Another photosensitive protein, melanopsin (gene OPN4), is intrinsically expressed in a group of retinal ganglion cells (RGCs) and so-called intrinsically photosensitive RGCs (ipRGCs). 53,54 Melanopsin is a G protein. Its signaling cascade is coupled to membrane channels, and opsin photopigments use cis-isoforms of retinaldehyde as a chromophore, 55 –58 so the light-evoked response is slower but more durable than that of ChR2 in the absence of photoreceptor-based input. Some labs, including that of the authors, 49 –52 have demonstrated that transfection of an adeno-associated virus (AAV) mouse melanopsin vector into RGCs of rd1 mice can restore visual function. These are the first data supporting the use of melanopsin in the restoration of visual function. Mouse melanopsin and human melanopsin (hMel) share only approximately 78% identity. 59,60 In addition to light sensitivity and kinetic differences, hMel is a better candidate for clinical applications.
Furthermore, Royal College of Surgeons (RCS) rats are a chronic retinal degeneration model that has a mutation in the Mertk gene expressed in RPE cells, resulting in progressive photoreceptor cell death and loss of electrophysiological response by postnatal day 60 (P60d), 61 which is an ideal model for early-onset retinal disease with slow progression. 62 Thus, this study constructed an AAV2/8-CMV-hMel/FYP vector to express hMel ectopically in the retina and found that it can partially restore visual function in an RP model and is safe in nonhuman primate models. Potentially, hMel is a candidate for a light-sensitive, gene-reformed retinal prosthetic for blindness.
Methods
Experimental design
This was a randomized controlled animal study. Detailed information can be found in Supplementary Tables S1 and S2 regarding the experimental design and workflow for RCS rats and macaques, respectively.
Animals
Sixty healthy SPF-grade RCS rats (irrespective of sex) and three adult macaques (two males and one female Macaca mulatta) were provided by the Experimental Animal Center of the Third Military Medical University. The animals were reared in light-controlled rooms with a fixed lighting schedule (08:00–20:00). The light was generated by two fluorescent lamps with 60 lux of intensity. The room humidity was maintained at 50–60%, while the temperature was maintained at 22–25°C. All experimental protocols followed the Guidance Suggestions for the Care and Use of Laboratory Animals (The Ministry of Science and Technology of the People's Republic of China) 63 and were approved by local ethics committees.
Anesthesia
RCS rats were anesthetized via intramuscular thigh injection with a mixture of haloperidol and dihydroetorphine at 0.1 mL/kg. Oxybuprocaine (0.4%; Santen Pharmaceutical Co. Ltd., Osaka, Japan) eye drops were used for superficial anesthesia. Macaques were anesthetized via intramuscular thigh injection with medetomidine (0.01 mg/kg, Domitor; Orion Corporation, Espoo, Finland) and ketamine (10 mg/kg, Ketalar; Daiichi Sankyo Espha Co. Ltd., Tokyo, Japan) and were then overdosed with sodium pentobarbital of 150 mg/kg intracardially.
Constructs and viral vectors
The pAAV2/8-CMV-hMel/FYP construct and viral vector production
Full-length hMel (GenBank accession no. NM_033282) was cloned into the AAV vector under the transcriptional control of the cytomegalovirus (CMV) promoter (a kind gift from Dr. Bin Lin, Hong Kong Technology University, Hong Kong). The viral vector was produced at Neuron Biotech Co. Ltd. (Shanghai, P.R. China). Two batches of vectors were purified and titrated according to previously published methods. The final concentrations of AAV2/8-CMV-hMel/FYP were 3.52 × 1013 vp/mL and 1.07 × 1013 vp/mL in balanced salt solution. To verify the viral vector before animal administration, HEK293 or cultured hippocampal neurons were infected with AAV2/8-CMV-hMel/FYP and further characterized by immunochemistry and patch-clamp electrophysiology.
Intravitreal injection of the AAV vector
After administration of anesthesia, the RCS rat was moved to an animal operating table under a stereo-surgery microscope on a biological bench under aseptic conditions. Two microliters of the viral suspension (1.07 × 1013 vp/mL) was delivered to the vitreous chamber in the superior hemisphere of the eyes using a Hamilton microinjector. In the behavioral testing, 14 RCS rats received the AAV2/8-CMV-hMel/FYP injection in both eyes in the experimental group, while eight RCS rats received a phosphate-buffered saline (PBS) injection as the control group. However, in the flash electroretinography (fERG) testing, to avoid individual differences, the right eye of each rodent received the AAV2/8-CMV-hMel/FYP injection, and the left eye was used as a control (n = 14).
The six eyes of three normal macaques were divided into two groups: three right eyes as a group were subjected to posterior segment vitrectomy, internal limiting membrane (ILM) peeling, C3F8 gas filling, and intravitreal injection of 30 μL of AAV2/8-CMV-hMel/FYP; three left eyes as a group received only direct intravitreal injection of 30 μL of the same viral vector.
Immunohistochemistry
For fluorescence immunohistochemistry, the whole-mount retinae were blocked in 3% bovine serum albumin (BSA) and 10% normal goat serum in 0.5% Triton X-100 (Sigma–Aldrich, St. Louis, MO) for 1 h at room temperature (25°C). Then, the retinas were incubated with primary and secondary antibodies diluted in PBS with 4% normal goat serum, 1% BSA, and 0.5% Triton X-100. The whole retinae were incubated with a rabbit polyclonal antibody against hMel (1:500; Abcam, Cambridge, United Kingdom) and goat anti-green fluorescent protein (GFP; 1:1,000, Sigma–Aldrich) for 1 day at 4°C and washed in PBS for 45 min (3 × 15 min). Subsequently, the retinae were incubated for 4 h in Cy3-conjugated goat anti-rabbit immunoglobulin G (IgG; (1:1,000; Life Technologies, Carlsbad, CA). The whole retinal tissues were thoroughly washed three times with PBS (3 × 15 min).
The frozen sections were air dried, washed in PBS for 5 min, and then incubated with primary rabbit antibodies against hMel, rhodopsin, and PKCα, and mouse antibodies against recoverin and beta III tubulin in 0.5% Triton X-100 (Sigma–Aldrich), 3% BSA, and 10% normal goat serum (Sigma–Aldrich) in PBS overnight at 4°C. The sections were rinsed three times and incubated with the secondary antibody (Cy3-conjugated goat anti-rabbit IgG 1:1,000; Cy3-conjugated goat anti-mouse IgG 1:1,000; Life Technologies) for 1 h at 37°C.
Imaging and cell counting
All fluorescent images, including the whole-mount retinae and frozen sections, were acquired using a confocal microscopy Zeiss LSM 780 microscope (Carl Zeiss Co. Ltd., Oberkochen, Germany). Offline image processing and analyses were carried out using ImageJ software (National Institutes of Health, Bethesda, MD). To estimate whether the ILM peeling would influence the expression of hMel in the retina of macaques, small areas of 0.09 mm2 were randomly chosen to count the GFP-positive cells. Moreover, the number of GFP-positive cells was averaged from several small areas and converted to cells/mm2 and is presented as the mean ± standard deviation (SD; Supplementary Fig. S2).
fERG recording
Animals were dark adapted for nearly 12 h and prepared for recording under dim red light. After proper anesthesia, the animals' pupils were dilated with tropicamide and phenylephrine drops. fERG responses were acquired from both eyes simultaneously with a contact lens for macaques or gold wire loops for RCS rats. Saline (0.9%) was frequently applied to the cornea to prevent dehydration and allow electrical contact with the recording electrode. Two of the needle electrodes inserted under the skin of angulus oculi temporalis served as the reference electrodes, and the other one placed in the tail served as the ground electrode. The data were acquired under the RETI-scan system (Roland Consult, Brandenburg, Germany).
For the RCS rats, dark-adapted intensity responses ranged from −2.5 to 1 log (cd*s/m2; −2.5, −0.5, −0.02, 0.5, and 1). To avoid adaptation to the previous flash, the flash interval was 60–120 s, depending on the stimulus intensity.
For the macaques, all experimental protocols followed the guidelines of the ISCEV (2015 update), 64 specifying the five standard responses: dark-adapted 0.01ERG (rod ERG); dark-adapted 3ERG (combined rod-cone standard flash ERG); dark-adapted 3 oscillatory potentials; light-adapted 3ERG (cone ERG); and light-adapted 30 Hz flicker ERG.
Flash visual evoked potential recording
The rats were reared in a regular light-controlled room (08:00–20:00). After proper anesthesia, the electrical activity was recorded by the silver wire needle electrodes, which were placed at the visual cortex region, with a reference electrode placed under the chin skin and a ground electrode placed in the tail. The data were acquired using the RETI-scan system (Roland Consult). The stimuli intensity was −0.02 log (cd*s/m2), the frequency was 1 Hz, and the flash duration was <5 ms. The average number of times were >64. The bandpass of the filter was from 0.01 to 300 Hz. The data were exported and processed by Igor Pro (WaveMetrics Inc.).
The macaques were also reared in a regular light-controlled room. After proper anesthesia, the electrical activity was recorded by the silver–silver chloride electrode, which was placed on the scalp over the visual cortex 2–3 cm above the occipital tuberosity, and the reference electrode was placed at the middle of the forehead, while the grounding electrode was attached to the earlobe. The impedance of the electrode was <5 kΩ. When the stimulation of light lasted, the eye was opened by the blepharostat, and the other eye was covered by gauze. All experimental protocols followed the guidelines of the International Society for Clinical Electrophysiology of Vision standard for clinical visual evoked potentials (VEPs; 2015 update). 64
Open field test
The open field test box was 45 cm × 30 cm × 40 cm, divided equally into a white open zone and a dark zone, with a door (10 cm × 10 cm) placed between them. The RCS rats received the intravitreal injection and were raised to 70 days after the test was performed. First, rodents were placed in the darkroom for 2 min for adaptation, and then the door was opened to observe the behavior of the test rats. In the test, the duration in each of the dark and white zones was recorded. The open field test was recorded using a video camera to enable subsequent evaluation under 300 lux of light intensity. 49 The floor was cleaned with 70% ethanol before each animal was tested.
Spatial visual acuity
The Virtual Optomotor System is a type of optometry testing apparatus that is used for detecting animal spatial visual acuity. 65 The apparatus, comprised of two mirrors with a small hole in its center, was placed in the top and bottom of the openings. Four computer monitors projected a virtual cylinder in 3D coordinate space with a vertical sine-wave grating. A platform was passed through the hole in the mirror to the bottom of the apparatus. Animals were placed on the platform, where they tracked the vertical sine-wave grating with head movements. The overhead video camera recorded the head movements. A schematic of the apparatus is shown in Fig. 1. The speed of the grating was 12 deg/s, and the spatial frequency of the grating was six spatial frequencies between 0.05 and 0.6 cpd (0.05, 0.075, 0.1, 0.2, 0.3, 0.4, 0.5, and 0.6 cpd). The acuity threshold was quantified by increasing the spatial frequency of the grating until the following response was lost, thus defining the visual acuity. Rats were tested at 30, 45, and 60 days after surgery.
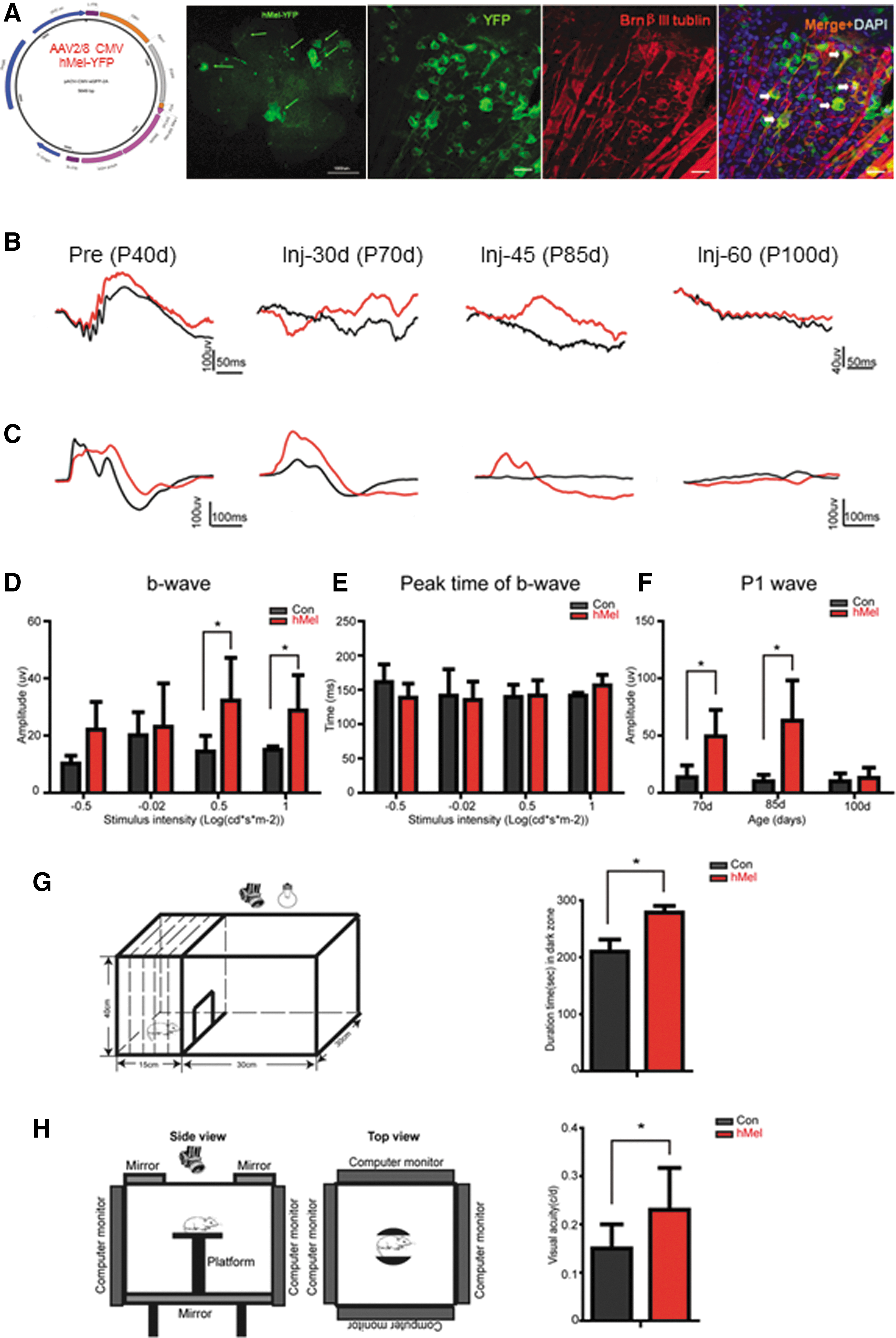
Ectopic expression of human melanopsin (hMel) in retinal ganglion cells restores visual function at the end-stage of disease in Royal College of Surgeons (RCS) rats.
Retina examination
Rhesus macaques underwent assessment by ophthalmologic examination, color retinal fundus photography, autofluorescence (Nonmyd a-D 5MEGA; Kowa, Nagoya, Japan; 488 nm was used to test autofluorescence of the retina, 514 nm was used to observe the thickness of the retina, 795 nm was used to test the autofluorescence of the choroid, 814 nm was used to test the infrared light of the retinal blood vessels), and optical coherence tomography (OCT; Heidelberg Engineering, Heidelberg, Germany) before intravitreal injection and at 1, 2, and 3 months after vector injection.
Patch-clamp recordings
Three months after surgery, the macaques were dark adapted for 2–3 h before the experiments. Then, they were sacrificed by overdosing with sodium pentobarbital at 150 mg/kg intracardially. The eyes were enucleated and placed in carboxygenated (5% CO2 and 95% O2) Ames' medium (Sigma–Aldrich). Then, the cornea and lenses were removed from the eyes, and the retina was separated from the sclera. A piece of the retina was placed photoreceptor-side down in the recording chamber, which was mounted on the stage of a fluorescence microscope (Olympus). Carboxygenated (5% CO2 and 95% O2) Ames' medium (pH 7.4) was continuously perfused into the recording chamber. To block the photoreceptor-mediated light responses, the drug (Tocris Bioscience, Bristol, United Kingdom), including AMPA/kainate receptor antagonist CNQX (25 μM), the mGluR6 receptor agonist L-APB (5 μM), and the NMDA receptor antagonist D-APV (25 μM), were added to the perfusion solution. Yellow fluorescent protein (YFP)-expressing cells were swiftly identified by fluorescence microscopy with blue light. Patch-clamp recordings were performed from the soma of the YFP-positive cells using 4–7 MΩ electrodes, and signals were amplified using Axopatch 700B amplifiers (Molecular Devices, San Jose, CA). The pipette solution for whole-cell voltage-clamp experiments contained (in mM): 118 Cs methanesulphonate, 12 CsCl, 5 EGTA, 0.5 CaCl2, 4 ATP, 0.3 GTP, 10 HEPES, adjusted to pH 7.3 with CsOH. The data were analyzed using Clampfit 10 software.
Pathology
After sacrifice, the heart, liver, spleen, kidney, and brain of the macaques were fixed in buffered formalin. After fixation, the optic nerve, optic tract, and visual cortex were isolated from the brain. All of the tissues listed above were sectioned, processed, and stained with hematoxylin and eosin (H&E).
Statistical analysis
Data analysis was carried out using SPSS for Windows v13.0 (SPSS, Inc., Chicago, IL). The data are expressed as the mean ± SD and were analyzed using independent samples t-tests to compare the treated and control groups in terms of electrophysiology recording and behavioral testing. A p-value of <0.05 was considered statistically significant.
Results
Ectopic expression of hMel in retinal ganglion cells rescues the visual function of RCS rats at the end stage
To assess the ectopic expression and function of hMel in retinal cells, first the expression of hMel in the retinae was examined after injection of the viral vector into the intravitreal space in the eyes of P40d rats. After 30 days of injection, YFP fluorescence was observed in all of the eyes injected with virus (12/12) but not in the retinae of the sham-injected eyes (n = 4) upon embedding the retinae in OCT and cutting them into frozen sections to identify the YFP-positive cells (Fig. 1A). At 30 days after injection, the RGCs expressed hMel (Fig. 1A). Moreover, at this time point, no YFP-positive cells were observed in the inner nuclear layer (INL) or inner plexiform layer (IPL) of the retina of control animal (Supplementary Fig. S1A). Once hMel was expressed in a retinal ganglion cell, the YFP signal was extended over the soma, dendrites, and axon (Fig. 1A, YFP panel). These results indicate that the AAV2/8-CMV-hMel/FYP vector mainly infects RGCs and that intense expression of hMel-YFP driven by the CMV promoter can be achieved in vivo.
At P60d, the RCS rats lost almost all of the rods and cones, thus losing the light response, although the RGCs were maintained. Previous studies confirmed that the b-wave and P1 wave is expected to be abolished after P60d because of the loss of light-evoked synchronous electric signal activities caused by the loss of photoreceptors. The control eyes showed the same tendency (Fig. 1B and C), and the ERG and VEP responses were not evoked by the flash. However, weak ERG and robust VEPs could be evoked in aged RCS rats that were intravitreally administered the AAV2/8-CMV-hMel/FYP vector at 30 days after injection. Additionally, a similar range was observed of the electrophysiological response recorded at 45 days after injection (Fig. 1B and C). While the a-wave was not detectable, a small b-wave was evoked by higher intensity (−0.5 to 1 log [cd*s/m2]; Fig. 1D; p < 0.05, n = 8), but the peak time of the b-wave did not show a difference between the treated group and the control group (Fig. 1E; p < 0.05, n = 8). A broad range of light intensity activated the visual system. The fERG amplitude had a positive relationship, whereas the latency had a negative relationship. At 60 days after injection, the restored response could no longer be detected, while the b- and a-waves were abolished as in the control eyes. A similar tendency was observed in the flash VEP (FVEP) test, but the P1 wave evoked at 30 and 45 days after injection was not detectable at P60d (Fig. 1F; p < 0.05, n = 8).
To determine whether AAV2/8-CMV-hMel/FYP restores the visual function of treated RCS rats, the behavior of each of the experimental animals was tested by using the open field test and the Virtual Optomotor test. In RCS rats, with chronic retinal degeneration, light can no longer be observed after P60d. However, at 30 days after injection of AAV2/8-CMV-hMel/FYP, when the RCS rats were P70d, the treated group stayed in the black zone for a longer time than the control rats did (Fig. 1D; p < 0.05, n = 8). This result suggests that in the treated group, light can be seen as high-intensity white light, and the animals prefer to avoid bright spaces. In the spatial visual acuity test, the rats were placed on the platform for adaptation for 2 min. When a grating was projected on the computer monitor and rotated, the rats usually stopped moving their body and would begin to track the grating. However, tracking in the control RCS rats was poor because of retinal degeneration. Thirty days after injection, the treated rats were placed on the platform, and the spatial frequency of the test grating was incrementally changed. The rats showed the same directional head tracking. The highest spatial frequency that the rats could track was identified as the visual acuity threshold. The treated group reached 0.23 cpd, which is significantly better than that of the control group (0.15 cpd; Fig. 1H; p < 0.05, n = 8).
Characterization of functional ectopic expression of hMel in retinal ganglion cells of macaques through the AAV2/8-CMV-hMel/FYP viral vector
To assess whether surgical ILM peeling increases the expression and of hMel in RGCs, the expression of hMel in retinae with or without surgical ILM peeling was examined at 3 months after intravitreal injection of the viral vector. A strong YFP-positive signal was observed in the somas and adjacent neurites, including dendrites, and axons, in the right and left retinae (Fig. 2A and B), suggesting a sufficient expression of hMel-2A-YFP. hMel antibody fluorescence staining confirmed that YFP-positive cells coexisted with hMel (Fig. 2B). No YFP-positive cells were seen in the INL or IPL of the retina of control animal (Supplementary Fig. S1B). Approximately 10% of the hMel-positive cells were YFP negative, which agrees with the hMel- and YFP-positive axon counting in the optical tract (Supplementary Fig. S1C). These cells may represent the endogenous ipRGC population. The abundant hMel/YFP-positive somas can be seen in the peripheral region of the retinae, the moderately abundant hMel/YFP-positive somas in the equatorial region, and very few hMel/YFP-positive somas but abundant axons in the central region (Fig. 2A). This distribution of hMel/YFP-positive cells was also confirmed by cell counting, as illustrated in Fig. 2C and Supplementary Fig. S2. There was no significant difference between the right eyes (ILM peeling plus intravitreous injection of the viral vector) and left eyes (intravitreous injection of the viral vector without ILM peeling) regarding YFP-positive cells (Fig. 2C; p > 0.05, n = 3).

Characterization of the ectopic expression of hMel/YFP in the macaque retina.
To determine whether ectopic hMel co-expressed with YFP responds to light, hMel-mediated light responses were examined using whole-cell patch-clamp recordings from the retinae of macaques 3 months after injection of the viral vector. To block the photoreceptor-mediated light response, the perfusion buffer contains the AMPA/kainate receptor antagonist CNQX, the NMDA receptor antagonist D-APV, and the mGluR6 receptor agonist L-APB. The recordings were performed in retinal cells with significant co-expression of YFP (Fig. 2D). Figure 2D (left panel) shows that current clamp recordings at 0.08 nA can evoke action potentials. Figure 2D (middle and right panels) shows representative recordings of the inward currents and action potentials evoked by light at a wavelength of 470 nm. Together, these results demonstrate that normal macaque retinal cells can react with light stimulation via the optogenetic method of expressing hMel.
Ectopic expression of hMel in retinal ganglion cells does not alter the ophthalmological morphological and electrophysiological characteristics
To determine whether the surgical peeling of the ILM and intravitreal injection of the AAV2/8-CMV-hMel/FYP vector caused morphological injury to the macaque eyes, the eyes were examined morphologically, including autofluorescence, fundus photography, and OCT, to observe the microstructure of the eyes before and after surgery. Macaque eyes are similar to those of humans. After posterior segment vitrectomy, ILM peeling, C3F8 gas filling, and intravitreal injection of AAV2/8-CMV-hMel/FYP, the macaque's light reflex from the right macula was dispersed, and slight depigment changes were observed, and there were no noticeable abnormal changes in the other segments of right eyes or the whole left eyes. There were no obvious surgical complications, such as hemorrhage, tearing, edema, or retinal detachment (Fig. 3A–D). The macaque eyes are similar to the human eyes through the OCT images. After surgery, the macaques' right eyes had no light reflex of the macular ILM, the thickness of retina was not uniform, and the light reflex of ONL and OLM was not very clear. However, the light reflex of the ellipsoid zone and the tissues out of the ellipsoid zone were clear. After direct intravitreal injection of the AAV2/8-CMV-hMel/FYP vector, the macaques' left eyes had no abnormal light reflex (Fig. 3A–D).
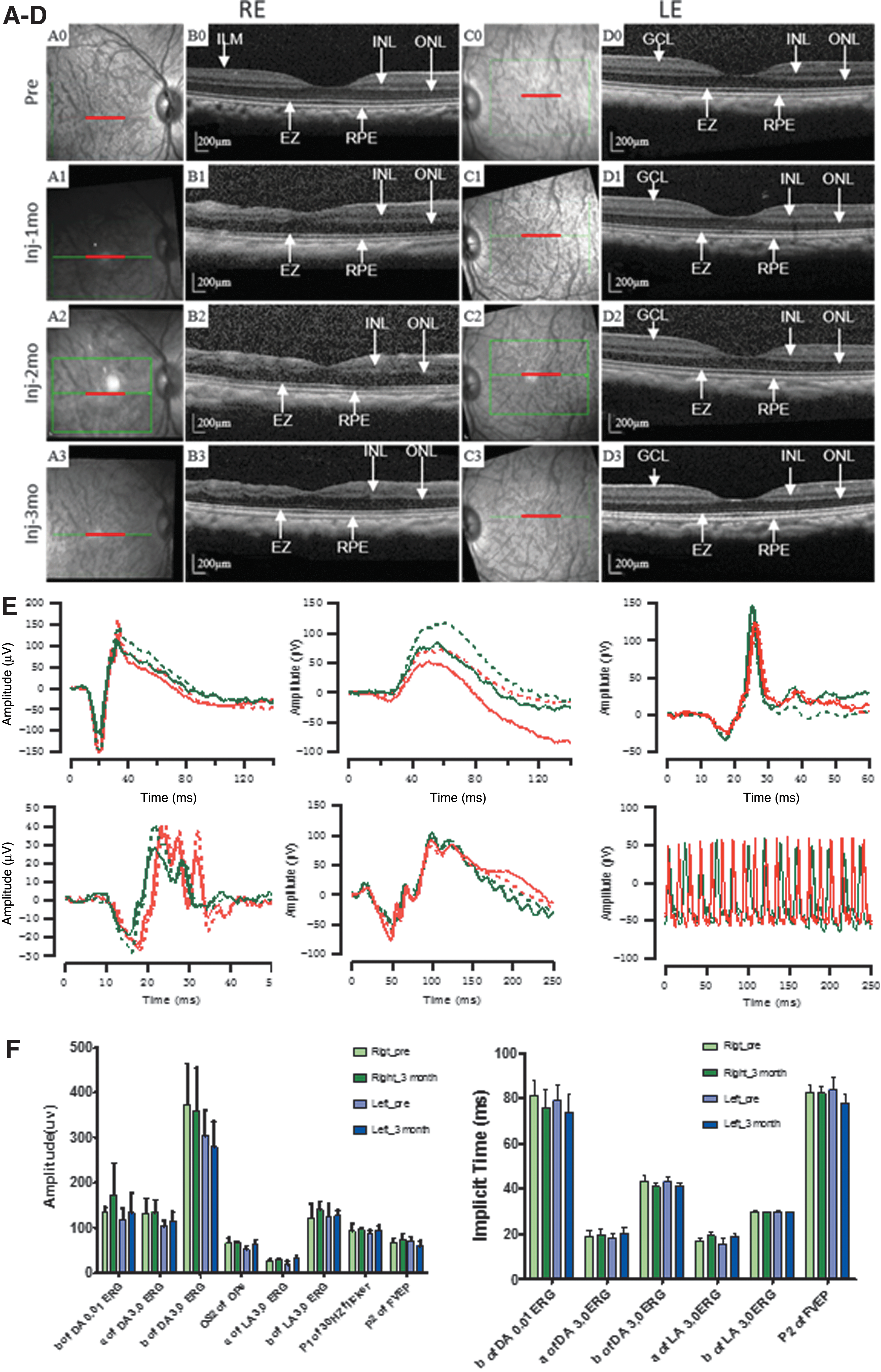
Optical coherence tomography (OCT) scanning and visual electrophysiology examination of macaques before and after the surgery.
Before and 3 months after surgery, fERG and FVEP tests were administered for parallel statistical analysis to observe whether the injection of the virus and the surgery caused serious eye complications that lead to functional ophthalmological changes. The waveforms of the fERG and FVEP are similar to the waveforms of humans (Fig. 3E). The right eyes' implicit timing and amplitude of the fERG and FVEP tests were not significantly different before and 3 months after posterior segment vitrectomy, ILM peeling, C3F8 gas filling, and intravitreal injection of AAV2/8-CMV-hMel/FYP vectors (Fig. 3E and F; p > 0.05, n = 3). There was no significant difference in the implicit time or amplitude of fERG and FVEP in the left eyes, which were intravitreally injected with the same viral vectors and tested before and 3 months after the injection (Fig. 3E and F; p > 0.05, n = 3). Moreover, the fundus fluorescein angiography and fundus photography data also suggest that the injection of the virus and surgery do not cause noticeable eye complications (Supplementary Fig. S3).
Safety of AAV2/8-CMV-hMel/FYP viral vector injection
Total T lymphocytes were determined with positive staining of anti-CD3. Analysis of T-lymphocyte subsets, namely CD4+ (T-helper cells) and CD8+ (cytotoxic cells), was performed. The CD4+/CD8+ ratio is a known indicator of the immune regulatory status. The results showed that the CD4+/CD8+ ratio after 1 month of AAV2/8-CMV-hMel/FYP injection was lower than the pre-injection ratio, as shown in Table 1, while CD8+ increased after 2 months. Alanine aminotransferase, aspartate transaminase, and interleukin 6 levels were progressively higher from 1 to 3 months. These data suggested that chronic inflammation may occur at 1 month due to surgery and AAV2/8-CMV-hMel/FYP administration. However, 3 months after intravitreal injection of the AAV2/8-CMV-hMel/FYP vector, no visible pathological changes were observed in the H&E staining of paraffin sections of the heart, liver, spleen, kidney, brain, optic nerve, optic tract, or visual cortex (Fig. 4), while a blood examination showed similar results (Supplementary Fig. S4).
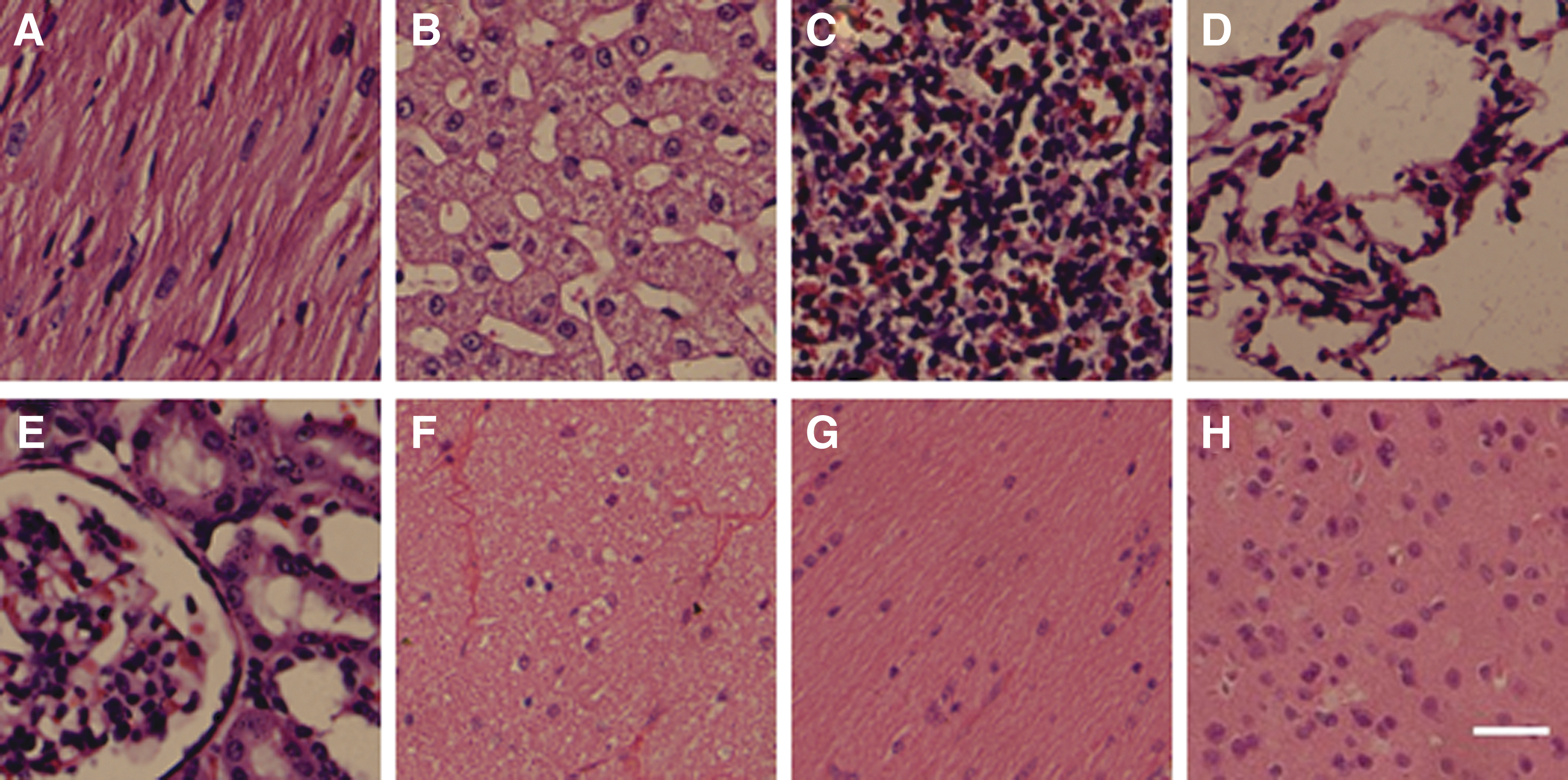
Pathological examination of the main organs in macaques after administration of the AAV2/8-CMV-hMel/FYP vector. Hematoxylin and eosin staining of paraffin sections in macaque no. 3 at 3 months after intravitreal injection of AAV2/8-CMV-hMel/FYP in the
Blood examination and T lymphocytes of macaque no.3
ALT, alanine aminotransferase; AST, aspartate transaminase; ALP, alkaline phosphatase; TBIL, total bilirubin; hs-CRP, high-sensitivity C-reactive protein; IL-6, interleukin-6; PCT, procalcitonin.
Discussion
This study demonstrates that ectopic expression of melanopsin in the degenerated rodent retina is a strategy of partial restoration of visual function based on the following evidence. First, many RGCs were shown to express melanopsin after injection of AAV2/8-CMV-hMel/FYP. Next, the moderate amplitude of fERG and robust amplitude of the FVEP wave can be restored after transduction with the AAV2/8-CMV-hMel/FYP vector. Third, the group treated with the AAV2/8-CMV-hMel/FYP vector displayed better behavior performance than control vector animals in both the light-avoidance and visual acuity behavioral tests. To the authors' knowledge, vision deteriorates over 2 months in the RCS rat because of rod and cone degeneration followed by RPE functional defects. Although some photoreceptor cells exist in the retina up to P60d, flashlight responses were not detectable on fERG. 66 In this study, the b-wave and P1-wave of fERG and FVEP disappeared in the RCS control group after P70d, but light responses were recorded in the AAV2/8-CMV-hMel/FYP-injected RCS rats. Taken together, the results suggest that retinal neurons can transmit visual signals into electric signals and then transfer to the visual cortex by the ectopic expression of hMel.
What types of retinal degenerative diseases or what disease stages are suitable for this potential treatment strategy? In current and previous studies, two types of retinal degenerative animal models have been used: the rd1 mouse 50,67 and the RCS rat. 68 After hMel was delivered into the retina by intravitreal injection, FVEP was restored without fERG recovery, indicating that the method is a potential treatment for patients suffering photoreceptor loss diseases while preserving RGCs at the end stage of the disease, including RP or photoreceptor dystrophy.
Furthermore, a significant issue in progressing viral-mediated gene therapies for human retinal disease from small-animal efficacy studies to clinical application lies in the potential differences between the eye and retina of the animal model and those of humans. Studies in the ideal preclinical animal model, the macaque, are necessary to assess the safety and expression of the therapy. The retinal transduction of AAV2 eGFP vector by intravitreal injection into the macaque retina studied by Takahashi et al. 69 speculated that the limited transduction might be due to the physical barriers posed by the anatomy of the macaque retina, such as the ILM, which lies along the surface of the retina, between the ganglion cell layer and vitreous. This study divided six eyes of three normal macaques into two groups: three right eyes as a group, which were subjected to posterior segment vitrectomy, ILM peeling, C3F8 gas filling, and intravitreal injection of 30 μL of AAV2/8-CMV-hMel/FYP (virus titer 3.52 × 1013 vp/mL); and three left eyes as a group, which were subjected to direct intravitreal injection of 30 μL of the same AAV2/8-CMV-hMel/FYP vector without ILM peeling. These groups were designed to observe whether the ILM peeling would influence the transduction of the viral vector and whether the surgery is safe.
The posterior segment vitrectomy and ILM peeling led to some complications, such as intraocular hemorrhage, iatrogenic retinal tear, and retinal detachment. This study demonstrated that the AAV2/8-CMV-hMel/FYP vector mainly infects RGCs not INL cells regardless of ILM peeling, suggesting that although the viral vector has limited penetration capability from the intravitreal space to the retinal space, it is sufficient to penetrate the ILM and infect RGCs. This feature might be an advantage for specifically targeting RGCs without complicated surgery of ILM peeling. Second, once the cells were infected with the viral vector, the intense expression of hMel/YFP driven by the CMV promoter was achieved and lasted >2 months in rats and 3 months in macaques, suggesting that the CMV promoter is sufficient to drive target gene expression over a relatively long period.
Furthermore, to observe whether the exogenous virus would lead to genetic mutation, activate proto-oncogenes, inactivate anti-oncogenes, and result in tumors or other complications in the macaque tissue, pathological observations were performed on the macaque heart, liver, kidneys, and other body organs and tissues. No visible pathological changes were observed on the H&E staining of paraffin sections of the systematic organs.
In summary, this study demonstrates that ectopic expression of hMel in the degenerated retinae of RCS rats restores visual function, and intravitreal injection of the AAV2/8-CMV-hMel/FYP vector without ILM peeling can achieve sufficient preclinical results in nonhuman primates.
Footnotes
Acknowledgments
We thank Dr. Bin Lin for providing the AAV hMel vector and Dr. ZhuoHua Pan for proofreading and commenting on the manuscript. This work was supported by the Military Key Program BWS13C015 and Chongqing International Cooperation Key Projects (No. CSTC2013GJHZ10004).
Author Disclosure
The authors have no conflicts of interest to declare.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.