
Abstract
It has previously been demonstrated that the self-inactivating γ-globin lentiviral vector GGHI can significantly increase fetal hemoglobin (HbF) in erythroid cells from thalassemia patients and thus improve the disease phenotype in vitro. In the present study, the GGHI vector was improved further by incorporating novel enhancer elements and also pseudotyping it with the baboon endogenous virus envelope glycoprotein BaEVRless, which efficiently and specifically targets human CD34+ cells. We evaluated the hypothesis that the newly constructed vector designated as GGHI-mB-3D would increase hCD34+ cell tropism and thus transduction efficiency at low multiplicity of infection, leading to increased transgene expression. High and stable HbF expression was demonstrated in thalassemic cells for the resulting GGHI-mB-3D/BaEVRless vector, exhibiting increased transduction efficiency compared to the original GGHI-mB-3D/VSVG vector, with a concomitant 91% mean HbF increase at a mean vector copy number per cell of 0.86 and a mean transduction efficiency of 56.4%. Transduced populations also exhibited a trend toward late erythroid, orthochromatic differentiation and reduced apoptosis, a further indication of successful gene therapy treatment. Monitoring expression of ATG5, a key link between autophagy and apoptosis, it was established that this correction correlates with a reduction of enhanced autophagy activation, a typical feature of thalassemic polychromatophilic normoblasts. This work provides novel mechanistic insights into gene therapy–mediated correction of erythropoiesis and demonstrates the beneficial role of BaEVRless envelope glycoprotein compared to VSVG pseudotyping and of the novel GGHI-mB-3D/BaEVRless lentiviral vector for enhanced thalassemia gene therapy.
Introduction
The β-thalassemia syndromes represent a group of monogenic, autosomal recessive blood disorders characterized by reduction or absence of the β subunit of the α2β2 heterotetramer of adult hemoglobin (HbA). More than 380 different mutations affecting the human β-globin gene and linked to β-thalassemia have been described, which are responsible for the significant heterogeneity of the thalassemic phenotype. 1 A key determinant of the disease pathophysiology is the excess of α-globin chain molecules, which precipitate in erythroid precursors, leading to impaired erythrocyte maturation, mechanical damage, and ultimately to apoptosis. 2
The only curative therapy for β-thalassemia so far is the allogeneic hematopoietic stem cell (HSC) transplantation of HLA-matched donors, 3 which is limited by the availability of compatible donors and the need for long-term immunosuppression to prevent or avert graft-versus-host disease. Routine disease management for β-thalassemia is instead based on chronic blood transfusions combined with lifelong iron chelation at a significant cost. Additionally, hydroxyurea may be used to supplement palliative treatment and reduce transfusion requirements by induction of fetal γ-globin. The latter combines with α-globin to form fetal hemoglobin (HbF; α2γ2) and thus alleviate anemia and reduce toxicity caused by the excess of α-globin. In light of the substantial drawbacks of the currently available treatment modalities, a molecular approach based on gene therapy has therefore long been pursued in an attempt to exploit the benefit of using autologous transplantation of genetically corrected cells. 4
A large number of studies have demonstrated that lentiviral vectors (LVs) with either β-globin 5 –11 or γ-globin 12 –14 gene addition can ameliorate thalassemia phenotype in vitro, while the first successful β-thalassemia gene therapy clinical trial 7 has been followed by several additional clinical trials based on similar VSVG-pseudotyped LVs, incorporating regulatory elements of β-globin locus control region (LCR). 10,15 –17 Data from preclinical and clinical studies using β-globin 16,18 –20 and γ-globin 19 LVs for sickle cell disease (SCD) treatment are also available, suggesting that gene therapy of SCD with globin LVs is also possible.
Previous work from the authors' group demonstrated that the LCR-free, self-inactivating γ-globin LV, designated as GGHI, led to significant HbF increase and corrected erythropoiesis in primary erythroid cells from thalassemic patients. 14 In the present study we incorporated the HPFH-1 enhancer element and the 3′ β-globin untranslated region (UTR) in the above vector, constructing the novel GGHI-mB-3D vector. We further pseudotyped GGHI-mB-3D with either the conventional VSVG or the alternative envelope glycoprotein BaEVRless 21,22 in order to exploit its capacity for tropism toward human HSCs, 23 and assessed the novel vectors in vitro at a low multiplicity of infection (MOI) using CD34+ cells from both normal and thalassemic individuals. Results showed that GGHI-mB-3D/BaEVRless LV achieved stable and high HbF expression in vitro, without marked positional effects and silencing. Furthermore, GGHI-mB-3D/BaEVRless demonstrated increased transduction efficiency upon a single vector incubation at a low MOI and improvement of erythropoiesis and reduction of apoptosis at a clinically relevant vector copy number (VCN)/cell of 0.86.
Methods
GGHI-mB-3D vector construction
In order to construct the GGHI-mB-3D vector, the 1,050 bp BamHI-EcoRI fragment of the pBR322 plasmid, containing the 3D enhancer element of the HPFH-1 deletion, was initially inserted into the XhoI site of the previously characterized GGHI vector 14 by blunt-end ligation, creating the GGHI-3D vector (Supplementary Fig. S1A). The BamHI-EcoRI 3D fragment was inserted in the same orientation with the transcription of the Aγ-globin gene. Then, the BstBI-BSu36I fragment of the GGHI vector, 14 containing the γ-globin 3′ UTR, was replaced with the 750 bp fragment of the β-globin 3′ UTR, present in the previously characterized γ-globin vector V5m3-400, 24 generating the GGHI-mB vector (Supplementary Fig. S1A). The BstBI site of the GGHI vector was created by introducing a point mutation with a QuickChange II Site-Directed Mutagenesis Kit (Agilent, Santa Clara, CA). Finally, the GGHI-mB-3D vector (Fig. 1 and Supplementary Fig. S1), which combines the 3D enhancer element and the β-globin 3′ UTR, was constructed by sub-cloning of the GGHI-mB 1,234-bp BSU36I-NsiI fragment into the GGHI-3D vector.
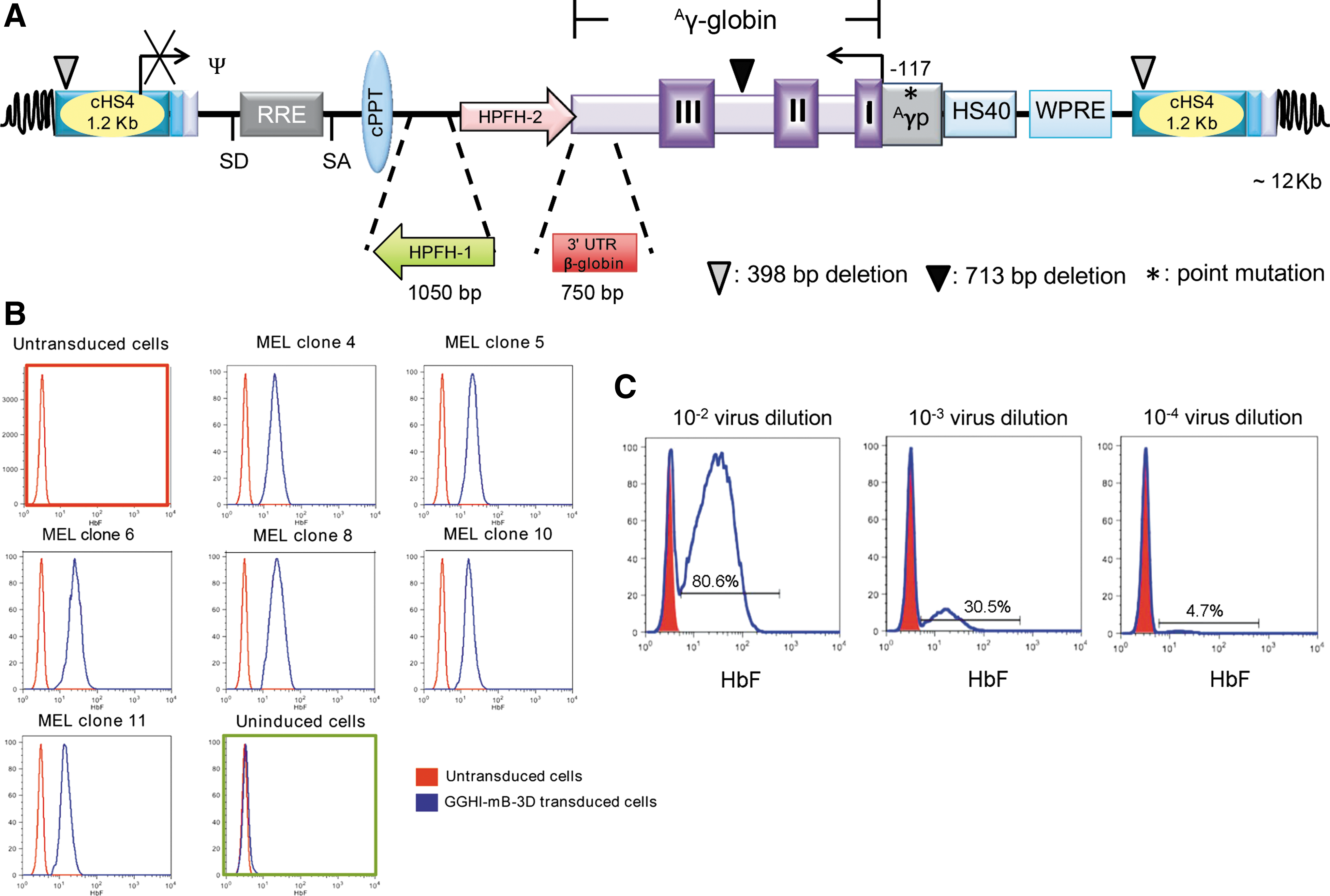
Schematic representation and initial characterization of the novel GGHI-mB-3D lentiviral vector (LV) in MEL-585 cells.
Virus production and titration
Viruses were produced by transient co-transfection of HEK-293T cells using a third-generation lentiviral system 25,26 comprised of the following plasmids: GGHI-mB-3D for γ-globin vector and GFPI for reporter vector, 27 as well as plasmids pMDLg/pRRE and pRSV-Rev, 25 a kind gift from Dr. Luigi Naldini, San Raffaele Telethon Institute for Gene Therapy, Milano, Italy. Vector production was carried out as previously described. 14,22,28 Briefly, 107 cells were seeded in 175 cm 2 flasks and transfection was performed at 80% confluency using calcium phosphate precipitation with 21 μg of VSVG/BaEVRless, 35 μg of pMDLg/pRRE, 18.2 μg of pRSV-Rev, and 70 μg of either γ-globin or GFPI plasmids. Virus-containing supernatant was collected as described by Papanikolaou et al. 27 Briefly, virus-containing supernatant was collected 48 and 72 h post transfection and concentrated 100-fold by ultrafiltration at 4,000 g for 40 min using 100 kDa centrifugal units (Millipore, Darmstadt, Germany). The concentrated supernatant was aliquoted and stored at −80°C.
Titration of the VSVG-pseudotyped viruses was determined by transducing 5 × 105 mouse erythroleukemia (MEL-585) cells with serial dilutions of concentrated virus as described by Papanikolaou et al. 14 More specifically, MEL-585 cells were allowed to differentiate for 3–4 days with 10 μM of hemin (Sigma–Aldrich, St. Louis, MO) and 3 mM of HMBA (N,N′-hexamethylene bisacetamide; Sigma–Aldrich), prior to detection of transgene expression by flow cytometry. Titration of VSVG-pseudotyped LVs in HEK-293T cells resulted in similar biological titers to the ones obtained with MEL-585 cells (representative LOT1 titration shown in Supplementary Table S3), with titers determined by either flow cytometry or quantitative polymerase chain reaction (qPCR) showing little or no differences (Supplementary Table S3) and in line with Girard-Gagnepain et al. 22 BaEVRless-pseudotyped viruses were titrated using HEK-293T cells, as described elsewhere, 29 using 1:50, 1:100, 1:200, and 1:1,000 dilutions of concentrated GFPI/BaEVRless plasmid stocks to transduce 50,000 cells. GFPI expression was assessed by flow cytometry on day 5. Dilutions yielding 1–20% GFPI-positive cells were used for titer calculation. In parallel, GFPI-transduced cells in the above dilutions were subjected to DNA isolation using the NucleoSpin Tissue Kit (Macherey-Nagel, Düren, Germany), followed by absolute quantitation using qPCR (SaCycler; Sacace Biotechnologies, Como, Italy) based on SYBR Green mix (Kapa Biosystems, Wilmington, MA) and the following pairs of primers: WPRE F: 5′-TTCTCCTCCTTGTATAAATCCTGGTT-3′ and WPRE R: 5′-CGCCACGTTGCCCA-3′ and hRNaseP F: 5′-ATGGAATTCGAACGCTGACG-3′ and hRNaseP R: 5′-GCGACATGCAAATATTGCAGG-3′. Standards of cells containing 0.1–0.005 copies of lentivirus per cell were used to produce the following standard curve: y = −4.5012x + 4.1418 (R 2 = 1), with y being DCt (Ct WPRE-CThRNAseP) and x the VCN, which was then used to extrapolate the unknown titer (TU/m) in GGHI-mB-3D/BaEVRless-transduced cells.
Human CD34+ cell isolation and transduction, sample collection, and processing
CD34 + hematopoietic stem cells isolated from three normal and nine β-thalassemia patients were used. Diagnosis of β-thalassemia was based on family history, complete transfusion-dependence history, and hemoglobin electrophoresis. All samples were obtained from non-mobilized peripheral blood and were harvested from volunteer donors using protocols approved by the local Institutional Review Board at the Saint Sophia Children's Hospital of Athens, based on written informed consent in accordance with the Declaration of Helsinki. Thalassemic samples were obtained shortly before the next scheduled blood transfusion. CD34 + cells were isolated from mononuclear cells using the EasySep Human CD34+ Selection Kit and the Purple EasySep magnet (STEMCELL Technologies, Cambridge, United Kingdom), according to the manufacturer's instructions. Samples obtained were >90% enriched in CD34 + cells (data not shown) and were cultured for up to 18 days in erythroid liquid cultures, as described previously 14 and shown in Supplementary Fig. S2. Briefly, CD34+ cells were re-suspended overnight in pre-transduction serum-free medium (StemSpan serum-free expansion medium [SFEM]; STEMCELL Technologies) at a density of 104–105 cells/mL containing the following recombinant human cytokines, all purchased from ImmunoTools (Friesoythe, Germany): 100 ng/mL of interleukin 6, 100 ng/mL of Flt-3 ligand, 50 ng/mL of thrombopoietin, and 100 ng/mL of stem cell factor. The following day, 104–105 cells were transduced in the same medium and in 48-well Retronectin-coated plates (Takara, Mountain View, CA) with either GFPI or γ-globin vectors at various MOIs. Mock-transduced cells were kept as controls. Cells were then washed, and 103 cells/mL/35 mm2 Petri dish were plated in MethoCult® H4434 methylcellulose medium (STEMCELL Technologies) for colony-forming assays. Colonies were picked 2 weeks later and used for DNA isolation and determination of VCN and the vector's transduction efficiency. The remaining cells were placed in liquid erythroid cultures for up to 18 days in StemSpan SFEM, containing 20% fetal bovine serum (FBS), 50 ng/mL of human stem cell factor (ImmunoTools), 10−6 M of β-estradiol (Sigma–Aldrich), 1 ng/mL of interleukin 3 (ImmunoTools), 1 IU/mL of erythropoietin (ImmunoTools), and 10−6 M of dexamethasone (Sigma–Aldrich). Analyses of liquid cultures included high-performance liquid chromatography (HPLC) and flow cytometry for the assessment of HbF production, microscopic scoring for erythroid cell expansion and differentiation, and Wright–Giemsa staining of cytocentrifugation samples for the assessment of orthochromatic erythroblasts. Apoptosis assessment was carried out with flow cytometry using the Annexin-V/7AAD detection kit (BioLegend, San Diego, CA) on cells harvested at day 18. RNA isolation was carried out on day 15 erythroblasts, as described below.
Reverse-phase HPLC analysis for HbF quantitation
In line with published methods, 30,31 cells were lysed by re-suspension in H2O (target concentration: 106 cells/25 μL) and two freeze–thaw cycles. After sedimentation at maximum velocity, the supernatant was transferred to 100 μL HPLC microvials (Altmann Analytik, Munich, Germany). Aliquots of 25 μL of cell lysate (or up to 50 μL for samples with low cell numbers or visibly low levels of differentiation) were injected per analysis. Samples were separated on a Prominence HPLC machine with an LC-20AD binary pump and an SPD-M20A photodiode array detector (Shimadzu, Kyoto, Japan). As stationary phase, a 25 cm C18 Jupiter column with corresponding C18 SecurityGuard cartridge was used (Phenomenex, Torrance, CA). As mobile phases, Phase A, which was 0.1% trifluoroacetic acid (TFA; Sigma–Aldrich) and 0.033% sodium hydroxide (Fluka, Munich, Germany), was used in a linear gradient against Phase B, which was 2:1 (v:v) acetonitrile:methanol (from 56% baseline to 68%) or pure acetonitrile (from 40% baseline to 46%) HPLC grade (Sigma–Aldrich).
For liquid cultures, about 5 × 106 cells were collected, while for semi-solid cultures, 10–15 pooled erythroid burst-forming units (BFUe)-derived colonies per patient were subjected to HPLC.
The Aγ/α globin chain ratios were calculated for each patient sample, which was mock-transduced or transduced with either GGHI-mB-3D/VSVG or GGHI-mB-3D/BaEVRless LVs. The percentage of HbF mean increase achieved in each case was corrected by dividing with the corresponding mean VCN.
RNA analysis using qPCR
Transcript levels were quantified by reverse-transcription qPCR based on SYBR Green chemistry and performed on a SaCycler (Sacace Biotechnologies). All primer sets were picked using Primer Express v3.0 (Applied Biosystems, Foster City, CA).
For quantification of γ-globin production and ATG5 from erythroid cultures, total RNA was isolated from both mock-transduced and transduced CD34 + cells on day 15 using the NucleoSpin RNA Plus Kit (Macherey-Nagel) and according to the manufacturer's instructions. Total RNA (100–300 ng) was reverse transcribed to cDNA using Superscript II (Invitrogen, Carlsbad, CA), and 20 ng of cDNA was then subjected to qPCR analysis. Production of γ-globin was measured using the following primers: gamma F, 5′-GCCATAAAGCACCTGGATGA-3′, and gamma R, 5′-GATTGCCAAAACGGTCACC-3′. The human α-globin gene was used as a reference gene for the level of differentiation based on the following primers: alpha-globin F, 5′-CACGCTGGCGAGTATGGTG-3′, and alpha-globin R, 5′-TTAACCCTGGGCAGAGCCGT-3′. Autophagy marker ATG5 mRNA was quantified using primers: ATG5 F, 5′-GCAGATGGACAGTTGCACACAC-3′, and ATG5 R, 5′-GAGGTGTTTCCAACATTGGCTCA-3′. The β-actin gene was employed as a reference gene for cDNA quantity calibration using the following primers: β-actin F, 5′-CCACACTGTGCCCATCTACG-3′, and β-actin R, 5′-CAGCGGAACCGCTCATTGCCAATGG-3′. Measurements of γ-globin, α-globin, ATG5, and β-actin mRNA levels were performed in duplicate for each sample using SYBR Green mix (Kapa Biosystems) under the following conditions: enzyme activation at 95°C for 3 min, denaturation at 95°C for 3 s, and annealing/extension at 60°C for 20 s for 40 cycles. In order to determine the fold increase of mRNA in transduced versus mock-transduced cell populations, the ΔΔCT method was used. 32
Determination of vector copy number and transduction efficiency
In order to assess vector transduction efficiency, 10–20 BFUe colonies per patient were subjected to colony PCR analysis using the γ-globin primers set described above. These primers generate a vector-specific product of 282 bp and a 1,009 bp product that corresponds to the endogenous human γ-globin gene. Each sample was analyzed in a 25 μL reaction using GoTaq (Promega, Madison, WI) and according to the manufacturer's instructions. Amplification was carried as follows: initial denaturation at 95°C for 10 min, 40 cycles of 95°C for 15 s, 60°C for 30 s, 72°C for 1 min, and final extension at 72°C for 15 min. Vector gene transfer efficiency in CD34+ cells was determined by assessing the proportion of BFUe colonies that were tested positive for the vector sequence. Those testing positive for vector-specific sequences were further subjected to absolute quantitation with qPCR analysis for VCN determination using the γ-globin and hRNase P primer sets, as previously described. 14 Analysis of each sample was performed in duplicate using SYBR Green mix (Kapa Biosystems) under the following conditions: enzyme activation at 95°C for 3 min, denaturation at 95°C for 3 s, and annealing/extension at 60°C for 20 s for 40 cycles.
Flow cytometry and apoptosis assay
VSVG-pseudotyped γ-globin LVs were titrated with flow cytometry as previously described 14 using a FITC-conjugated anti-HbF monoclonal antibody (BD Pharmingen, Franklin Lakes, NJ). With regards to GFPI vectors, GFP expression was measured on live cells gated according to their FSC/SSC profile.
Mock-transduced and transduced CD34+ cells were processed for flow cytometry analysis on days 15–18 using an anti-HbF FITC-conjugated monoclonal antibody (BD Pharmingen) and a Cytofix/Cytoperm kit (BD Pharmingen). In order to test the efficiency of erythropoiesis in erythroid cultures, cells were also stained with anti-glycophorin A (CD235a) PE-conjugated monoclonal antibody (BD Pharmingen). It should be noted that flow cytometry detection of HbF is not Aγ-globin specific, in contrast to HPLC-based detection, and therefore underestimates the percentage of Aγ-globin increase achieved by γ-globin vectors.
Apoptotic assays were performed on cells derived from erythroid cultures on day 18 using a FITC Annexin-V Apoptosis Detection kit with 7-AAD (Biolegend), according to the manufacturer's instructions. Briefly, cells were washed twice with cold phosphate-buffered saline (PBS) containing 1% FBS and then re-suspended in Annexin-V binding buffer at a concentration of 0.25–1.0 × 107 cells/mL. Cells were then incubated for 15 min at room temperature in the dark and analyzed by flow cytometry within 1 h.
All samples were analyzed in a Cytomics FC 500 CXP Series Flow Cytometry System (Beckman Coulter, Nyon, Switzerland). Flow cytometry analysis was performed using FlowJo v8.7 analysis software.
Statistical analysis
Student's t-test was used to determine statistically significant differences between mean values of different data sets, and Pearson's r was used for correlation between values, both using Graph Pad Prism v7 (GraphPad Software, Inc., La Jolla, CA). The latter software was also employed for outlier identification.
Results
Novel γ-globin LVs GGHI-mB-3D/VSVG and GGHI-mB-3D/BaEVRless are erythroid specific and allow high titer production
The construction of the novel γ-globin LV GGHI-mB-3D (Fig. 1A) was based on the previously characterized LV GGHI 14 and the incorporation of novel regulatory elements, which demonstrated an additive effect on γ-globin expression (Supplementary Fig. S1). Initially, we asked whether the incorporation of a second enhancer element, that is, the 3D fragment of HPFH-1 33 in the previously characterized GGHI vector 14 carrying HPFH-2 enhancer element, has an additive effect on HbF expression. To this end, we cloned the 3D enhancer element from the natural occurring HPFH-1 deletion of the β-globin locus at the 3′ end of the Aγ-globin expression cassette, next to the HPFH-2 enhancer and in the same orientation as the transcription of the Aγ-globin gene, thus creating GGHI-3D. Following MEL-585 cells' transduction at a MOI of 20 (see Supplementary Data), HbF expression by GGHI and GGHI-3D LVs in positive MEL-585 clones (n = 6) was monitored for up to 4 months. GGHI-3D led to stable and significantly higher HbF levels compared to GGHI (p = 0.0045, n = 6; Supplementary Fig. S1B), demonstrating a clear synergistic effect of HPFH-1 and HPFH-2 enhancer elements on γ-globin expression. Based on the above, the 3′ β-globin UTR, shown to offer stability at the mRNA level, 24,34 –37 was ultimately combined with the aforementioned vector, creating the GGHI-mB-3D vector (Supplementary Fig. S1A). Comparison of HbF expression driven by all LVs in MEL clones at the end of the 4-month period showed that GGHI-mB-3D can also lead to significantly higher HbF compared to GGHI, both at the protein level (p = 0.033, n = 8; Supplementary Fig. S1C) and mRNA level (p = 0.0029, n = 6; Supplementary Fig. S1D), while both GGHI-3D (p = 0.0084, n = 7) and GGHI-mB-3D (p = 0.022, n = 8) LVs containing both HPFH-1 and HPFH-2 enhancer elements are superior to GGHI-mB regarding HbF expression. Cloning of 3′ β-globin UTR alone into GGHI, thus creating GGHI-mB, is not sufficient to raise HbF levels significantly (p = 0.88, n = 5), as shown in Supplementary Fig. S1C, despite the significantly increased human γ-globin mRNA transcript observed (p = 0.0052, n = 3; Supplementary Fig. S1D). It is conceivable that post-transcriptional modifications and affinity of human γ-globin for mouse α-globin chains may account, among others, for the above discrepancies in protein and mRNA expression. No statistical significance was observed between GGHI-3D and GGHI-mB-3D at the protein level (p = 0.669, n = 8), while human γ-globin mRNA expression was raised more than eightfold upon addition of the 3′ β-globin UTR into GGHI-3D (p = 0.0044, n = 6; Supplementary Fig. S1D). Moreover, compared to the GGHI base vector, GGHI-mB-3D achieved a 15-fold (p = 0.0029, n = 6) increase in γ-globin mRNA expression at a significantly lower VCN/cell (2 vs. 5.4, p = 0.02, n = 6; Supplementary Table S1). The 1.5-fold higher mRNA expression compared to GGHI-mB was accompanied by substantial intragroup variation for both LVs, so that both vectors did not test significantly different. The novel γ-globin GGHI-mB-3D LV (Fig. 1A) was functional and stable and expressed γ-globin in a pancellular and erythroid-specific manner throughout the 4-month inspection period. Titration of GGHI-mB-3D/VSVG virus led to titers as high as 1.6 × 108 TU/mL, as calculated by serial dilutions of virus supernatants 14,27 and shown in Fig. 1C and Supplementary Table S2.
Based on these features of the novel vector, GGHI-mB-3D vector was then pseudotyped with the alternative envelope glycoprotein BaEVRless, shown to exhibit tropism toward human HSCs. 21,22 In the case of BaEVRess-pseudotyped GFPI vectors, 27 two batches (LOTs 5 and 7) led to titers <107 TU/mL (Supplementary Table S2), as a result of syncytia formation and packaging cells' toxicity induced by the deletion of the fusion-inhibitory R sequence, 38 while the remaining five expressed high biological titers (Supplementary Tables S2 and S3). Regarding the therapeutic γ-globin vectors (Supplementary Tables S2 and S3), high mean titers (TU/mL) were observed for both BaEVRless-pseudotyped (1.9 × 108 ± 5.6 × 107 TU/mL) and VSVG-pseudotyped (1.6 × 108 ± 3.2 × 107 TU/mL) LVs. Overall, in the specific batches produced, no significant pseudotype-related differences in vector titers were observed for both GFPI (p = 0.13, n = 7) and γ-globin encoding LVs (p = 0.58, n = 5).
Stable and high transgene expression at a low MOI with novel γ-globin LV GGHI-mB-3D/BaEVRless in healthy individuals
Prior to the in vitro assessment of GGHI-mB-3D/VSVG and GGHI-mB-3D/BaEVRless vectors in thalassemic CD34+ cells, the study aimed to assess whether these novel vectors can demonstrate high and stable transgene expression in liquid cultures throughout erythroid differentiation. To this end, CD34+ cells were isolated from the peripheral blood of healthy donors and were initially transduced with GFPI/VSVG or GFPI/BaEVRless vectors at MOI of 10 and 25. Transduction was carried out for 24 h in the presence of Retronectin and cytokines, as previously described 14 (Supplementary Fig. S2) and outlined in the Methods.
Figure 2A shows the percentage of GFPI-positive cells at different time points of erythroid differentiation in liquid cultures after a single transduction with either GFPI vector pseudotype at low MOIs. The BaEVRless-pseudotyped vector led to significantly higher GFPI expression levels at both MOIs of 10 and 25 compared to its VSVG counterpart. Specifically, 8 days post transduction at a MOI of 10 (Fig. 2A, left panel), GFPI/BaEVRless, depicted in red, demonstrated a significantly increased and stable GFPI expression, leading to a mean peak percentage of GFPI-positive cells of 48% compared to 23.8% obtained with GFPI/VSVG vector, depicted in black (p = 0.0015, n = 3). Predictably, differences between BaEVRless and VSVG pseudotypes became more pronounced at a MOI of 25 (Fig. 2A, right panel) with BaEVRless vector, reaching a percentage of 65.1% GFPI-positive cells (p < 0.0001, n = 3) on day 8 compared to 24.7% observed in the case of the VSVG-pseudotyped vector.
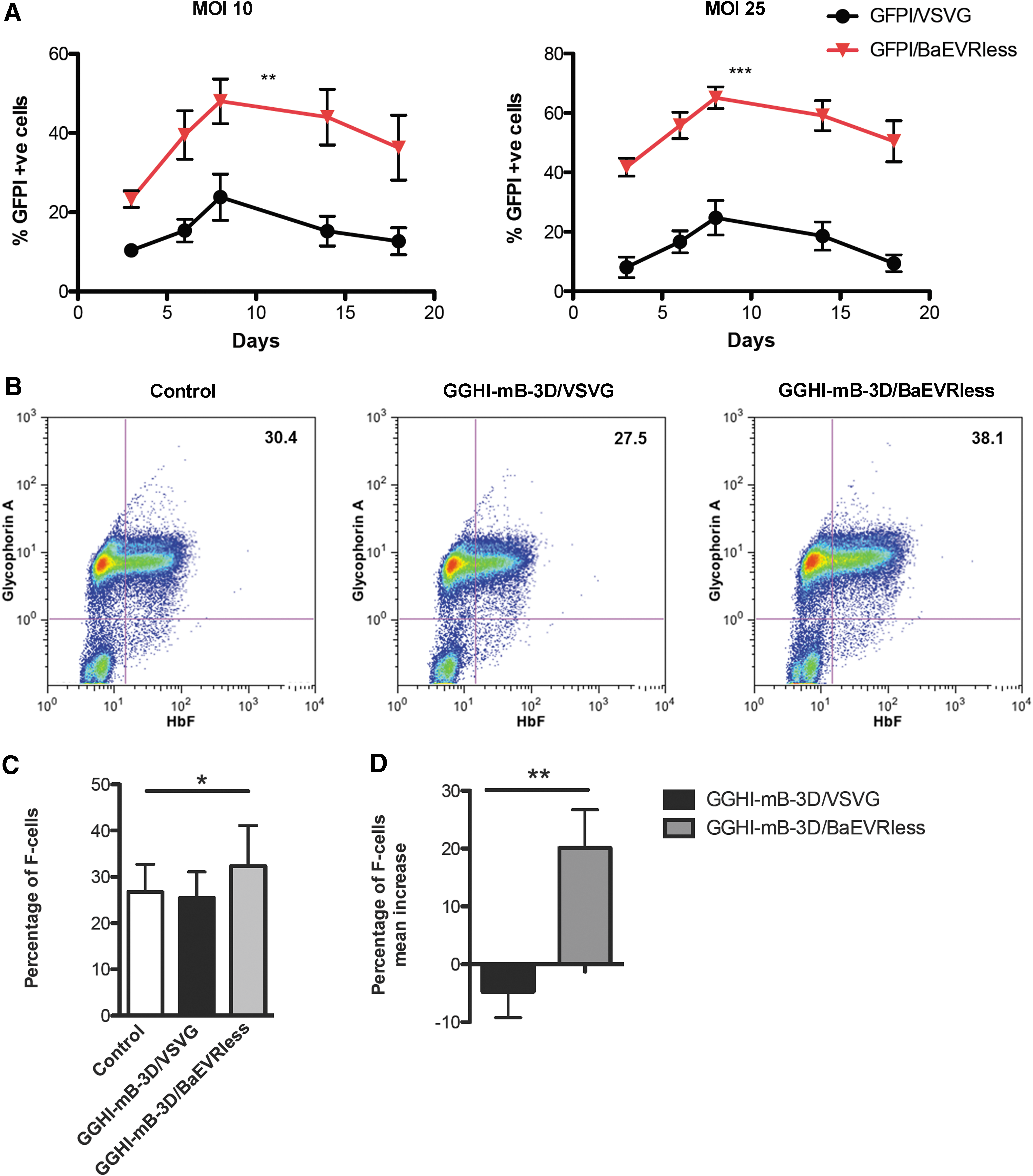
Assessment of novel LVs in normal CD34+ cells.
Given the increased transduction efficiency obtained with the BaEVRless-pseudotyped vector in the above culture conditions and the high transgene transfer observed at a MOI of 25, the study proceeded to the transduction of normal CD34+ cells with the γ-globin vectors using a MOI of 25, at which the highest transgene transfer was observed. In a representative experiment, GGHI-mB-3D/BaEVRless-transduced cells led to a 25% increase of cells positive for both anti-HbF and anti-glycophorin A monoclonal antibodies (referred as F-cells from this point onwards), reaching 38.1% compared to 30.4% observed in the control (Fig. 2B). Transduction with GGHI-mB-3D/VSVG at a MOI of 25 failed to increase the percentage of F-cells, which remained at baseline levels. Across three independent experiments (Fig. 2C), GGHI-mB-3D/BaEVRless led to a statistically higher percentage of F-cells, with a mean of 32.4 ± 8.7% compared to 26.7 ± 6% observed in the control (p = 0.0357, n = 3). The mean F-cell increase achieved with GGHI-mB-3D/BaEVRless vector was significantly higher than that observed with the VSVG counterpart, reaching a mean increase of 20.1 ± 6.6% compared to just −4.7 ± 4.5% (p = 0.0058, n = 3; Fig. 2D).
Increased HbF production in β-thalassemia patient CD34+ cells following transduction with the novel γ-globin LVs
Non-mobilized peripheral blood CD34+ cells from nine thalassemia major patients were isolated and processed. Five out of nine patients were homozygotes for β+-thalassemia mutations, and four patients were compound heterozygotes for β0 and β+ mutations. All nine patients had four intact α-globin genes, and all of them were transfusion-dependent since infancy. Typical yield from an initial volume of 20 mL of peripheral blood ranged from 2 × 104 to 3 × 105 CD34+ cells. Consistent with the vast majority of thalassemia major patients in Greece exhibiting high percentages of F-cells, 39 samples from the patient cohort also gave on average of 48.2 ± 14.7% F-cells in erythroid cultures, as detected by flow cytometry.
Based on the encouraging results from normal CD34+ cells, the efficacy of HbF induction by the novel γ-globin vectors was then assessed in thalassemic samples in vitro. To this end, thalassemic CD34+ cells were isolated and cultured under serum-free conditions for 18 h, dividing them into three subpopulations. Two subpopulations were transduced with each type of γ-globin vectors for 24 h in the presence of Retronectin, while the third was mock-transduced and served as a control. In order to achieve therapeutic HbF levels, that is, increase of HbF >30%, 40 the MOI used for vector comparisons in normal CD34+-derived cells was doubled to a MOI of 50. The following day, cells were washed twice with PBS and re-suspended in erythroid differentiation medium, as described in the Methods.
The progression of differentiation in these cultures was assessed on days 15–18 by flow cytometry using anti-HbF and anti-CD235a antibodies. Representative analyses and summarized results are depicted in Fig. 3, Tables 1 and 2, and Supplementary Fig. S3. As can be seen from the representative experiment shown in Fig. 3A, transduction with GGHI-mB-3D/BaEVRless vector led to an increased percentage of F-cells. Specifically, in the case of patient Thal 3, GGHI-mB-3D/BaEVRless-transduced F-cells reached 39.5% (mean fluorescence intensity [MFI] = 29.7), compared to 26.9% in the control (MFI = 10.9). F-cell increase following transduction with GGHI-mB-3D/BaEVRless vector was statistically significant (p = 0.02, n = 9), reaching a mean F-cell percentage of 54.6 ± 11.7% compared to 49.7 ± 12.8% of the control. Regarding transduction with GGHI-mB-3D/VSVG at a MOI of 50, the mean F-cell percentage of 50.8 ± 9.9% was close to control levels (p = 0.33, n = 9). The overall mean value of F-cell increase due to vector transduction, as observed by flow cytometry, was 5.5 ± 21.6% for GGHI-mB-3D/VSVG and 12.3 ± 17.4% for GGHI-mB-3D/BaEVRless (p = 0.47, n = 9; Fig. 3C). The study failed to detect a significant F-cell increase between GGHI-mB-3D/VSVG- and GGHI-mB-3D/BaEVRless-transduced cells due to the great variation among the thalassemic patient samples.
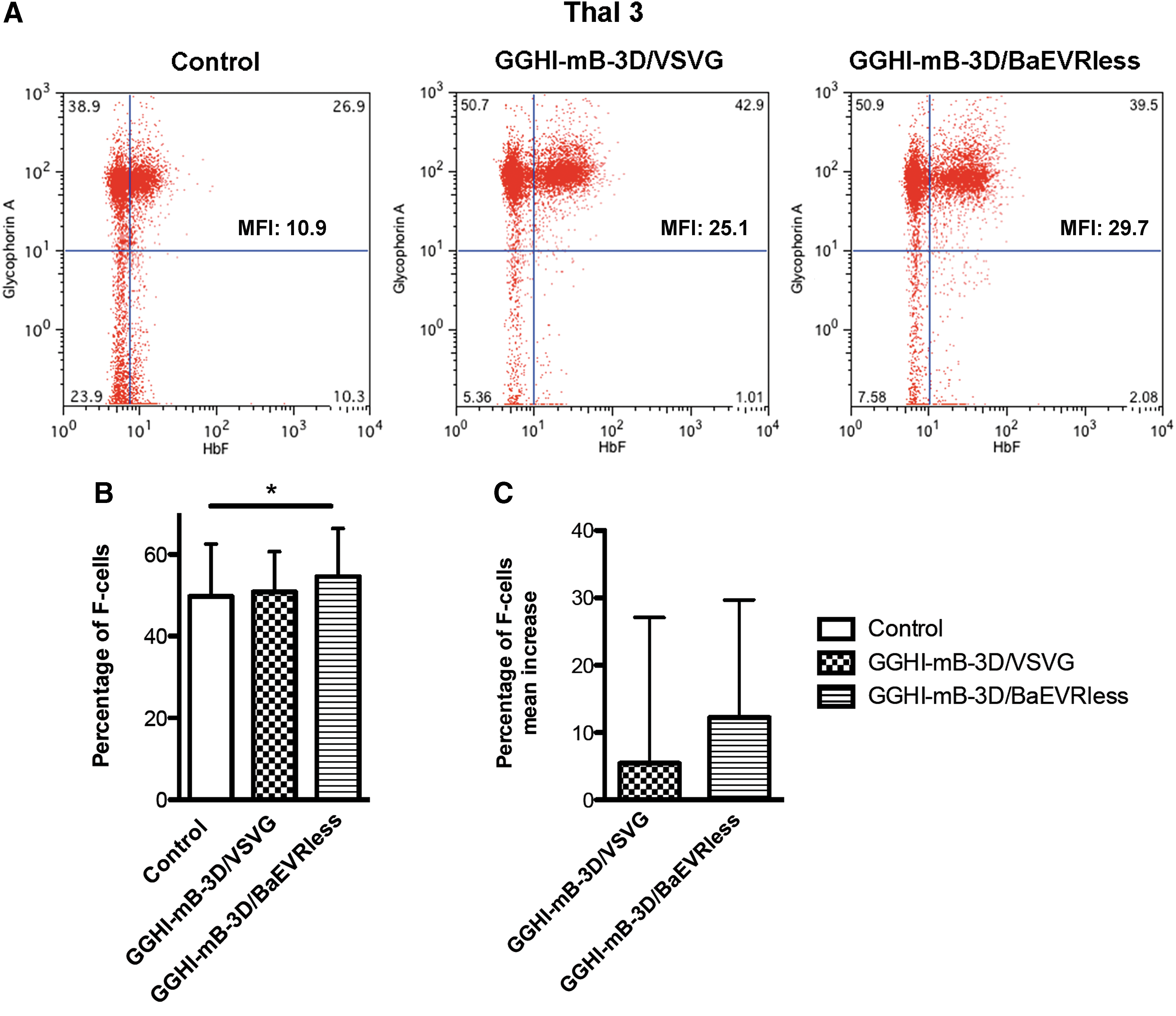
Functional correction of thalassemic CD34+ cells measured by flow cytometry.
Detailed data from all patients with GGHI-mB-3D/VSVG vector
Dashes indicate that the relevant assays were not done because of limited number of cells.
Outlier, not included in the statistical analysis.
FACS, fluorescence-activated cell sorting; HbF, fetal hemoglobin; HPLC, high-performance liquid chromatography; TSD, transduction; VCN, vector copy number; BFUe, erythroid burst-forming units.
Detailed data from all patients with GGHI-mB-3D/BaEVRless vector
Dashes indicate that the relevant assays were not done because of limited number of cells.
As a more sensitive and Aγ-globin-specific means of quantification, HPLC analysis was then employed for further characterization of the GGHI-mB-3D/VSVG and GGHI-mB-3D/BaEVRless vectors based on cell lysates from liquid cultures from all experiments. Under the specified experimental conditions, both mock and transduced cells generated measurable amounts of HbF (Tables 1 and 2 and Supplementary Fig. S3). As can be seen from Fig. 4A and Table 2, transduction with GGHI-mB-3D/BaEVRless led to a statistically significant increase of Aγ/α ratio in liquid cultures, reaching an average of 0.74 ± 0.72 compared to 0.35 ± 0.23 observed in the control (p = 0.0308, n = 9). This corresponds to a mean increase of 118 ± 102% when normalized to mean VCN/cell. Transduction with the specific vector led to an increase >30% in 8/9 (88.9%) patients, reaching its highest levels in patients Thal 3 and Thal 8 (Fig. 4B) where the Aγ/α ratio was increased almost threefold. Specifically in the latter, it was raised from 0.087 to 0.252 (Fig. 4C). Regarding GGHI-mB-3D/VSVG vector (Fig. 4A and Table 1), transduction was not accompanied by a significantly increased Aγ/a ratio, which reached an average value of 0.37 ± 0.11 (p = 0.32, n = 8).

Functional correction of thalassemic CD34+ cells measured by high-performance liquid chromatography (HPLC).
Regarding HbF production at the clonal level, cell lysates from 15 to 20 pooled BFUe, from five patients for GGHI-mB-3D/BaEVRless, or from three patients for GGHI-mB-3D/VSVG were also subjected to HPLC (Tables 1 and 2). GGHI-mB-3D/VSVG and GGHI-mB-3D/BaEVRless LVs led to a mean HbF increase, normalized to mean VCN per patient, of 214% (n = 3) and 776% (n = 5), respectively. For GGHI-mB-3D/BaEVRless specifically, the highest increase of as much as 10-fold was achieved for patient Thal 9 (Table 2). No statistical significance was observed in the Aγ/α ratio between mock-transduced, GGHI-mB-3D/VSVG-, and GGHI-mB-3D/BaEVRless-transduced cells (Table 1 and 2).
Gene transfer efficiency and vector copy number in transduced thalassemic BFUe
To determine gene transfer efficiency, CD34+ cells derived from thalassemia patients that were either mock-transduced or transduced with GGHI-mB-3D/VSVG or GGHI-mB-3D/BaEVRless LVs were plated in methylcellulose semi-liquid medium for colony-forming assays; 15–20 individual BFUe per patient were analyzed in each experiment by colony scoring using PCR and vector-specific primers. Gene transfer efficiency of either LV in the CD34+ progenitors was determined by assessing the proportion of BFUe colonies that tested positive for the vector sequence. The mean transduction efficiency for GGHI-mB-3D/VSVG was 38.5 ± 20.3% (median 40%; range 9–73%), while for GGHI-mB-3D/BaEVRless it was 56.4 ± 13.6% (median 56%; range 36%–77%; Fig. 5A, Tables 1 and 2, and Supplementary Fig. S4). Overall, GGHI-mB-3D/BaEVRless exhibited a significantly higher transduction efficiency than GGHI-mB-3D/VSVG (p = 0.023, n = 9). The clonogenicity of either mock or γ-globin vector-transduced CD34+ cells was similar (data not shown). Furthermore, the VCN was calculated for each BFUe from each patient and then a mean value was extracted from all BFUe for each patient (Fig. 5B, Tables 1 and 2, and Supplementary Fig. S4). The mean VCN for GGHI-mB-3D/VSVG was 0.67 ± 0.50 (median 0.44; range 0.18–1.69), while for GGHI-mB-3D/BaEVRless it was 0.86 ± 0.48 (median 0.75; range 0.36–1.57).

Performance of LVs in thalassemic CD34+ cells, following transduction at a MOI of 50, measured at the DNA and RNA level.
Improvement of thalassemic erythropoiesis in vitro following transduction with GGHI-mB-3D/BaEVRless
To test the effects of transduction with the newly constructed LVs on the correction of the thalassemic erythropoiesis, CD34+ cells were placed in liquid cultures that promoted erythroid differentiation and thus recapitulated in vivo erythropoiesis (Supplementary Fig. S2). Effective differentiation at the proerythroblast stage in these cultures was documented on days 15–18 by employing flow cytometry and anti-glycophorin A monoclonal antibody as a marker of late-stage erythropoiesis. 41 The erythroid maturation was further quantified by performing differential counts of the percentage of orthochromatic erythroblasts on cytocentrifugation samples of liquid cultures on day 18. It is well documented that erythroid cells in thalassemic erythropoiesis are arrested at the polychromatophilic normoblast stage, with very little progression to the orthochromatophilic normoblast stage. 42 In the present case, GGHI-mB-3D-transduced CD34+ cells exhibited a trend toward higher levels of erythroid differentiation (Fig. 6B, right panel, and Fig. 6C), with the mean percentage of orthochromatophilic erythroblasts being 55 ± 20% (p = 0.40, n = 7) in GGHI-mB-3D/VSVG and 56 ± 14% (p = 0.26, n = 7) in GGHI-mB-3D/BaEVRless-transduced cells compared to 52.8 ± 17% in mock-transduced cells (Fig. 6B, right panel).

Effect of novel γ-globin LVs in thalassemic erythropoiesis.
In parallel, improvement of the thalassemic erythropoiesis in liquid cultures was assessed on day 18 with apoptosis assays employing flow cytometry and Annexin-V/7AAD monoclonal antibodies. The study shows that GGHI-mB-3D/BaEVRless vector can improve thalassemic erythropoiesis at a MOI of 50 by reducing apoptosis. In a representative experiment (Fig. 6A), transduction with GGHI-mB-3D/BaEV-Ress led to 25.7% apoptotic cells compared to 89.7% observed in mock-transduced cells. Overall, in nine thalassemic patients, transduction with GGHI-mB-3D/BaEVRless led to a mean percentage of apoptotic cells of 46.4 ± 18% (median 44%) compared to 58.6 ± 17% (median 54%) for the control (p = 0.064, n = 9; Fig. 6B, left panel), demonstrating a mean decrease of 17.6 ± 26.8% compared to control. Apoptosis decrease was observed in 5/9 (56%) patients, with the highest reduction reaching 71.3% observed in patient Thal 7 (Table 2). By comparison, GGHI-mB-3D/VSVG led to 52.2 ± 14% (median 49%) mean percentage of apoptotic cells (p = 0.109, n = 9; Fig. 6B, left panel), with a mean decrease of just 8.1 ± 17.9% compared to control. A decrease in the number of apoptotic cells was observed in 6/9 (67%) patients, with the highest again detected in patient Thal 7 (Table 1).
Finally, expression of autophagy initiation marker ATG5 43,44 in mock-transduced and GGHI-mB-3D/BaEVRless-transduced cells showed that autophagy initiation is decreased upon transduction with the latter vector. Overall, ATG5 relative expression decreased by 35% (n = 8) in GGHI-mB-3D/BaEVRless-transduced cells (Fig. 6D, left panel), while the decrease was more profound, reaching 60% (n = 5) in patients where apoptosis decrease was also documented, as shown in Fig. 6D (right panel).
Discussion
This study generated the novel γ-globin LV GGHI-mB-3D by sub-cloning the 3D enhancer element and the β-globin 3′ UTR into GGHI, demonstrating a significant synergistic effect of the novel elements in HbF expression. Then, GGHI-mB-3D was pseudotyped with the envelope glycoprotein BaEVRless, 21 –23 shown to exhibit tropism toward human HSCs and human B cells. 28 All newly constructed LVs were functional and drove stable and high transgene expression in a pancellular and erythroid-specific manner. Although vector titers tended to be lower for GFPI-encoding BaEVRless- than for VSVG-based vector (p = 0.13, n = 7), they were highly similar for the therapeutically relevant VSVG- and BaEVRless-based γ-globin-encoding vectors (Supplementary Table S2). In the case of GFPI/BaEVRless LVs, the deletion of the fusion-inhibitory R peptide, which induces syncytia formation and HEK-293T cell toxicity during virus production, 38 together with the method of titer determination (i.e., flow cytometry instead of qPCR) may partly account for the striking but nonsignificant difference between VSVG and BaEVRless LVs in the specific batches produced. Also, differences in the GFP/γ-globin lentiviral constructs may present a potential reason for titer discrepancy between GFPI and γ-globin vectors. In direct comparison with the VSVG-pseudotyped vector, initially normal and then thalassemic cells were transduced with a variety of genotypes to ascertain the ability of the novel vector to drive stable and high HbF expression and eventually correct the thalassemic phenotype in vitro. The study shows that GGHI-mB-3D/BaEVRless consistently outperforms the GGHI-mB-3D/VSVG vector at low MOI, both in normal and in thalassemic cells, for HbF expression, correction of thalassemic phenotype, and apoptosis. These results were achieved at an average VCN of 0.86, calculated per haploid genome, approaching the proposed ideal target VCN range of one to two copies per cell for LV-based thalassemia gene therapy. 45 Specifically, the patient cohort was comprised of nine β-thalassemia patients, with an average HbF in erythroid culture of 48.2% prior to transduction. As previously demonstrated by Papanikolaou et al., 14 the culture system employed in this study leads to minimal culture-related induction of HbF in vitro, permitting an accurate assessment of the vector-derived net γ-globin production. The results show that with the exception of Thal 5, all patients exhibited a HbF increase (corrected to mean VCN/cell) >30% 40 following transduction with GGHImB3D/BaEVRless. In the case of GGHI-mB-3D/VSVG, 5/8 (63%) patients (Thal 2, Thal 4, Thal 8, Thal 9, and Thal 10; Thal 12 was excluded as an outlier) exhibited an increase >30%, outperforming the BaEVRless counterpart on four occasions (Thal 2, Thal 4, Thal 8, and Thal 10). This is possibly due to a higher proliferation rate in these patients post transduction. Representative expansion profiles in erythroid cultures, expressed as total cell counts, are shown in Supplementary Fig. S5. Overall, transduction with GGHI-mB-3D/BaEVRless led to a marked HbF increase, reaching 118 ± 102% (n = 9) on average based on HPLC analysis and following correction to VCN. Specifically, GGHI-mB-3D/BaEVRless vector raised the Aγ/α ratio to an average of 0.74 compared to 0.35 observed in the control (p = 0.0308, n = 9). The Aγ/α ratio in the case of GGHI-mB-3D/VSVG reached 0.37 (p = 0.32, n = 8). The lack of a significant difference between BaEVRless and VSVG vector treatment is primarily due to the marked inherent variability of the Aγ/α ratio among the thalassemic patients. The highest increase was observed in patients Thal 3 and Thal 8, with HbF reaching an increase of 273% and 253%, respectively, compared to control (Fig. 4B). To the authors' knowledge, this is the highest HbF increase achieved by a γ-globin LV in a thalassemic background at a MOI of 50, suggesting that GGHI-mB-3D/BaEVRless vector can be considered as a vector with promising therapeutic potential in a clinical setting.
The above statistically significant and therapeutic HbF levels achieved by GGHI-mB-3D/BaEVRless vector correlated with an increase at the mRNA level. According to the experimental evidence obtained from β-thalassemia mouse models, γ-globin mRNA levels around 13% of the endogenous α-globin result in a protein equivalent of 27%, thus leading to nearly complete phenotypic correction. 40 In this study, transduction with GGHI-mB-3D/BaEVRless led to an average increase of γ-globin mRNA level of 44 ± 150% relative to the endogenous α-globin (Fig. 5C), which in turn led to high HbF protein levels.
Regarding erythropoiesis and phenotypic correction, a higher trend toward reduction of apoptosis was observed with GGHI-mB-3D/BaEVRless-derived HbF expression (Pearson r = −0.55, p = 0.128). More specifically, transduction with GGHI-mB-3D/BaEVRless resulted in a marked decrease of apoptotic cells from 58.4% observed in the mock-transduced to 46.4% in GGHI-mB-3D/BaEVRless-transduced cells. According to this trend, it should be noted that a significant decrease in apoptosis was also observed in patients Thal 3, Thal 7, and Thal 8, who demonstrated a marked HbF increase. These results were further corroborated by a concomitant decrease in the ATG5 mRNA expression (Fig. 6D), a key molecule switch linking the autophagy and apoptosis pathway, as reviewed by Yousefi and Simon, 43 suggesting that the observed decrease in apoptosis is related to a decrease in autophagy initiation known to be enhanced in thalassemia. 46 Although the results did not show a statistically significant decrease of apoptosis in transduced cells compared to mock-transduced cells, it should be noted that the observed high variability, which reduces the power of group-wise comparisons, is in line with recent reports, 47,48 which indicated that apoptosis in later stages of erythroblast maturation may not always be increased in thalassemic samples.
Tightly linked with the results from apoptosis and necrosis detection by Annexin-V/7AAD, orthochromatic erythroblast counts of cytocentrifugation samples were also indicative of a trend toward improvement of erythropoiesis by GGHI-mB-3D/BaEVRless.
At the clonal level and based on BFUe colonies, the mean increase observed in transduced clones employing HPLC was greater than the one seen in erythroid liquid cultures, which represent a mixture of transduced and untransduced cells. Specifically, GGHI-mB-3D/VSVG and GGHI-mB-3D/BaEVRless LVs led to a mean HbF increase normalized to a mean VCN per patient of 214% (n = 3) and 776% (n = 5), respectively. Specifically, for GGHI-mB-3D/BaEVRless, the highest increase, reaching 10-fold, was achieved in patient Thal 9. Comparing HPLC results obtained from both liquid and methylcellulose erythroid cultures, it was observed that although GGHI-mB-3D/BaEVRless outperformed the corresponding VSVG-pseudotyped vector in most cases, the highest HbF increase was not observed in the same patients for each vector. The latter does not come as a surprise, taking into account that pooled BFUe from each patient may not contain the same number of transduced cell clones, and therefore the HbF increase outcome can vary. Also, due to cell number limitations, BFUe from fewer patients were subjected to HPLC, and therefore the mean HbF increase could be easily overestimated in each case.
The average VCN calculated for all the novel vectors was around 1.0, as mentioned above, reflecting a near-ideal value for a clinical setting. Specifically, GGHI-mB-3D/VSVG vector led to a mean VCN of 0.67 and GGHI-mB-3D/BaEVRless vector to a mean VCN of 0.86. This is not the first time that a mean VCN <1.0 has been reported in the thalassemic setting. There are a number of reports in the literature involving gene therapy experiments on thalassemia patients that demonstrated a VCN <1.0. Specifically, Roselli et al. 49 used thalassemic CD34+ cells and transduced them with GLOBE β-globin LV, demonstrating an average VCN per cell of 1.6. In this study, however, 4/22 patients had a VCN per cell of <1.0, ranging from 0.4 to 0.9 in colony-forming units. Also, in the conducted thalassemia clinical trial organized by the group of Philippe Leboulch, VCN was calculated by qPCR on pooled in vitro colony-forming cells 27 months post transplantation, and gave an average readout of 0.14 copies/cell. 7 Moreover, in another study involving transduction of CD34+ cells derived from three thalassemic donors by a γ-globin LV, 13 the average VCN was 0.73. Also, Breda et al. 9 used the AnkT9W vector to transduce CD34+ cells from the peripheral blood of β0/β0, β0/β+, and β+/β+ thalassemic patients, resulting in a VCN of 0.9. Furthermore, in the studies by Karponi et al., 11 the average VCN calculated in pooled BFUe was 0.67. Lastly, in our previous study, 14 transduction of thalassemic CD34+ cells with the GGHI γ-globin vector resulted in a VCN <1.0 in 55% of the patient cohort.
The novel vector GGHI-mB-3D/BaEVRless demonstrated significantly increased transduction efficiency compared to the GGHI-mB-3D/VSVG vector and at a MOI of 50 reached the levels achieved by the latter at a MOI of 100 (author's unpublished results). Furthermore, the GGHI-mB-3D/BaEVRless vector displayed significantly increased transduction efficiency, accompanied by significant Aγ/a ratio increase, exhibiting a moderate correlation coefficient (Pearson's r = 0.55, p = 0.125) between transduction efficiency and γ-mRNA levels. With regards to transduction and VCN, although there was a good correlation (Pearson's r = 0.67, p = 0.0465), the highest VCN was not observed in the patient with the highest transduction. Also, the study failed to detect the highest VCN in patients Thal 3 and Thal 8 who showed the greatest HbF increase, suggesting that expression from GGHI-mB-3D is not entirely dependent on the vector per se but may also be influenced by the site of integration, an observation also reported previously. 14
In conclusion, this study successfully generated and characterized a novel γ-globin vector, designated GGHI-mB-3D/BaEVRless, which combines (1) novel regulatory elements that lead to stable and high levels of HbF expression and (2) an alternative envelope glycoprotein, which mediates increased transduction efficiency in CD34+ cells. Both novel features led to improved erythropoiesis in vitro in a thalassemic background of CD34+ cells at a MOI of 50. Moreover, the study demonstrates—in direct comparison to the widely used VSVG glycoprotein—the benefit of BaEVRless pseudotyping, specifically in a β-thalassemia setting, which has significant implications for the efficient transduction and clinical efficiency of the currently ongoing clinical trials employing lentiviral transfer vectors with conventional features.
Footnotes
Acknowledgments
This research was supported by the European Research Projects on Rare Diseases, E-RARE 2nd Joint Call, Improvements of vector technology and safety for gene therapy of thalassemia (GETHERTHAL; grant no. 11E-RARE-09-155) from the European Commission to N.P.A. We would like to thank Luigi Naldini and Giuliana Ferrari for providing the initial lentiviral plasmids.
Author Disclosure
The authors have no conflicts of interest and have nothing to disclose.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.