
Abstract
Idiopathic pulmonary fibrosis (IPF) is a fibroproliferative disorder with limited therapeutic options. An aberrant wound healing process in response to repetitive lung injury has been suggested for its pathogenesis, and a number of cytokines including transforming growth factor β1 play pivotal roles in the induction and progression of fibrosis. Thus, the regulation of these pro-inflammatory conditions may reduce the progression of IPF and ameliorate its symptoms in patients. Interleukin-10 (IL-10), a pleiotropic cytokine, exerts anti-inflammatory and anti-fibrotic effects in numerous biological settings. In the present study, we investigated the preventive effects of IL-10 on bleomycin-induced pulmonary fibrosis in mice with the continuous expression of this cytokine via an adeno-associated virus serotype 6 vector. Mice were administered the adeno-associated virus serotype 6 vector encoding mouse IL-10 by intratracheal injection, and osmotic minipumps containing bleomycin were subcutaneously implanted seven days later. Lung histology and the expression levels of pro-inflammatory cytokines and fibrogenic cytokines were then analyzed. In mice exhibiting persistent IL-10 expression on day 35, the number of infiltrated inflammatory cells and the development of fibrosis in lung tissues were significantly reduced. Increases in transforming growth factor β1 and decreases in IFN-γ were also suppressed in treated animals, with changes in these cytokines playing important roles in the pathogenesis of pulmonary fibrosis. Furthermore, IL-10 significantly improved survival in bleomycin-induced mice. Our results provide insights into the potential benefit of the anti-fibrotic effects of IL-10 as a novel therapeutic approach for IPF.
Introduction
Idiopathic pulmonary fibrosis (IPF) is a chronic, progressive, and devastating disease of unknown etiology. 1 Its prognosis is poor, with a mean survival of 3–5 years after diagnosis. 2 Although the underlying mechanisms have not yet been elucidated in detail, the following pathogenic events are accepted: constant bronchoalveolar epithelial injury by various agents activates epithelial cells to secrete a number of chemical mediators and cytokines such as transforming growth factor (TGF)-β1. 3 These molecules induce the migration, proliferation, and activation of fibroblasts (termed myofibroblast differentiation), followed by destruction of the lung parenchyma with an excessive fibrillary extracellular matrix. 3 These processes are collectively called “abnormal lung tissue remodeling” and result in IPF and the loss of gas exchange function. The pivotal roles of immune cells were previously assumed in the pathogenesis of IPF, and thus, immune suppressive agents such as corticosteroids were administered to patients. 4 However, current evidence suggests more important roles for the fibrosing process in IPF than inflammation. Therefore, anti-fibrotic drugs such as pirfenidone and nintedanib have been approved for the treatment of IPF. These drugs inhibit TGF-β1 and tyrosine kinases respectively and have been shown to slow down disease progression. 1 Nevertheless, neither of these antifibrotic drugs has the ability to reverse the course of the disease, and more effective strategies are awaited.
As a potential negative regulator of inflammation and fibrosis, we have focused on interleukin-10 (IL-10). IL-10 was initially described as a cytokine synthesis inhibitory factor with anti-inflammatory functions. 5,6 This interleukin is classified as a type-2 helper T (Th2) cytokine that is secreted by a number of cells such as macrophages, monocytes, type-1 helper T (Th1) cells, Th2 cells, B cells, mast cells, dendritic cells, and regulatory T cells. 7 We previously reported that IL-10 gene transfer significantly reduced damage to various organs and improved survival rates in several animal disease models such as pulmonary arterial hypertension, stroke-prone spontaneous hypertension, and peritoneal fibrosis. 8 –10 Collectively, these findings suggest that IL-10 reduces fibrotic changes in various organs, including the lung, heart, and kidney, by inhibiting TGF-β1. 8 –11
IL-10 must be maintained at an appropriate level in the target tissue in order to exert its anti-fibrotic functions in the lung; a high concentration of the cytokine in the systemic circulation may cause undesirable adverse events. Therefore, an efficient gene transfer vehicle and targeting strategy to the lung are mandated. We recently optimized adeno-associated virus (AAV) vector–mediated gene transfer to the respiratory tract and improved transduction efficiency over that reported previously. 12 An optimal AAV serotype for respiratory cells was AAV6, and the cytomegalovirus early enhancer/chicken β-actin (CAG) promoter achieved robust and sustained expression in respiratory cells. Regarding the route of administration, an intratracheal injection was more suitable for targeting the lung than other methods, and the delivered AAV6-CAG vector efficiently transduced pericytes in the lung parenchyma. This is of interest because pericytes constitute a source of myofibroblasts, the activation and proliferation of which have been suggested to play a pivotal role in lung fibrosis, and thus, may be an appropriate therapeutic target for IPF.
In the present study, we examined the effects of IL-10, delivered via an AAV6-CAG vector, in a mouse model of pulmonary fibrosis induced by bleomycin. We also investigated the molecular events potentially involved in fibrotic changes in the lung and the effects of IL-10.
Materials and Methods
AAV vector for mouse IL-10 expression
Mouse IL-10 (mIL10) was cloned from the pCMVmIL10 plasmid 13 by a polymerase chain reaction (PCR) using the following primers: forward, 5′- GGATCCGCCACCATGCCTGGCTCAGCAC-3′, and reverse, 5′- CTCGAGTTAGCTTTTCATTTTGATCATCATG-3′. The 540-base pair PCR product was cloned into the pMD20-T vector (Takara Bio, Shiga, Japan) using the Mighty TA-cloning Reagent Set for PrimeSTAR (Takara Bio). Subcloned mIL10 cDNA was transferred to pAAV-CAG-Luc, 12 replacing the luciferase gene. The resultant AAV vector plasmid (AAV-CAG-mIL10) contains the cytomegalovirus early enhancer/chicken β-actin (CAG) promoter, mIL10 gene, and AAV2-derived inverted terminal repeats. The recombinant AAV6-CAG-mIL10 vector was prepared as previously described. 14 Briefly, 60% confluent human embryonic kidney cells (HEK293; Agilent Technologies, Palo Alto, CA) were incubated in large culture vessels and cotransfected with an AAV vector plasmid (either pAAV-CAG-mIL10 or pAAV-CAG-Luc), an adenoviral helper plasmid pHelper (Agilent Technologies), and an AAV6 chimeric helper plasmid (pAAV2 Rep/AAV6 Cap). 15 Crude viral lysates were purified twice on a CsCl two-tier centrifugation gradient. Viral titers were measured by quantitative PCR, and the viral stock was dissolved in HEPES-NaCl (HN) buffer [50 mmol/L 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid, pH 7.4, 150 mmol/L NaCl] before the injection.
Human bronchial epithelial cells (BEAS-2B; American Type Culture Collection, Manassas, VA) were maintained in serum-free bronchial epithelial cell growth medium (BEGM; Lonza, Walkersville, MD) supplemented with a bullet kit (Lonza) and were kept in an incubator at 37°C with 5% CO2. BEAS-2B cells were seeded on 96-well plates at a density of 2 × 104 cells/well 24 h prior to transduction. Cells were transduced with AAV6-CAG-mIL10, AAV6-CAG-Luc at 3 × 105 viral genomes (vg)/cell, or no vector treatment. After a 48-h incubation, each conditioned medium was recovered and centrifuged at 3,000 g at 4°C for 10 min. The concentration of mIL10 in the supernatant was assessed using an enzyme-linked immunosorbent assay (ELISA) (mouse IL-10 ELISA; eBioscience, San Diego, CA).
Animal models
All animal experiments were performed in accordance with the guidelines issued by the Jichi Medical University (Tochigi, Japan) Ethics Committee. Male C57BL/6J mice aged 10–12 weeks (from SLC Japan, Shizuoka, Japan) weighing 25–30 g were used. Mice were randomly divided into four groups of 6–10 mice each (Fig. 1): mice administered saline after the HN buffer (group 1, no vector + saline); mice administered saline after AAV6-CAG-mIL10 (group 2, AAV-IL10 + saline); mice administered bleomycin after the HN buffer (group 3, no vector + bleomycin); and mice administered bleomycin after AAV6-CAG-mIL10 (group 4, AAV-IL10 + bleomycin). Mice were injected with HN buffer (50 μL) or AAV6-CAG-mIL10 (5 × 1010 vg/mouse, 50 μL) intratracheally on day 0. The intratracheal injection was performed as previously described. 16 Under anesthesia, mice were placed and secured in a supine position and a small incision was made in the midline of the neck. The 29-gauge needle was inserted between the cartilaginous rings of the trachea and HN buffer or rAAV vector was injected. The skin incision was then sutured, and the mice were allowed to recover. Seven days later (on day 7), osmotic minipumps (Alzet 2001; DURECT, Cupertino, CA) containing 200 μL of sterile saline solution with or without 125 mg/kg of bleomycin (Nippon Kayaku, Tokyo, Japan) were implanted subcutaneously on the left side of the back. The minipumps were adjusted to deliver their contents at 1 μL/h for 7 days. The body weights of mice were measured every other day or every 3 days, and mice were sacrificed at the indicated time points.
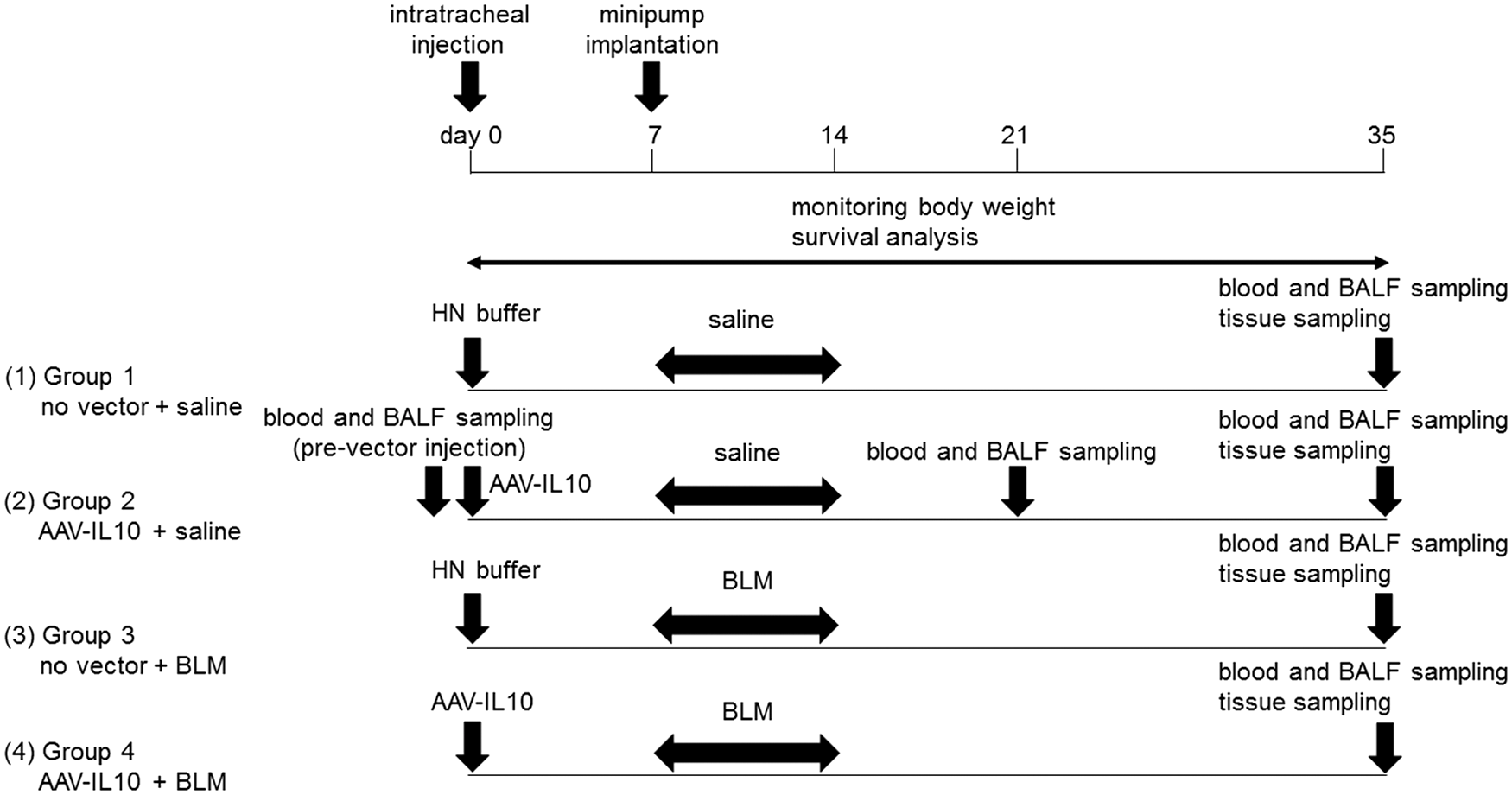
Experimental protocol. Male C57BL/6J mice aged 10–12 weeks were injected intratracheally with AAV6-CAG-mIL10 (5 × 1010 vg/ mouse) or vehicle (HN buffer) on day 0. From day 7, mice were continuously treated with 125 mg/kg of bleomycin or saline for one week using subcutaneously implanted osmotic minipumps. Mice were randomly divided into four groups as follows: group 1, no vector + saline ; group 2, AAV-IL10 + saline ; group 3, no vector + BLM ; and group 4, AAV-IL10 + BLM. As part of the time-course experiment, mice were sacrificed on days 0, 21, and 35, and samples such as blood, BALF, and lung tissue were collected for further analysis. AAV, adeno-associated virus; CAG, cytomegalovirus early enhancer/chicken β-actin promoter; mIL10, mouse interleukin-10; BLM, bleomycin; HN buffer, HEPES-NaCl buffer [50 mmol/L 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid, pH 7.4, 150 mmol/L NaCl]; BALF, bronchoalveolar lavage fluid.
In order to examine the time course of mIL10 expression, blood and bronchoalveolar lavage fluid (BALF) were collected on days 0 (prevector injection), 21, and 35 in group 2 mice (AAV-IL10 + saline group). The concentrations of mIL10 in plasma and BALF were measured using mouse IL-10 ELISA (eBioscience).
BALF collection and histology
Mice were sacrificed on day 35 and BAL was performed by cannulating the trachea with a 20-gauge catheter and completely infusing the lungs four times with 0.8 mL of cold phosphate-buffered saline. The recovery of BALF ranged between 2.4 and 3.0 mL per mouse. BALF was centrifuged at 1,000 g at 4°C for 10 min, and supernatants were collected. The number of white blood cells was counted using a hemocytometer, and a differential cell analysis was performed by Diff-Quick staining (Sysmex, Kobe, Japan) after cytospinning at 800 rpm at 22°C for 10 min.
After lavage on day 35, the lungs were fixed by an intratracheal injection of 1 mL of 10% formalin neutral buffer solution (Wako, Osaka, Japan) and embedded in paraffin. Left lung sections were cut into 3-μm-thick slices, which were then stained with hematoxylin and eosin or Masson's trichrome. The degree of fibrosis was scored using the Ashcroft method 17 and 20 random fields per lung were assessed at a magnification of 100 × . The grade of lung fibrosis was scored between 0 and 8: grade 0, normal lung; grade 1, minimal fibrous thickening of the alveolar or bronchial walls; grade 3, moderate thickening of the walls without obvious damage to the lung architecture; grade 5, increased fibrosis with definite damage to the lung structure and the formation of fibrous bands or small fibrous masses; grade 7, severe distortion of the structure and large fibrous areas; grade 8, total fibrous obliteration of the field.
Measurement of cytokines and collagen in mouse lungs
BALF and lung homogenates on day 35 were analyzed for cytokines and collagen in treated mouse lungs. Each right lung was homogenized in 1,000 μL of sterile saline solution and centrifuged at 10,000 g at 4°C for 10 min, and the supernatants were assayed by ELISA. The levels of the following cytokines: IL-1β, IL-6, tumor necrosis factor (TNF)-α, and interferon (IFN)-γ, were measured in BALF or lung homogenates using mouse IL-1β ELISA (eBioscience), mouse IL-6 ELISA (eBioscience), and mouse TNF-α ELISA (eBioscience), and mouse IFN-γ ELISA (eBioscience), respectively. In order to measure total TGF-β1 levels, BALF was acid treated and neutralized according to the manufacturer's instructions because we used mouse TGF-β1 ELISA (eBioscience). Soluble collagen levels in BALF were assessed using the Sircol Soluble Collagen Assay (Biocolor, Belfast, UK) according to the manufacturer's instructions.
Statistical analysis
Data are summarized as the mean ± standard error of the mean. Regarding ELISA, undetectable levels were defined as zero. Comparisons among multiple groups were made using a one-way analysis of variance with Tukey's correction or Games-Howell's correction, and a survival analysis was performed using the Kaplan-Meier method and compared by log-rank test. Statistical analyses were performed with SPSS software (IBM Japan, Tokyo, Japan). In all tests, p < 0.05 was considered to denote significance.
Results
Transduction of the mouse IL-10 gene in vitro and in vivo
We initially examined the production of mouse IL-10 (mIL10) from the AAV6-CAG-mIL10 vector in vitro. Human bronchial epithelial cells (BEAS-2B) were transduced with AAV6-CAG-mIL10 or AAV6-CAG-Luc. 12 The concentration of mIL10 in the conditioned medium was ca. 300 ng/mL (Fig. 2A). This concentration was higher than that of our previous AAV vector in which the CMV promoter drove mIL10 (ca. 80 ng/mL); 13 however, a direct comparison was difficult due to differences in the cell type and culture condition.
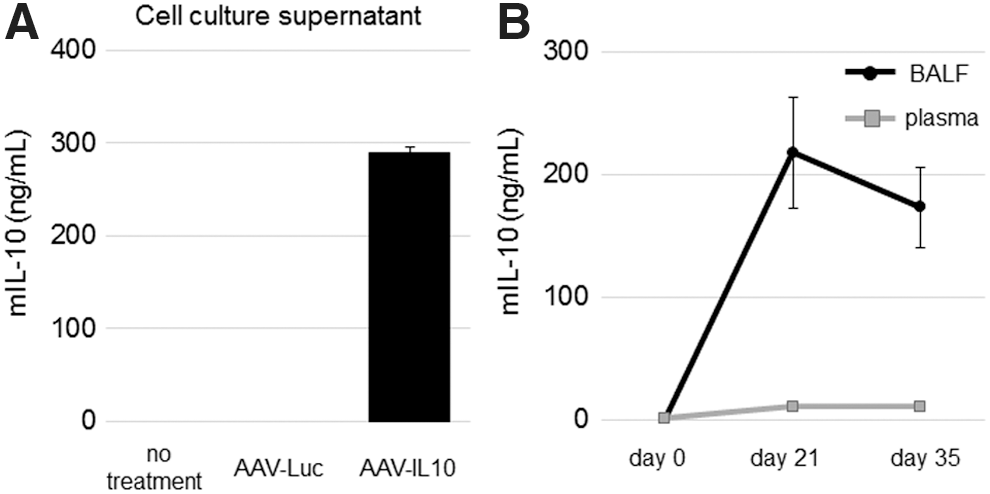
Vector-derived mIL10 in vitro and in vivo.
Vector-derived mIL10 production was also confirmed in vivo. The time course of mIL10 expression after the intratracheal injection of the AAV6-CAG-mIL10 vector was followed in mice (group 2, AAV-IL10 + saline, n = 6). Blood and BALF were collected on days 0 (pre-vector injection), 21, and 35. The mIL10 concentration in BALF was very high on day 21 (217 ± 45 ng/mL) and was maintained until day 35 (172 ± 32 ng/mL), comparable to that in the conditioned medium of BEAS-2B. In contrast, the mIL10 level was 20-fold lower in plasma (10 ± 2 ng/mL) (Fig. 2B). Taken together with our previous findings, 12 the intratracheal injection of the recombinant AAV (rAAV) vector is suitable for targeting the lung.
Prevention of bleomycin-induced pulmonary fibrosis by a rAAV encoding mIL10 in mice
We investigated whether the mIL10-encoding AAV vector prevents pulmonary fibrosis in mice. As a mouse disease model, bleomycin-induced pulmonary fibrosis has been well established, in which the drug is delivered to the trachea by a bolus injection or subcutaneously with a continuous osmotic minipump. 18,19 The former induces entire lung injury, followed by severe lung fibrosis with a bronchocentric distribution. As a result, this method is associated with marked weight loss and considerable mortality. The latter method with minipumps induces mild to moderate fibrosis similar to human IPF, in which heterogeneous lesions are distributed in the distal area and are often located in the subpleural compartment with vascular involvement. Therefore, a continuous subcutaneous injection of bleomycin is associated with milder weight loss and a lower mortality rate, 18,19 and we employed this method to prepare lung fibrosis in mice in subsequent experiments.
Prior to the bleomycin infusion, mice were administered either AAV6-CAG-mIL10 or vehicle. Seven days later, a 7-day course of subcutaneous bleomycin or saline infusion was initiated in each group, and the 4 cohorts of mice were observed for the next 3 weeks (groups 1–4 in Fig. 1). Regarding survival, we only lost 3 mice (out of 10) in group 3 (no vector + bleomycin), suggesting the moderate morbidity of the bleomycin treatment (Fig. 3A). No mouse in the other groups died and this difference was significant (p = 0.018; data were analyzed by log-rank test).
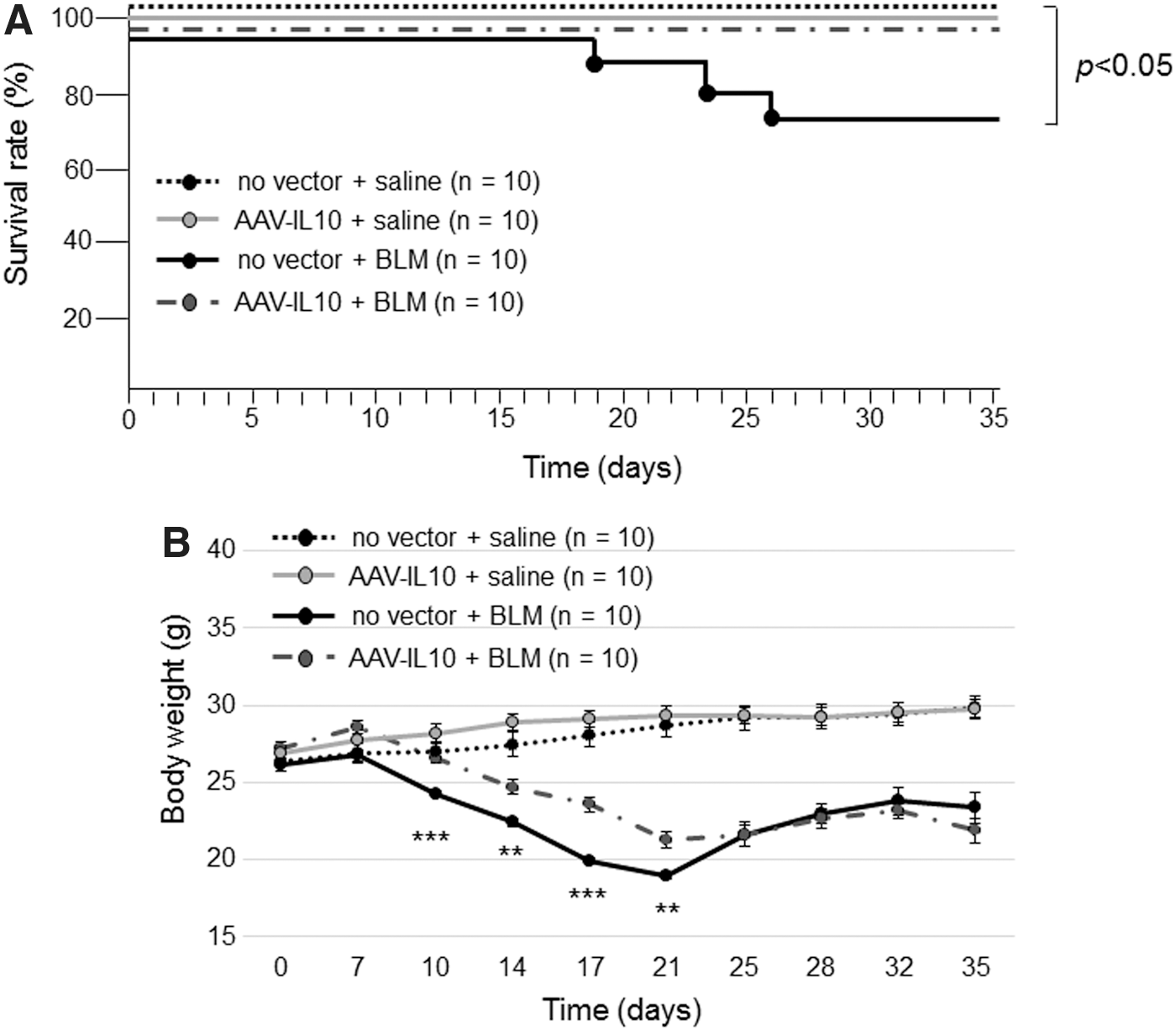
Effects of AAV-mediated mIL10 expression on body weight and survival rate in mice treated with bleomycin. Mice were injected intratracheally with AAV6-CAG-mIL10 (5 × 1010 vg/ mouse) or vehicle (HN buffer) on day 0. On day 7, 125 mg/kg of bleomycin or saline was given using osmotic minipumps. No vector + saline mice, AAV-IL10 + saline mice, no vector + BLM mice, and AAV-IL10 + BLM mice (n = 10 for each group) were examined.
Average body weights at baseline (day 0) did not significantly differ among the four groups, and saline-treated mice gained weight throughout the experiment without a significant difference. Bleomycin-treated mice lost weight during drug administration and the initial week of the observation period, with the degree of weight loss being milder in mice pretreated with AAV6-CAG-mIL10 (group 4) than in those pretreated with vehicle (group 3) (Fig. 3B). The body weight difference between these two cohorts was significant from days 10 to 21. Bleomycin-treated mice (group 3 and group 4) gradually recovered their body weight, and the difference between the two groups was not significant in the last 2 weeks of the observation period.
Three weeks after the bleomycin treatment (on day 35 in Fig. 1), mice were sacrificed for histological and biochemical examinations. Lung sections were prepared in order to evaluate inflammatory responses and fibrotic changes (Fig. 4A). Moderate to severe inflammatory infiltration in the peribronchiolar and perivascular areas was observed in group 3 (no vector + bleomycin), while lung tissues from group 4 (AAV-IL10 + bleomycin) exhibited less inflammatory cell infiltration than that in group 3. No significant inflammation was observed in group 1 (no vector + saline) or group 2 (AAV-IL10 + saline).
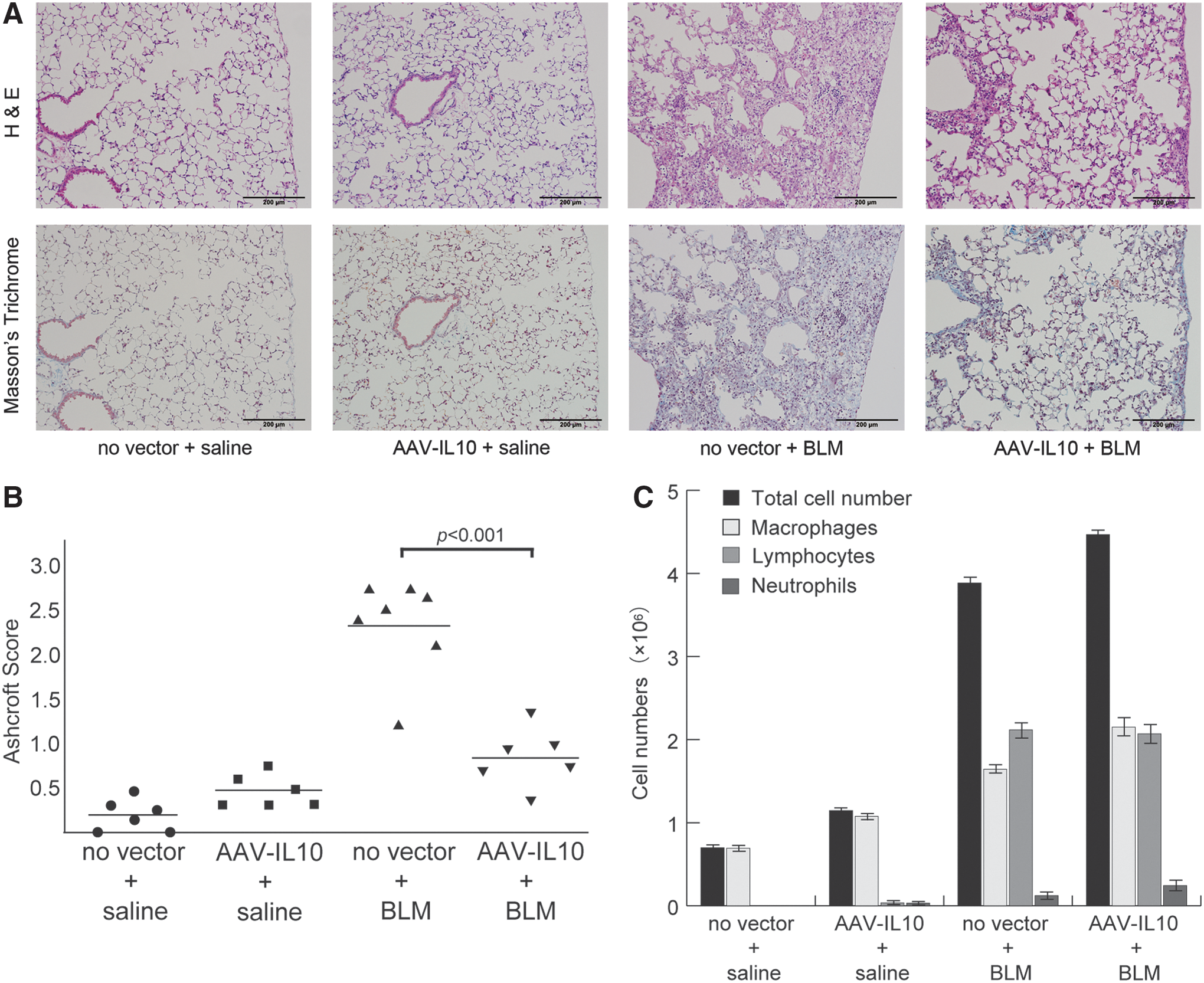
Anti-inflammatory and anti-fibrotic effects of mIL10 on bleomycin-induced pulmonary fibrosis. Mice were injected intratracheally with AAV6-CAG-mIL10 (5 × 1010 vg/ mouse) or vehicle (HN buffer) on day 0. On day 7, 125 mg/kg of bleomycin or saline control was given using osmotic minipumps. No vector + saline mice (n = 6), AAV-IL10 + saline mice (n = 6), no vector + BLM mice (n = 7), and AAV-IL10 + BLM mice (n = 6) were analyzed on day 35.
Masson's trichrome staining revealed marked fibrotic changes in mice in Group 3 (no vector + bleomycin), with destruction of the alveolar architecture and interstitial fibrosis in subpleural regions. Lung tissues from group 4 mice (AAV-IL10 + bleomycin) exhibited less fibrotic changes. In saline-treated control mice (groups 1 and 2), almost no fibrosis was observed in the lungs on day 35. The extent of lung fibrosis was quantified by the Ashcroft method (Fig. 4B). The baseline Ashcroft score was 0.21 ± 0.07 (group 1, n = 6), and the pretreatment with AAV6-CAG-mIL10 slightly increased the fibrotic score in group 2 (0.48 ± 0.06, n = 6), although the difference was not significant (p = 0.5). Bleomycin induced moderate lung fibrosis in mice, with an Ashcroft score of 2.3 ± 0.2 (group 3, n = 7), whereas the pretreatment with AAV6-CAG-mIL10 significantly lowered the fibrotic score in group 4 (0.84 ± 0.13, n = 6; p < 0.001).
Inflammatory responses
In order to evaluate the effects of the AAV6-CAG-mIL10 vector on inflammatory responses induced by bleomycin, BAL was obtained on day 35 (Fig. 4C). The numbers of inflammatory cells such as macrophages, lymphocytes, and neutrophils were significantly higher in bleomycin-treated mice (group 3: no vector + bleomycin, and group 4: AAV-IL10 + bleomycin) than in saline-treated control mice (group 1: no vector + saline, and group 2: AAV-IL10 + saline). The pretreatment with the AAV vector did not affect inflammatory cells in BALF; the total cell number and differential cell count were similar in bleomycin-treated animals regardless of whether they were given vehicle (group 3) or AAV-IL10 (group 4).
The levels of pro-inflammatory cytokines such as IL-1β, IL-6, and TNF-α in BALF or the supernatants of lung homogenates were assayed on day 35. The concentrations of IL-1β and TNF-α in BALF were undetectable in all cohorts (Supplementary Table S1; Supplementary Data are available online at
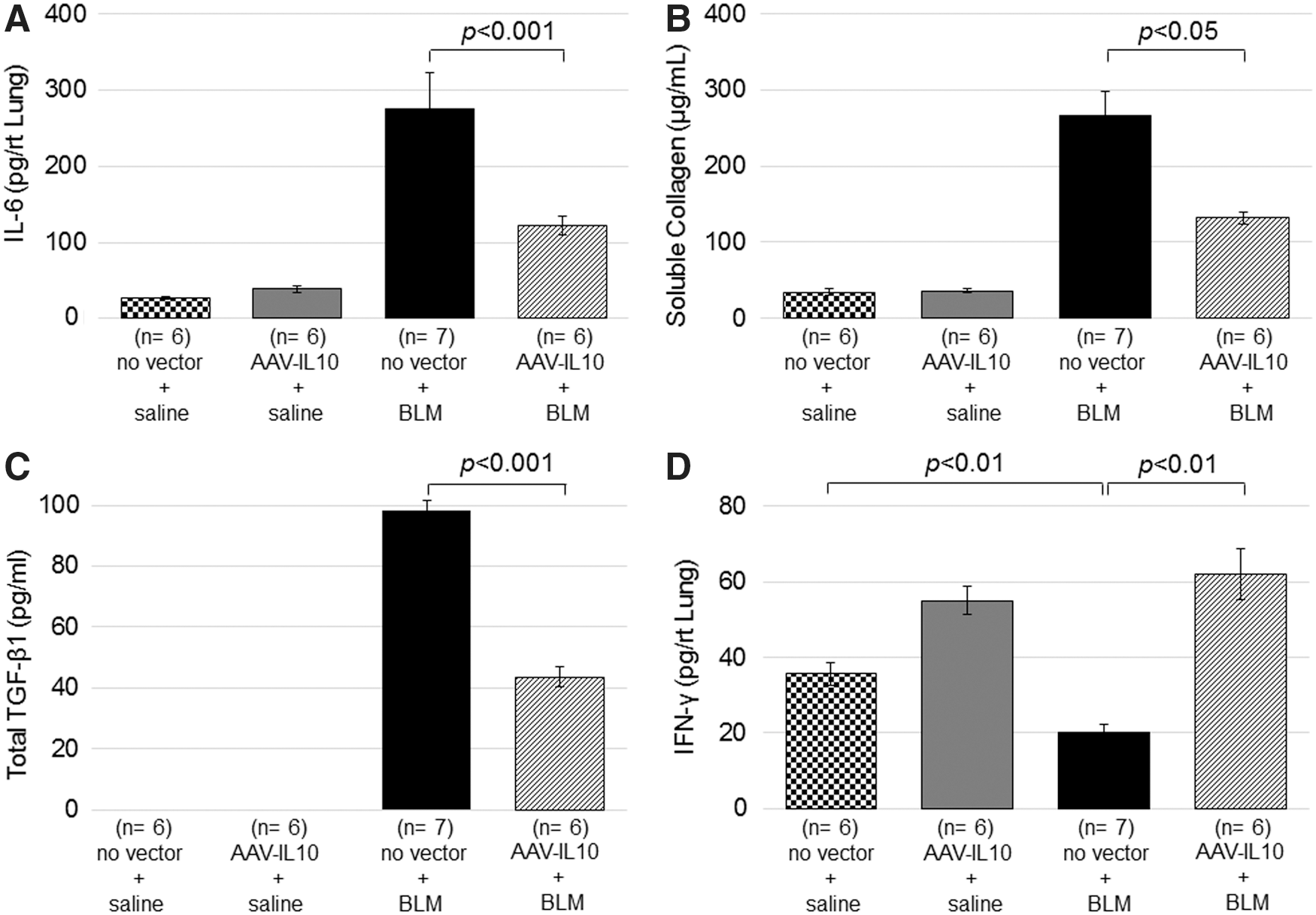
Effects of mIL10 on collagen contents and IL-6, TGF-β1, and IFN-γ levels in the lung. Mice were injected intratracheally with AAV6-CAG-mIL10 (5 × 1010 vg/ mouse) or vehicle (HN buffer) on day 0. On day 7, 125 mg/kg of bleomycin or saline control was given using osmotic minipumps. No vector + saline mice (n = 6), AAV-IL10 + saline mice (n = 6), no vector + BLM mice (n = 7), and AAV-IL10 + BLM mice (n = 6) were analyzed on day 35.
Fibrotic responses
As observed in the lung parenchyma, in order to evaluate the degree of bleomycin-induced fibrosis, the suppression of collagen production by AAV6-CAG-mIL10 was also confirmed in the BALF analysis. The collagen content in BALF on day 35 was analyzed using the Sircol Soluble Collagen Assay (Fig. 5B). The collagen content increased after the injection of bleomycin (group 3: no vector + bleomycin, and group 4: AAV-IL10 + bleomycin), and it was significantly reduced by the administration of the AAV6-CAG-mIL10 vector (group 4). In saline-treated control mice (group 1: no vector + saline; and group 2: AAV-IL10 + saline), the collagen content in BALF was not increased.
TGF-β1 has been reported to play pivotal roles in the development of pulmonary fibrosis. The expression of total TGF-β1 in BALF on day 35 was analyzed by ELISA after complete activation by acidification (Fig. 5C). Total TGF-β1 levels increased after the injection of bleomycin (group 3: no vector + bleomycin; and group 4: AAV-IL10 + bleomycin) and were significantly suppressed by the administration of the AAV6-CAG-mIL10 vector (group 4 mice). In saline-treated control mice (group 1: no vector + saline, and group 2: AAV-IL10 + saline), TGF-β1 levels were not increased.
In addition to its immunoactivating roles, IFN-γ is known to have an anti-fibrotic function. IFN-γ levels in BALF and lung homogenates were analyzed on day 35 (Supplementary Table S1 and Fig. 5D). With moderate fibrosis induced by bleomycin, IFN-γ in BALF decreased to an undetectable level (Supplementary Table S1) and was significantly lower in the lung homogenate (Fig. 5D). With the administration of the AAV6-CAG-mIL10 vector, baseline IFN-γ slightly increased in the lung homogenate (no vector + saline vs. AAV-IL10 + saline), and the bleomycin-induced down-regulation of IFN-γ was completely abrogated (no vector + bleomycin vs. AAV-IL10 + bleomycin). Taken together, these results indicate that AAV vector-derived IL-10 ameliorates lung fibrosis by at least two-fold through mechanisms such as the suppression of TGF-β1 and up-regulation of IFN-γ.
Discussion
In the present study, we demonstrated that an intratracheal injection of the AAV6-CAG-mIL10 vector prevented pulmonary fibrosis in a mouse disease model and improved survival. Bleomycin induces inflammation in the lung, in which leukocytes infiltrate and pro-inflammatory cytokines such as IL-6 are overexpressed. Tissue destruction and subsequent repair processes often result in lung fibrosis, in which TGF-β1 and IFN-γ may play positive and negative roles, respectively. We showed that the pretreatment with the rAAV expressing IL-10 suppressed the aberrant expression of these cytokines (i.e., the up-regulation of IL-6 and TGF-β1 and down-regulation of IFN-γ).
We speculate that this suppression of pulmonary fibrosis involves multiple functions of IL-10. In the present study, IL-10 expression suppressed inflammatory cell activation in the lungs of bleomycin-treated mice and subsequent IL-6 elevations, which may account, in part, for the preventive effect observed against pulmonary fibrosis. However, the lack of evidenced efficacy of the anti-inflammatory therapy for IPF suggests that the suppression of IL-6 is not a central role of IL-10 in preventing the progression of IPF. 22,23 A major mode of action of IL-10 may instead be the suppression of exaggerated wound healing in which TGF-β1 functions as a key pro-fibrotic cytokine. 24 TGF-β1, which is secreted from a number of cell types in the lung such as alveolar macrophages, smooth muscle cells, fibroblasts, and airway and alveolar epithelial cells, 25,26 up-regulates fibrillary collagens and extracellular matrix proteins through Smad-mediated signaling 27,28 and induces epithelial-mesenchymal transition via Smad-independent signaling pathways. 29 Taken together with our previous findings, the present results demonstrate that IL-10 significantly suppresses the up-regulation of TGF-β1, resulting in milder fibrotic changes and tissue damage in many organs such as the heart, kidney, peritoneum, and lung. 8 –11 These results suggest that the suppression of TGF-β1 is one of the major mechanisms responsible for the anti-fibrotic effects of IL-10. In addition to direct transcriptional regulation, 10 IL-10 may antagonize TGF-β1 via IFN-γ signaling. IFN-γ, a Th1-type cytokine, inhibits transcriptional responses to TGF-β signaling, known as an anti-fibrotic cytokine. 30 In the present study, endogenous lung IFN-γ levels decreased during the development of bleomycin-induced pulmonary fibrosis, and IL-10 gene transfer prevented further decreases in this down-regulation. Similarly, IFN-γ levels in IPF patients were low, 31,32 and these findings prompted clinical trials to administer IFN-γ to these patients. 33 –36 These trials showed that airway inhalation is a preferable method over subcutaneous injection, and support the idea that maintaining lung IFN-γ levels by local IL-10 production may be beneficial in IPF patients.
The efficacy of IL-10 gene transfer in bleomycin-treated mice has been discussed previously. 37,38 Arai et al. injected liposome-conjugated IL-10 plasmids intraperitoneally and found lower myeloperoxidase, TNF-α mRNA, and hydroxyproline levels than those in control mice treated with bleomycin. 37 Nakagome et al. moderately suppressed lung fibrosis in bleomycin-treated mice by hydrodynamic-based IL-10 plasmid transduction. 38 However, the expression of vector-derived IL-10 in these studies was only transient, and the long-term impact on disease progression was not pursued. In contrast, our intratracheal injection of AAV6-CAG-mIL10 resulted in the continuous and relatively lung-specific expression of IL-10 associated with a better physical outcome. In addition, the absence of baseline inflammation or fibrosis in mice given AAV6-CAG-mIL10 suggested the safety of an intratracheal injection of this rAAV vector.
One of the limitations of the present study is that the design only allowed us to show the protective, not curative effect of the vector. Nevertheless, our results indicate some benefits with the administration of the vector in later stages of IPF because of its anti-fibrotic effects through the regulation of TGF-β1 and IFN-γ. It currently remains unclear whether the administration of an IL-10-expressing vector is beneficial for patients with ongoing lung fibrosis, and we are now preparing for such an experiment.
In summary, the AAV6-CAG-mIL10 vector successfully prevented bleomycin-induced pulmonary fibrosis and prolonged survival in mice. These results suggest that our AAV6 vector-mediated IL-10-based strategy has the potential to prevent the progression of pulmonary fibrosis in humans and will contribute to a novel therapeutic approach for IPF.
Footnotes
Acknowledgments
We thank Miyoko Mitsu and Satomi Fujiwara for their technical assistance and Drs. Teruhiko Kanabayashi and Tomomichi Kanabayashi (Biopathology Institute) for their generous help in analyzing lung section slides. This work was supported in part by a grant from the Jichi Medical University Young Investigator Award to F.K. This study was also supported by the Research Program on HIV/AIDS and the Practical Research Project for Rare/Intractable Diseases from the Japan Agency for Medical Research and Development.
Author Disclosure
One of the authors (R.U.) is supported by Takara Bio, Inc. The other authors have no conflicts of interest to declare.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.