
Abstract
The tumor-homing properties of mesenchymal stem cells (MSC) have led to their development as delivery vehicles for the targeted delivery of therapeutic genes such as the sodium–iodide symporter (NIS) to solid tumors. External beam radiation therapy may represent an ideal setting for the application of engineered MSC-based gene therapy, as tumor irradiation may enhance MSC recruitment into irradiated tumors through the increased production of select factors linked to MSC migration. In the present study, the irradiation of human liver cancer cells (HuH7; 1–10 Gy) showed a strong dose-dependent increase in steady-state mRNA levels of CXCL8, CXCL12, FGF2, PDGFB, TGFB1, THBS1, and VEGF (0–48 h), which was verified for most factors at the protein level (after 48 h). Radiation effects on directed MSC migration were tested in vitro using a live cell tracking migration assay and supernatants from control and irradiated HuH7 cells. A robust increase in mean forward migration index, mean center of mass, and mean directionality of MSCs toward supernatants was seen from irradiated as compared to non-irradiated tumor cells. Transferability of this effect to other tumor sources was demonstrated using the human breast adenocarcinoma cell line (MDA-MB-231), which showed a similar behavior to radiation as seen with HuH7 cells in quantitative polymerase chain reaction and migration assay. To evaluate this in a more physiologic in vivo setting, subcutaneously growing HuH7 xenograft tumors were irradiated with 0, 2, or 5 Gy followed by CMV-NIS-MSC application 24 h later. Tumoral iodide uptake was monitored using 123I-scintigraphy. The results showed increased tumor-specific dose-dependent accumulation of radioiodide in irradiated tumors. The results demonstrate that external beam radiation therapy enhances the migratory capacity of MSCs and may thus increase the therapeutic efficacy of MSC-mediated NIS radionuclide therapy.
Introduction
The sodium–iodide symporter (NIS) is an intrinsic transmembrane glycoprotein with 13 putative transmembrane domains. NIS mediates the active uptake of iodide into the thyroid gland and, as such, has formed the basis for the diagnostic and therapeutic application of radioiodine in benign and especially malignant thyroid diseases for almost 70 years. 1 Its properties as a reporter and therapy gene allow noninvasive imaging of functional NIS expression by 123I-scintigraphy and 124I-PET imaging and robust therapeutic effects by the application of 131I and related radionuclides. 1 –4 The extensive clinical experience derived from work with NIS-biology and NIS-mediated radioiodine accumulation in diagnosis and imaging of differentiated thyroid cancer has allowed expansion of this effective theranostic strategy to non-thyroidal tumor settings through the use of various delivery vehicles for tumor-targeted NIS gene expression. The approaches under evaluation include viral and non-viral vectors such as genetically engineered mesenchymal stem cells (MSC). The results to date have demonstrated tumor-selective NIS-mediated radioiodine accumulation leading to decreased tumor growth and prolonged survival of treated animals. 5 –22 MSCs are very promising potential vehicles for the delivery of therapeutic genes to tumors, as solid tumors are thought to have many of the characteristics of chronic wounds due to ongoing and extensive hypoxia, necrosis, and angiogenesis, which drive MSC recruitment. 23,24 The enhanced production of inflammatory cytokines, growth factors, and chemokines by the tumor cells is thought to underlie the directed migration of MSCs to solid tumors. Previous reports have suggested that platelet-derived growth factor (PDGF), epidermal growth factor (EGF), vascular endothelial growth factor (VEGF), interleukin-6 (IL-6), and the chemokine (C-X-C motif) ligand 12/stromal cell-derived factor 1 (CXCL12/SDF-1) play important roles in the recruitment of MSCs into tumor environments. 25 Tumor external beam irradiation may represent a promising tool to enhance MSC homing to tumor microenvironments based on the localized tissue damage seen with irradiation leading to the increased secretion of inflammatory chemokines and growth factors. 26,27
The present study sought to evaluate the effect of external beam radiation on tumor-homing of adoptively applied engineered MSCs to determine if this treatment would improve or enhance functional MSC-mediated NIS gene expression within the experimental tumors. To this end, the study investigated the effect of radiation on the chemokine and growth factor expression profile of the human hepatocellular carcinoma (HCC) cell line HuH7, as well as of the human breast adenocarcinoma cell line MDA-MB-231, and the overall effect on the directed migration of MSC. The in vitro results where then validated in a HCC xenograft mouse model taking advantage of the reporter gene function of NIS.
Materials and Methods
Cell culture
The human hepatocellular carcinoma cell line HuH7 was purchased from JCRB Cell Bank (JCRB 0403; Osaka, Japan) and cultured in Dulbecco's modified Eagle's medium (Sigma–Aldrich, St. Louis, MO) supplemented with 10% fetal bovine serum (FBS; Biochrom/Merck Millipore, Berlin, Germany) and 100 IU/mL penicillin/100 μg/mL streptomycin (Sigma–Aldrich). The human breast adenocarcinoma cell line MDA-MB-231 was cultured in RPMI (Sigma–Aldrich) supplemented with 10% FBS and 100 IU/mL penicillin/100 μg/mL streptomycin. A previously established SV40 large T antigen immortalized MSC cell line derived from human bone marrow 28 was cultured in RPMI supplemented with 10% FBS and 100 IU/mL penicillin/100 μg/mL streptomycin. Primary human bone marrow–derived CD-34-negative MSCs were cultured as described previously. 29 Cells were maintained at 37°C and 5% CO2 in an incubator.
Stable transfection and establishment of MSCs
The establishment of wild-type MSCs (WT-MSC) transfected with the expression vector CMV-NIS-pcDNA3 (full-length NIS cDNA coupled to the cytomegalovirus [CMV] promoter) and the resulting stably transfected cell line (CMV-NIS-MSC) has been described previously. 12
125I uptake assay
Radioiodide uptake of CMV-NIS-MSC was determined at steady-state conditions before in vivo application, as described previously by Spitzweg et al. 30 Results were normalized to cell survival measured by cell viability assay (see below).
Cell viability assay
Cell viability was determined as described previously. 15
Irradiation of HuH7 and MDA-MB-231 cells
HuH7 or MDA-MB-231 cells (1 × 106) were seeded on 100 mm3 surface cell culture plates. Twelve hours before radiation, cells were switched to serum-free medium. Tumor cells were irradiated with doses from 0 to 10 Gy using an Xstrahl Cabinet Irradiator RS225 (200 kV, 10 mA, Thoraeus filter, 1 Gy in 1 min 3 s; Xstrahl Ltd., Camberley, United Kingdom).
Quantitative real-time polymerase chain reaction
Total RNA from HuH7 cells was extracted after 4, 8, 12, 24, and 48 h post radiation (0–10 Gy) using the RNeasy Mini Kit with QIAshredder (Qiagen, Hilden, Germany). The same procedure was performed for MDA-MB-231 cells after 8, 24, and 48 h post radiation. Reverse transcription and quantitative real-time polymerase chain reaction (qRT-PCR) were conducted as previously described using a Mastercycler ep gradient S PCR cycler (Eppendorf, Hamburg, Germany). 13 The following primers were used: ACTB (β-actin): forward primer (5′-AGAAAATCTGGCACCACACC-3′) and reverse primer (5′-TAGCACAGCCTGGATAGCAA-3′); CXCL8 (chemokine (C-X-C motif) ligand 8): forward primer (5′-TCTGCAGCTCTGTGTGAAGG-3′) and reverse primer (5′-TTCTCCACAACCCTCTGCAC-3′); CXCL12: forward primer (5′-AGAGCCAACGTCAAGCATCT-3′) and reverse primer (5′-TAGCACAGCCTGGATAGCAA-3′); FGF2 (basic fibroblast growth factor): forward primer (5′-GGAGAAGAGCGACCCTCAC-3′) and reverse primer (5′-AGCCAGGTAACGGTTAGCAC-3′); PDGFB: forward primer (5′-TTGGCTCGTGGAAGAAGG-3′) and reverse primer (5′-CGTTGGTGCGGTCTATGA-3′); TGFB1 (transforming growth factor beta1 [TGF-β1]): forward primer (5′-CAGCACGTGGAGCTGTACC-3′) and reverse primer (5′-AAGATAACCACTCTGGCGAGTC-3′); THBS1 (thrombospondin 1 [TSP-1]): forward primer (5′-TTGTCTTTGGAACCACACCA-3′) and reverse primer (5′-CTGGACAGCTCATCACAGGA-3′); and VEGF: forward primer (5′-CTACCTCCACCATGCCAAGT-3′) and reverse primer (5′-ATGATTCTGCCCTCCTCCTT-3′). Relative expression levels were calculated from ΔΔCt values normalized to internal β-actin. Results are expressed as fold change relative to controls.
Enzyme-linked immunosorbent assay
Supernatant from untreated and irradiated HuH7 cells was removed after 4, 8, 12, 24, or 48 h, centrifuged and stored at −80°C. Enzyme linked immunosorbent assays (ELISA) for CXCL8, CXCL12/SDF-1, FGF2, PDGF beta homodimer (PDGF BB), TGF-β1, TSP-1, and VEGF were performed using the respective DuoSet ELISA kit (R&D Systems, Abington, United Kingdom) following the manufacturer's instructions. Results are expressed as fold change relative to controls.
Cell migration assay
Chemotaxis of MSCs in relation to a gradient between irradiated and non-irradiated supernatants of HuH7 or MDA-MB-231 cells (48 h post radiation) was tested using a live cell tracking migration assay (IBIDI μ-slide Chemotaxis; ibidi GmbH, Martinsried, Germany) used according to the manufacturer's directions. After seeding of 0.3 × 106 MSC/mL in collagen, MSCs were exposed to a gradient between supernatants of non-irradiated and irradiated HuH7 or MDA-MB-231 cells. Monitoring of chemotaxis and tracking of 20 randomly selected cells was done as previously described. 31 For quantification of the migratory behavior of MSCs, the forward migration index (FMI), the center-of-mass (CoM), and directness were calculated, reflecting the distance and directness of migration for all tracked cells.
Animals
Five-week-old female CD1 nu/nu mice from Charles River (Sulzfeld, Germany) were housed under specific pathogen-free conditions with access to mouse chow and water ad libitum and were allowed to acclimatize for 1 week before tumor cell implantation. The experimental protocol was approved by the regional governmental commission for animals (Regierung von Oberbayern), and experiments were performed in accordance with institutional guidelines of the Klinikum rechts der Isar, Technische Universität München.
HuH7 xenograft tumors
HuH7 xenograft tumors were established by subcutaneous injection of 5 × 106 HuH7 cells in 100 μL PBS into the flank region, as described previously. 12 Tumor volumes, which were regularly measured, were estimated using the equation length × width × height × 0.52, and animals were sacrificed when tumors were >1,500 mm3.
Irradiation of HCC xenografts
Experiments were initiated when the tumors had reached a diameter of approximately 5–10 mm. The irradiation was performed using the high precision image-guided small-animal radiation research platform (SARRP; X-Strahl Ltd., Walsall, United Kingdom). Mice were anesthetized by isoflurane/oxygen inhalation for the duration of each treatment. Cone-beam computed tomography (CBCT) using 60 kV and 0.8 mA photons was performed for each mouse to visualize the subcutaneous tumors. All tumors were irradiated using 220 kV and 13 mA x-ray beams using a 3 × 9 mm2, 5 × 5 mm2, or 10 × 10 mm2 collimator. The central axis of the beams was set in the isocenter of the tumor with a mean irradiation dose of 2 or 5 Gy. The SARRP control software and Muriplan were used to target tumors and irradiation doses precisely.
MSC application and 123I-scintigraphy
Mice were given 5 mg/mL levothyroxine (LT4; Sigma–Aldrich) in their drinking water to suppress thyroidal iodide uptake. CMV-NIS-MSCs were applied once via the tail vein at a concentration of 5 × 105 cells/500 μL PBS 24 h post tumor radiation. Seventy-two hours after MSC application, mice received 18.5 MBq (0.5 mCi) 123I intraperitoneally, and radioiodide biodistribution was assessed using a gamma camera provided with a low-energy high resolution collimator
Immunohistochemical NIS staining
Immunohistochemical NIS staining of paraffin-embedded tissue sections derived from HCC xenografts or other organs (liver, lung, and spleen) after systemic CMV-NIS-MSCs administration was performed as described previously. 32
Statistical methods
Results are expressed as the mean ± standard error of the mean (SEM) or mean fold change ± SEM, and statistical significance was tested by two-tailed Student's t-test, where p-values <0.05 were considered significant. For migration assays, results are shown by Rayleigh test, where p-values <0.05 were considered significant.
Results
Irradiated HuH7 cells show increased expression of chemokines and growth factors implicated in MSC migration
Tumor irradiation is thought to increase the expression of diverse chemokines and growth factors, many of which have been implicated in directed migration of MSCs. To study this in detail, HuH7 cells were evaluated for their steady-state mRNA expression of CXCL8, CXCL12, PDGFB, FGF2, TGFB1, THBS1, and VEGF using qRT-PCR. Upon irradiation, the mRNA expression for each of these factors was increased compared to non-irradiated cells, but the time course and response to radiation levels varied among the genes tested (Fig. 1). The in vitro irradiation (1–10 Gy) of HuH7 cells did not result in a change in cell viability compared to non-irradiated cells during 0–48 h (data not shown). For radiation with 0, 2, and 5 Gy (as used in vivo), HuH7 cell viability was examined at later time points, such as 5 or 7 days, and no decrease in cell viability was seen, except after 5 days using 5 Gy where a decrease of 18% was seen (data not shown).
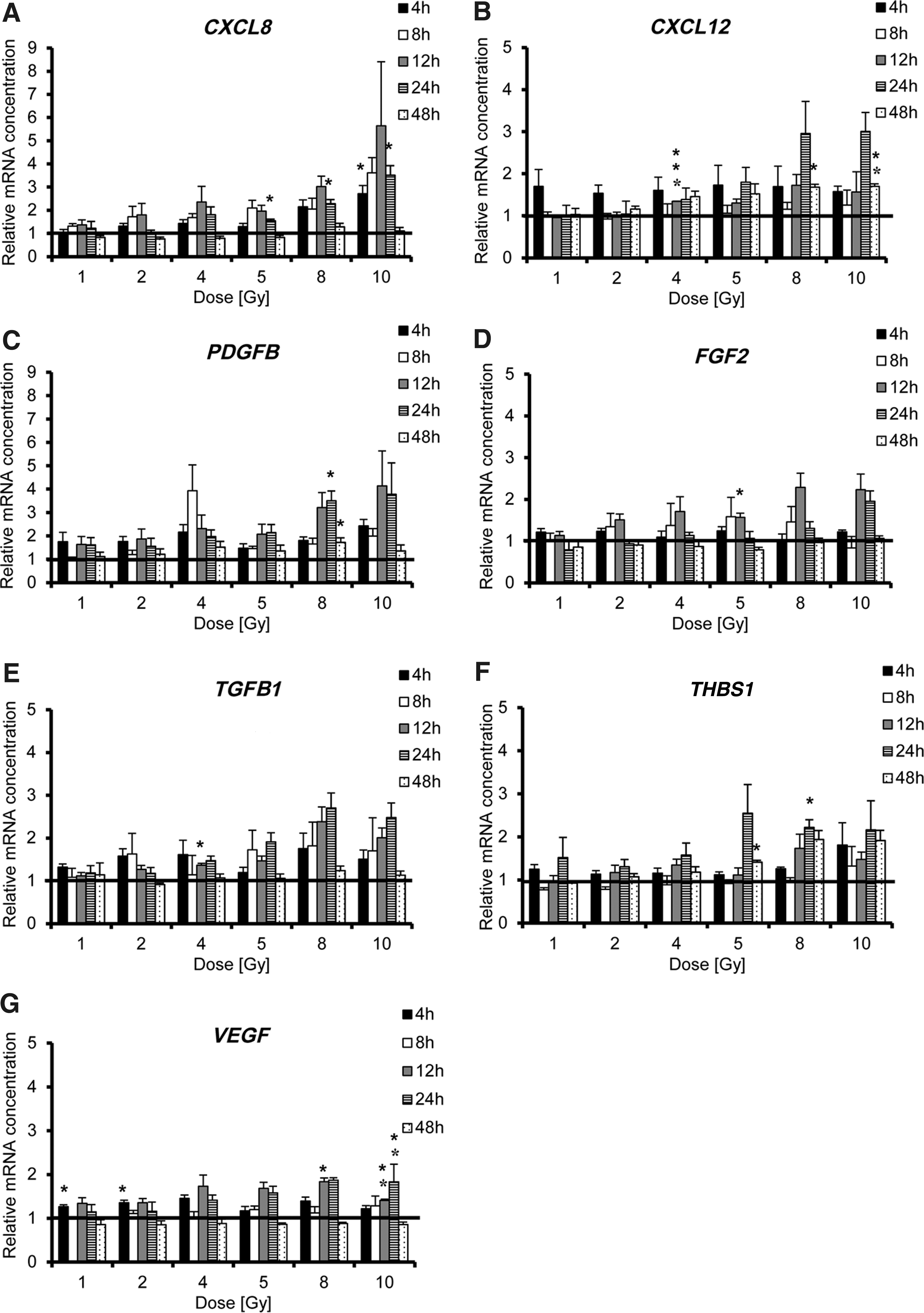
Gene expression of inflammatory cytokines and growth factors is enhanced by irradiation of HuH7 cells (1–10 Gy) during a period of 4–48 h
Changes in mRNA levels were observed as early as 4 h post radiation treatment for all factors, with some of them increasing in a radiation dose-dependent manner (CXCL8, PDGFB, TGFB1, and THBS1; Fig. 1A, C, E, and F). A peak in mRNA levels after irradiation was observed at 12 h for CXCL8 and FGF2, which was also dose dependent, whereas a dose-dependent peak in CXCL12 and THBS1 mRNA expression was seen at 24 h (Fig. 1A, B, D, and F). The maximum dose-dependent increase in PDGFB, TGFB1, and VEGF mRNA expression was seen similarly at 12 and 24 h post irradiation (Fig. 1C, E, and G). An effect of radiation on the steady-state mRNA expression of CXCL12, PDGFB, and THBS1 was observed up to 48 h (Fig. 1B, C, and F).
Effects seen on the mRNA level were then confirmed by ELISA (CXCL8, CXCL12/SDF-1, TGF-β1, TSP-1, and VEGF; Fig. 2). Protein levels for FGF2 and PDGFB remained below the detection limit of the ELISA kits. No increase in protein secretion was detected from 4 to 24 h post radiation (data not shown). CXCL8, CXCL12/SDF-1, and TGF-β1 secretion showed a strong increase 48 h after the irradiation of HuH7 cells that also occurred in a dose-dependent manner, with peak production seen at 4–5 Gy (Fig. 2A–C), whereas only a marginal increase of TSP-1 and VEGF was observed (Fig. 2D and E). Significant secretion of CXCL8 protein was observed after irradiation with 8 or 10 Gy and of CXCL12/SDF-1 when using 2 Gy (Fig. 2A and B). Irradiation of HuH7cells with 4 Gy showed a significant increase in TGF-β1 protein secretion (Fig. 2C).
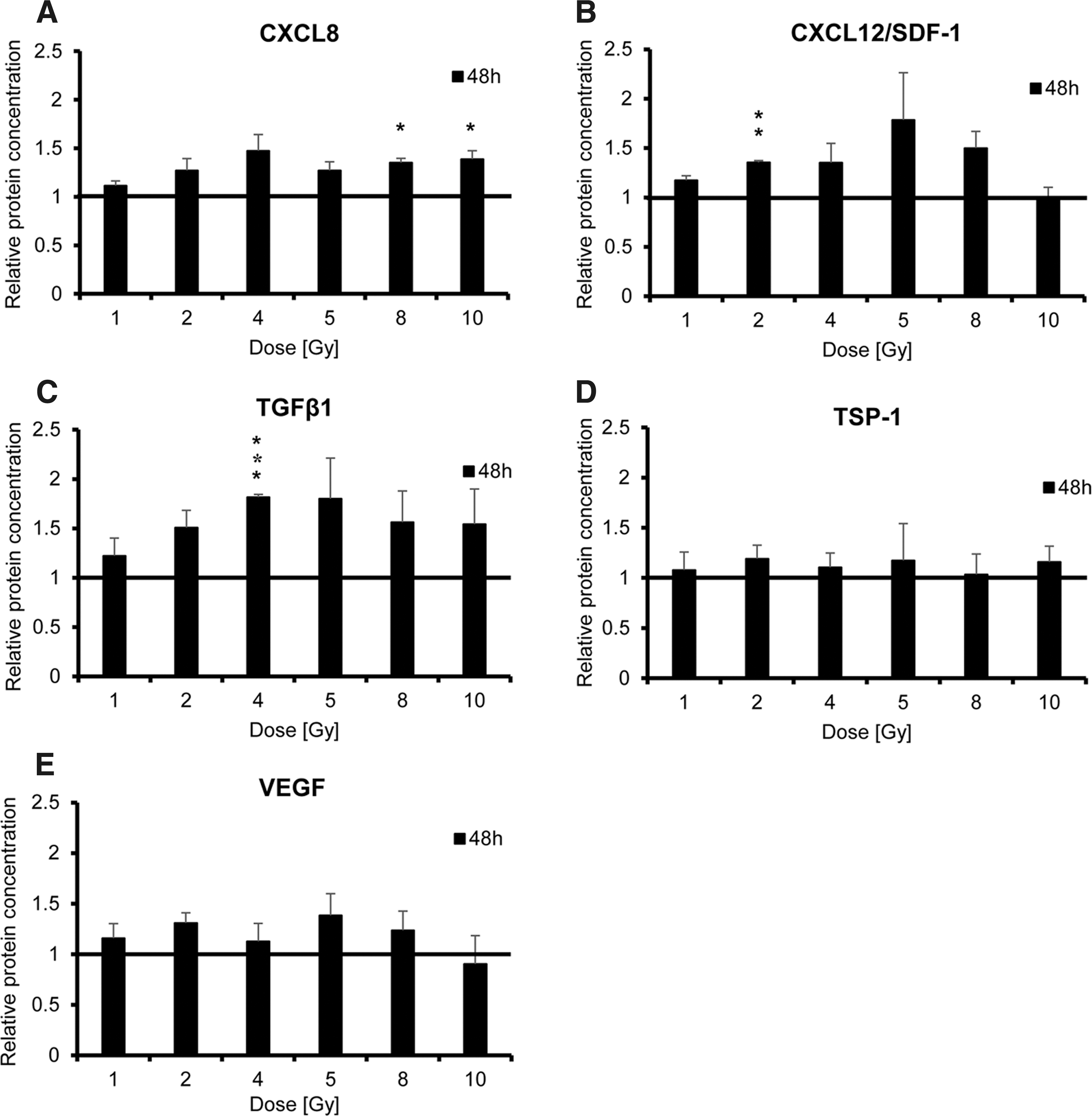
Protein secretion was assessed by enzyme-linked immunosorbent assay and showed enhanced secretion of CXCL8, CXCL12/SDF-1, TGF-β1, TSP-1, and VEGF 48 h post irradiation of HuH7 cells (1–10 Gy)
MSCs exposed to irradiated HuH7 supernatant show enhanced directed migration
Based on the results detailed above, it is clear that a complex set of factors linked to MSC recruitment are induced in the tumor line in response to external beam radiation. To validate the effect of HuH7 irradiation functionally on the directed migration of MSCs, a commercial 3D migration chamber and pseudo video imaging was applied. MSCs were subjected to a gradient generated between supernatants derived from non-irradiated and irradiated HuH7 cells, and the migratory behavior of individual cells was monitored over a 24 h period (Fig. 3). MSCs showed enhanced and directed chemotaxis toward the irradiated supernatants. The analysis revealed a robust increase of mean FMI (yFMI), mean CoM (yCoM), and mean directionality of MSCs toward supernatants from irradiated as compared to non-irradiated tumor cells (Fig. 3I–K). Enhanced MSC migration toward the supernatant isolated from irradiated cells was seen at 4 Gy (yFMI: 0.115 ± 0.015; Rayleigh values p < 0.05; yCoM: 94.42 ± 3.19 μm; Fig. 3E and I–J), 5 Gy (yFMI: 0.105 ± 0.026; Rayleigh values p < 0.05; yCoM: 73.67 ± 12.69 μm; Fig. 3F and I–J), and 10 Gy (yFMI: 0.105 ± 0.005; Rayleigh values p < 0.05; yCoM: 103.02 ± 4.57 μm; Fig. 3H and I–J) compared to controls. MSCs that were subjected to medium and supernatant from untreated HuH7 cells (yFMI: −0.01 ± 0.025; yCoM: −9.04 ± 15.07 μm; Fig. 3A and I–J) or to supernatant from untreated HuH7 cells in both chambers (yFMI: 0.055 ± 0.03; yCoM: 46.94 ± 20.64 μm; Fig. 3B and I–J) showed random chemokinesis but no directed migration. This was further confirmed by testing primary MSCs, which were subjected to untreated supernatants from HuH7 cells and from cells that were irradiated with 0, 1, 5, or 10 Gy (Fig. 4). Primary human bone marrow–derived MSCs showed similar migratory behavior as seen in the immortalized MSCs (Figs. 3 and 4).
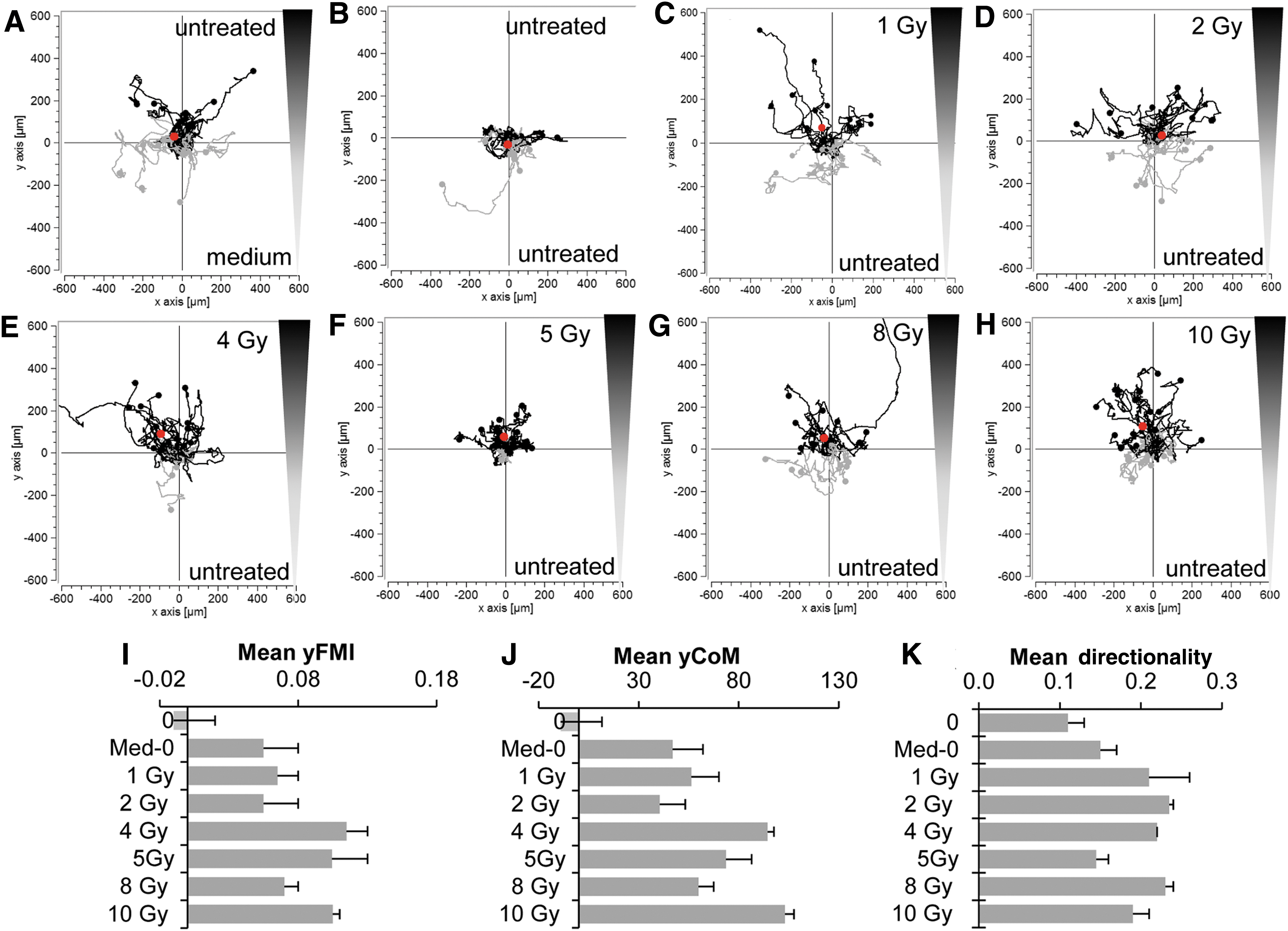
Mesenchymal stem cells (MSCs) subjected to a gradient of isolated supernatants from non-irradiated and irradiated HuH7 cells (1–10 Gy) showed enhanced direct migration over 24 h
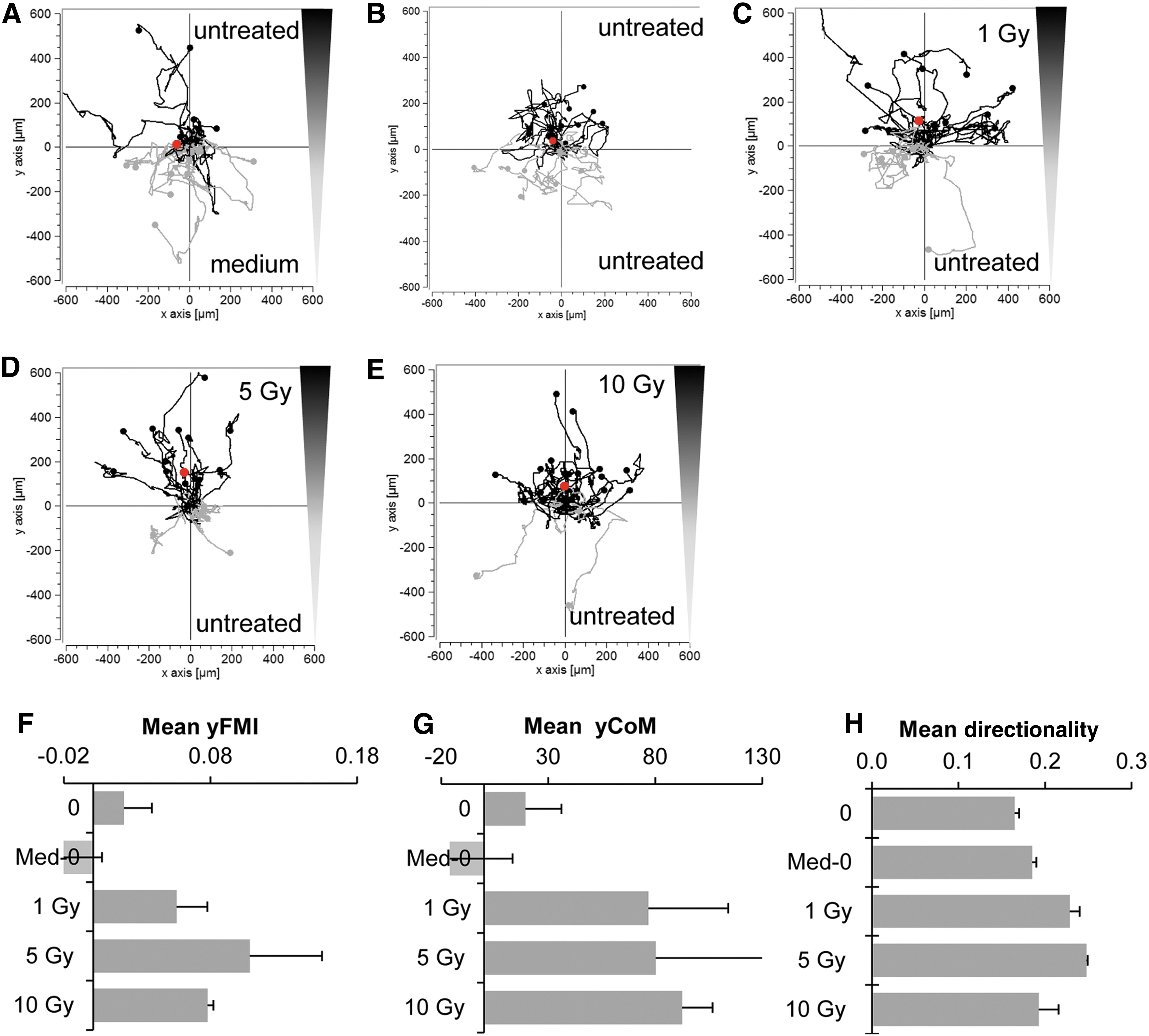
Chemotactic behavior of MSCs was further confirmed using primary human bone marrow–derived MSCs
Irradiation of MDA-MB-231 cells leads to enhanced chemokine and growth factor expression, as well as directed MSC migration to cell supernatants
To demonstrate that the effects of irradiation on human tumor cells is not only seen in HuH7 cells but also in other cancer cell lines, the human breast adenocarcinoma cell line MDA-MB-231 was irradiated in vitro using 0, 2, 5, or 10 Gy, and mRNA expression was tested 0, 8, 24, and 48 h post radiation (Supplementary Fig. S1; Supplementary Data are available online at
To validate the effect of MDA-MB-231 irradiation functionally on the directed migration of MSCs, MSCs were subjected to a gradient generated between supernatants derived from non-irradiated and irradiated MDA-MB-231 cells, as described above (Supplementary Fig. S2). Similar to the effects seen above with supernatants from HuH7 cells, MSCs showed directed chemotaxis toward irradiated supernatants. An increase of yFMI, yCoM, and mean directionality of MSCs toward supernatants from irradiated compared to non-irradiated MDA-MB-231 cells was observed (Supplementary Fig. S2F–H). While there was only a minor effect observed using 2 and 10 Gy (Supplementary Fig. S2C, E, and F–H), clearly enhanced MSC migration toward the supernatant isolated from 5 Gy irradiated cells was observed (yFMI: 0.121 ± 0.036; Rayleigh values p < 0.01; yCoM: 66.04 ± 23.93 μm; Supplementary Fig. S2D and F–H) compared to controls (Supplementary Fig. S2A, B, and F–H). Controls, in which MSCs were subjected to medium and supernatant from untreated MDA-MB-231 cells (yFMI: −0.061 ± 0.012; yCoM: 34.02 ± 0.78 μm; Supplementary Fig. S2A and F–H) or to supernatant from untreated MDA-MB-231 cells in both chambers (yFMI: −0.01 ± 0.01; yCoM: −0.28 ± 0.21 μm; Supplementary Fig. S2B and F–H), only showed random chemokinesis.
Radiation pretreatment of tumors enhances MSC recruitment in vivo
To validate radiation-enhanced MSC recruitment in an in vivo setting, subcutaneous HCC xenograft tumors implanted in nude mice were irradiated with 0, 2, or 5 Gy before systemic application of NIS-reporter engineered MSCs (Fig. 5). Using high precision image-guided SARRP, local tumor irradiation with CT-guided imaging was performed (Fig. 5C). Isocenters were set in the tumor image to locate beams and calculate dose volume distribution within the tumors (Fig. 5A and C). After one intravenous injection of CMV-NIS-MSCs 24 h post radiation, MSC-mediated delivery of NIS expression into tumors was monitored by NIS reporter gene imaging using 123I-scintigraphy (Fig. 6). Imaging studies revealed a significant tumor-specific accumulation of 7.9 ± 1.4% and 9.2 ± 1.5% of the total amount of applied 123I dose per gram tumor (ID/g) in tumors pretreated with 2 or 5 Gy, respectively, compared to 5.3 ± 0.8% ID/g in non-irradiated tumors (Fig. 6A–D). A tumor-absorbed dose of 17.4 mGy/MBq/g tumor (non-irradiated), 27.6 mGy/MBq/g tumor (2 Gy), and 64.4 mGy/MBq/g tumor (5 Gy) 131I with an effective half-life of 2.7 h (non-irradiated), 2.3 h (2 Gy), and 2.5 h (5 Gy) was determined. Endogenous NIS expression was also seen in the thyroid gland, the salivary glands (SG), as well as in the stomach and, due to renal excretion of radioiodide, in the urinary bladder (Fig. 6A–C).
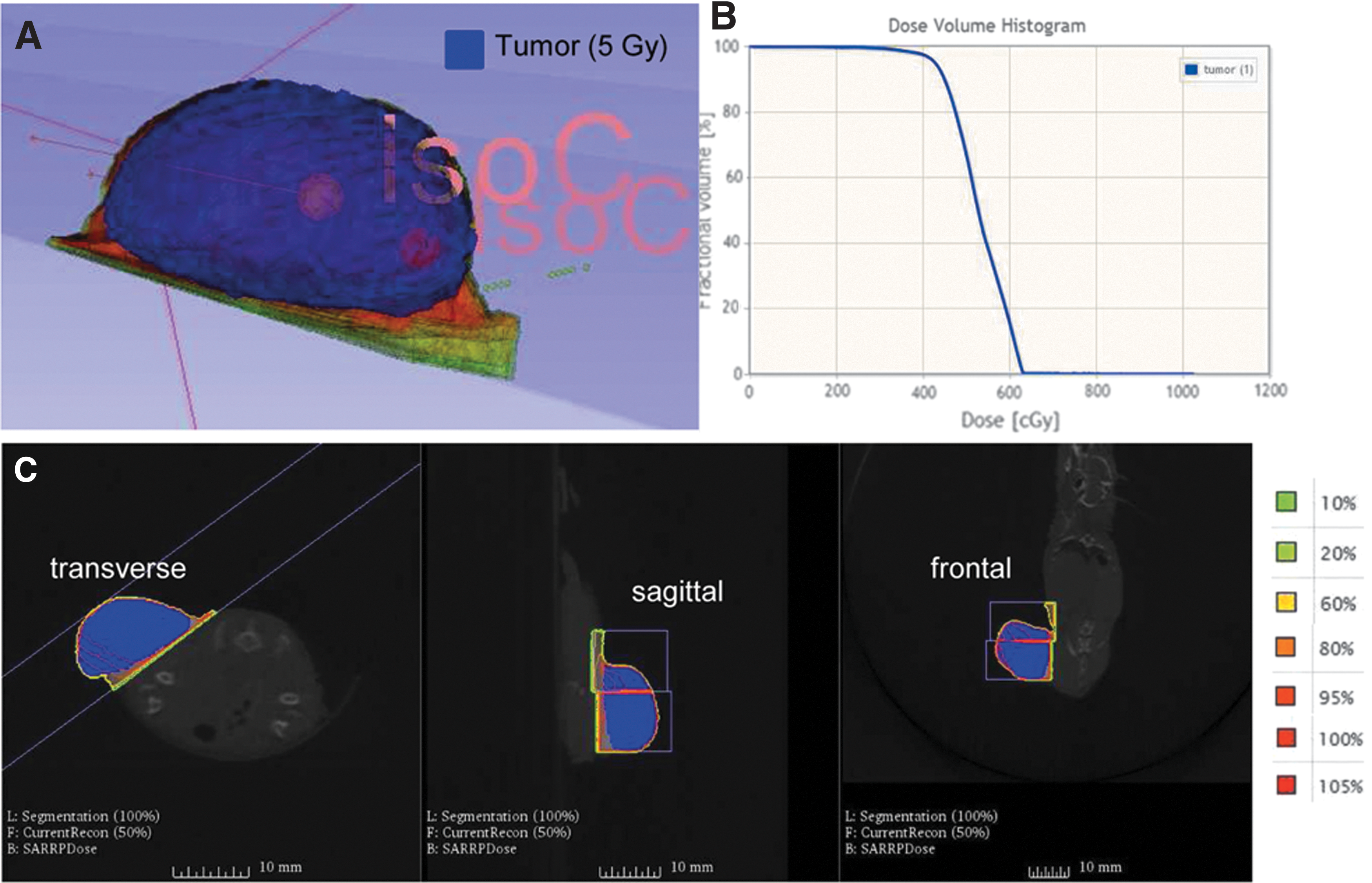
Irradiation was performed using the small-animal radiation research platform. Upon radiation with 2 or 5 Gy, a computed tomography (CT) scan was done allowing the tumor to be marked and defined (blue)

One hour after radioiodide injection, 123I-scintigraphy
After 123I-scintigraphy, tissue sections of the tumors and non-target organs (liver, lung, and spleen) were stained for NIS protein expression to determine the presence of MSCs by MSC-mediated NIS expression (Fig. 7). Non-irradiated tumors revealed only weak NIS expression in the tumors (Fig. 7A), whereas a pretreatment with 2 or 5 Gy led to higher levels of NIS expression, confirming a higher recruitment of MSCs to irradiated tumors (Fig. 7E and I). No staining was observed in non-target organs (Fig. 7B–D, F–H, and J–L).
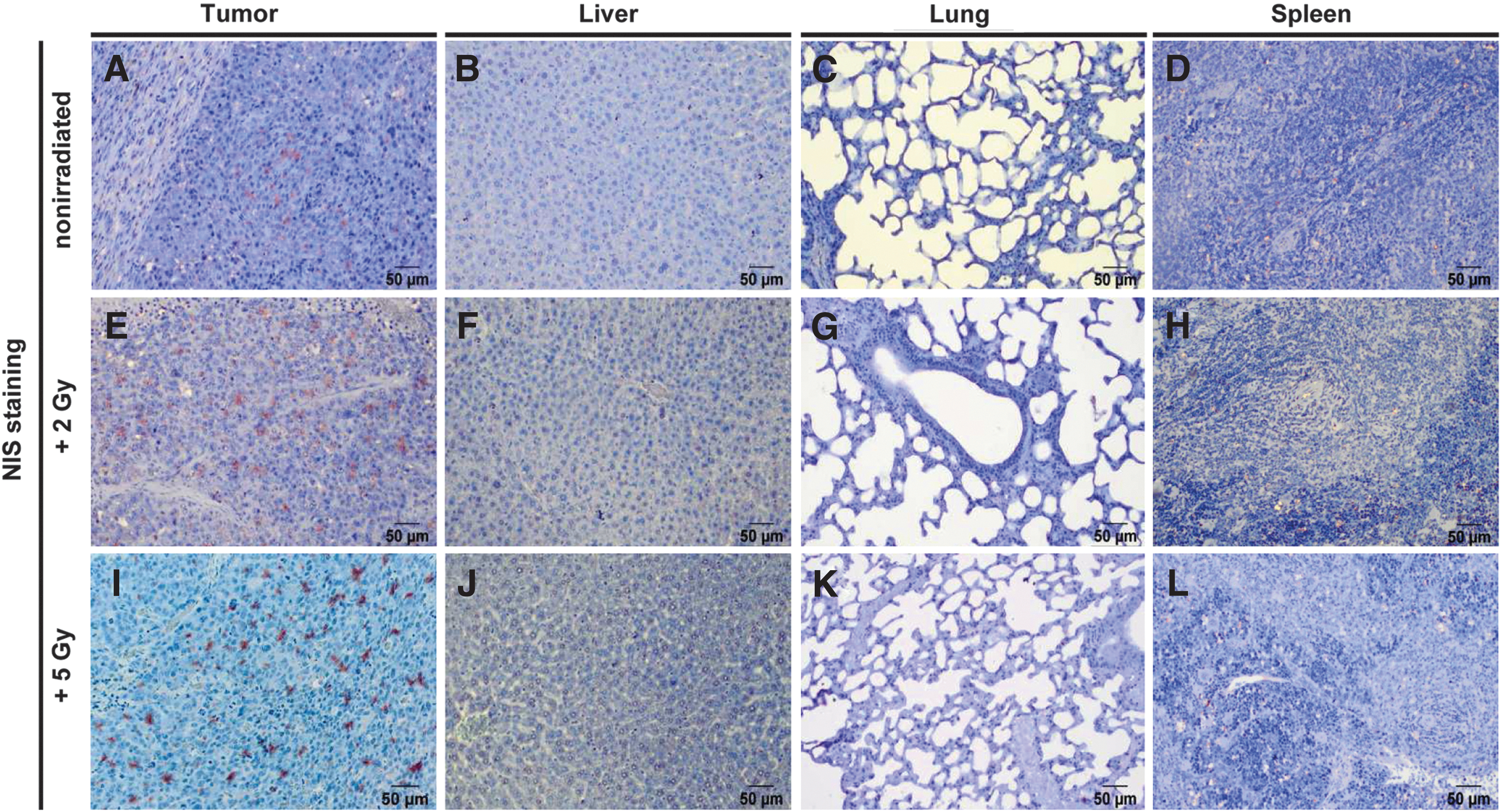
Compared to non-irradiated tumors
Discussion
MSCs are currently under evaluation as tumor delivery vehicles based in part on their excellent tumor-homing properties. An important issue in the further development of this strategy is how well it could work when used in combination with existing tumor therapy. External beam radiotherapy is the most common form of cancer radiotherapy in use today. The present study sought to evaluate the potential effects of radiation on the ability of MSCs, engineered to express the reporter gene NIS, to invade experimental tumors. The mechanisms underlying MSC recruitment to tumors have not been fully elucidated. The process is thought to mirror the mechanisms that are used by leukocytes during their recruitment to inflamed tissues, utilizing a cascade of events driven by cytokines, chemokines, integrins, and selectins. A group of factors including PDGFB, VEGF, and FGF2, chemokines/cytokines such as CXCL8 and CXCL12/SDF-1, and the pleotropic growth factor TGF-β1/3 among others have been linked to MSC migration. 33 The homing of MSCs to tumor tissue can in some instances be enhanced by external beam radiation tumor pretreatment, potentially through increased secretion of chemokines and growth factors resulting from radiation injury. 33 Previous studies by Klopp et al. have shown improved engraftment of adoptively applied MSCs to irradiated murine mammary carcinoma compared to non-irradiated tumors. 27 Similar results were also demonstrated in gliomas using hematopoietic progenitor cells, where increased migration was documented after gliomas were irradiated. 34
To test enhanced MSC homing following radiation treatment in a model of human HCC, first the question was addressed as to whether the expression and secretion of factors involved in the regulation of MSC migration were increased in the HCC cell line HuH7 following treatment. 35 –37 The response to radiation differed between the genes tested with regards to level, time course, and general response to radiation. The pleotropic cytokine TGF-β1 is expressed in HCC and has been shown to promote tumorigenesis in established HCC tumors and to have an immunosuppressive effect. 37 In this study, irradiation of HuH7 cells revealed a dose-dependent increase in TGFB1 mRNA expression, as well as enhanced protein secretion, though a slight decline in expression was observed when using high radiation doses (8 or 10 Gy). A similar effect was observed for VEGF, a central regulator of angiogenesis. 37 Irradiated HuH7 cells also showed an increase in expression of another angiogenic factor, PDGFB. At the mRNA level, the highest increase was observed 12 h post radiation treatment. However, PDGF-BB protein remained under the detection limit. Similar results were described in previous studies by Klopp et al., where a dose-dependent increase of TGF-β1, PDGF-BB, and VEGF protein expression was observed over a period of 48 h, when a murine mammary carcinoma cell line (4T1 cells) was irradiated with 0, 1, or 2 Gy. 27 In the HuH7 cell line, protein expression was not influenced by radiation before 48 h, whereas changes at the mRNA level appeared by 4 h post radiation treatment. In addition, FGF2 was investigated, which is also known as a central regulator of the migratory potential of human MSCs. 38 Analysis of FGF2 revealed an increase on the mRNA level similar to PDGFB gene expression, but it remained below the detection limit for protein expression of irradiated HuH7 cells. Further investigation of the effect of radiation on HuH7 cells showed an increase of TSP-1 and the potent pro-inflammatory chemokines CXCL8 and CXCL12/SDF-1 on the mRNA and protein level. An early effect of HuH7 cell radiation was observed at the level of mRNA, where a wide range of chemokines and growth factors were increased as early as 4 h after irradiation and remained elevated until 24–48 h post radiation treatment and for most factors an increase in protein secretion after 48 h. Thus, the data clearly demonstrate the ability of HuH7 cells to increase their expression of cytokines and growth factors directly involved in MSC recruitment after radiation treatment. These findings were further confirmed in another human cancer cell line derived from breast adenocarcinoma (MDA-MB-231). MDA-MB-231 showed a similar response to radiation as HuH7 by increasing mRNA expression levels of CXCL8, TGFB1, THBS1, and VEGF in a dose-dependent manner using 0, 2, 5, or 10 Gy, but differed in regard to mRNA levels and time course (0, 8, 24, or 48 h).
An in vitro migration assay was then performed to corroborate that irradiation of tumor cells can cause an increased migration of MSCs. Those experiments confirmed enhanced direct chemotaxis of MSCs to supernatants isolated from irradiated HuH7 and MDA-MB-231 cells, substantiating the hypothesis, and formed the basis for an in vivo approach to improve the NIS-mediated iodide uptake in non-thyroidal tumors using MSCs as gene delivery vehicles by pretreating tumors with radiation. Previous studies demonstrated the excellent tumor-homing of MSCs into the microenvironment of different tumors. 12 –15,31,39 –41 The theranostic gene NIS is used for noninvasive imaging of functional NIS expression by 123I-scintigraphy, allowing efficient tracking of NIS engineered MSCs in vivo by dynamic radioiodide imaging. The authors and others have extensively investigated the capacity of NIS to induce radioiodide accumulation in non-thyroidal tumors using MSCs as NIS gene delivery vehicles. 12 –15,19 The efficacy of MSCs to deliver the NIS gene deep into tumors has been shown in different approaches by the authors' group. 12 –15 Active MSC engraftment has been demonstrated in various tumor models, including subcutaneous and orthotopic HCC xenografts, as well as a hepatic colon cancer metastases model. 12 –15 These MSC-based NIS gene therapy approaches resulted in a significant delay in tumor growth and prolonged animal survival after radioiodine accumulation. The authors sought to build upon these results to improve the level and selectivity of MSC tumor homing, and to enhance the therapeutic efficacy of NIS-mediated 131I treatment to increase selectivity, efficacy, and safety further. Using a previously described subcutaneous HCC mouse model and reporter MSCs engineered with a CMV-NIS construct, significantly increased MSC recruitment was demonstrated into tumor sites that had been irradiated with low doses (2 and 5 Gy). Francois et al. demonstrated enhanced MSC migration after treatment of tumors with high doses (26.5 Gy) of radiation. 42 Studies by Zielske et al. showed that more MSCs migrated to tumors irradiated with a minimum of 5 Gy. 43 Different tumor types, including breast, colon, and head/neck tumors, were irradiated with 2, 5, or 9 Gy. Analysis of MSC recruitment showed enhanced recruitment in all tumors irradiated with 5 or 9 Gy, except in tumors established from UMSCC1 head/neck carcinoma cells. 43 On the other hand, Klopp et al. were able to attract MSCs efficiently to tumor sites of breast cancer using low doses of irradiation such as 1 and 2 Gy. 27
The present study examined if MSC recruitment is enhanced in tumors pretreated with low-dose irradiation (2 or 5 Gy) using NIS imaging (123I-scintigraphy). Tumor-specific radioiodide uptake activity was observed in all groups (0, 2, and 5 Gy). Tumors locally treated with radiation revealed a significantly higher NIS-mediated iodide uptake after a single MSC application 24 h post radiation treatment compared to non-irradiated tumors. The maximum radioiodide uptake activity of tumors irradiated with 2 or 5 Gy was 7.9% and 9.2% ID/g, respectively, whereas non-irradiated tumors accumulated only 5.3% ID/g. The enhanced recruitment of MSCs into irradiated HuH7 tumors was also verified ex vivo using NIS immunohistochemistry that showed increased recruitment of NIS-MSCs in tumors pretreated with low-dose radiation.
Importantly, the positive effects seen with low-dose irradiation (with lower toxicity and side effects on the normal surrounding tissues) effectively sets the stage for more detailed studies of the combined effects of enhanced delivery of therapy genes by MSCs with cancer radiation treatment. Radiotherapy has also been shown to have effects on the tumor stroma, which is comprised of tumor endothelial cells and cancer-associated fibroblasts (CAFs) as well as other cell types. 44 Radiation of tumors not only leads to enhanced stimulation of cytokine and growth factor secretion by tumor cells, but is also responsible for radiation-induced dysfunction or even apoptosis of tumor endothelial cells, which in turn lead to hypoxia, increased permeability, as well as an enhanced inflammatory response. 44,45 Due to vascular depletion, the pro-angiogenic and pro-vasculogenic factors VEGF and CXCL12/SDF-1 are increased. 44,46,47 Radiation also drives activation of CAFs within the tumor stroma, which in turn increase growth factor secretion and the release of cytokines such as HGF, PDGF, CXCL12/SDF-1, and TGF-β. 44 Another aspect besides an increased inflammatory response, which was shown in this study to increase MSC recruitment in vitro, is a possible radiation-induced stimulation of tumor vascular permeability, which could be another important trigger for enhanced MSC recruitment in vivo. Using doses of 5–10 Gy, an initial increase of tumor blood flow after irradiation has been reported, with a return to levels seen before irradiation 2–3 days post radiation. 48 Taking this into consideration, irradiation-induced enhanced MSC migration to tumors might be a combinatorial effect of stimulated secretion of cytokines and growth factors involved in the process of MSC tumor homing, as well as increased tumor permeability, allowing MSCs to passage the tumor stroma more easily. These complex effects of irradiation on tumor cells and their (micro-) environment not only enhances MSC tumor recruitment, as demonstrated in this study, but may also foster MSC tumor engraftment and therefore represents a highly promising tool to stimulate MSC-driven gene therapy approaches significantly. Taken together, the results show a dose-dependent increase of NIS-mediated accumulation of 123I due to enhanced MSC recruitment to irradiated tumors using low-dose irradiation. Based on these findings, an enhanced accumulation of the therapeutically active radioisotope 131I is expected after combining external beam radiotherapy and MSC gene delivery, which will be evaluated in future studies.
In summary, this study demonstrates that radiation of experimental HCC enhances the secretion of diverse inflammatory cytokines and growth factors involved in MSC migration, which can be linked to enhanced migration of adoptively applied MSCs deep into tumor microenvironments. These data open up the exciting prospect for a new therapy approach, in which HCC tumors are pretreated with low-dose radiation prior to systemic MSC application, aiming to increase the efficacy of MSC-mediated NIS gene therapy.
Footnotes
Acknowledgments
We are grateful to Dr. S.M. Jhiang, Ohio State University, Columbus, Ohio, for supplying the full-length human NIS cDNA. We also thank Prof. Dr. K. Scheidhauer and Jakob Allmann, Department of Nuclear Medicine, Klinikum rechts der Isar der Technischen Universität München, Munich, Germany, for their assistance with the imaging studies. We thank Doris Mayr (Department of Pathology, Ludwig-Maximilians-University, Munich, Germany) for preparation of paraffin-embedded slides. This work was supported by grants from the Deutsche Forschungsgemeinschaft within the Collaborative Research Center SFB 824 to C.Sp. (project C8) and within the Priority Program SPP1629 to C.Sp. and P.J.N. (SP 581/6-1, SP 581/6-2, NE 648/5-2), as well as within a grant from the Wilhelm-Sander-Stiftung to C.Sp. (2014.129.1).
Author Disclosure
The authors declare no conflict of interest. This work was performed as partial fulfillment of the doctoral thesis of C.Sc. within the LMU Medical Faculty.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.