
Abstract
Muscle-invasive bladder cancer represents approximately 25% of diagnosed bladder cancer cases and carries a significant risk of death. Oncolytic viruses are novel antitumor agents with the ability to selectively replicate and lyse tumor cells while sparing healthy tissue. We explored the efficiency of the oncolytic YB-1-selective adenovirus XVir-N-31 in vitro and in an orthotopic mouse model for bladder cancer by intramural injection under ultrasound guidance. We demonstrated that XVir-N-31 replicated in bladder cancer cells and induced a stronger immunogenic cell death than wild-type adenovirus by facilitating enhanced release of HMGB1 and exosomal Hsp70. The intratumoral delivery of XVir-N-31 by ultrasound guidance delayed tumor growth in an immunodeficient model, demonstrating the feasibility of this approach to deliver oncolytic viruses directly into the tumor.
Introduction
B
Oncolytic viruses (OVs) selectively replicate in tumor cells and destroy them. Although the host's immune system limits the efficacy of virotherapy by rapid clearance of infection, it is becoming increasingly clearer that the induction of innate and adaptive immune responses by OVs against the tumor contributes significantly to the therapeutic efficacy of virotherapy. 6,7 In particular, the induction of immunogenic cancer cell death (ICD) by OVs has been previously suggested to be beneficial for a positive clinical response. 8,9 ICD is characterized by the release of damage-associated molecular patterns (DAMPs), such as HMGB1, ATP, calreticulin, and heat-shock proteins. 10 These molecules are released from dying tumor cells and induce signals that increase effective antigen presentation and T-cell activation and thus play an important role in eradication of cancer cells in vivo. 9 OVs have been suggested to be ideally suited for the treatment of cancer in combination with immune-checkpoint inhibitors (ICIs). 6,11,12 Early clinical phase trials have indicated that the combination has a greater antitumor activity without additional safety concerns, suggesting that oncolytic virotherapy may improve the efficacy by shutting down the immunosuppressive effects of the tumor environment. 12,13 ICIs have also recently been approved for metastatic and advanced bladder cancer. 14,15 The response rates have been limited to 20%–30%, indicating the need for additional treatment strategies such as virotherapy to overcome the immunosuppressive mechanisms. These mechanisms may act in concert to counteract effective immune responses and thereby increase the number of responding patients with MIBC. 12,14
We have developed the replication-selective adenovirus XVir-N-31. Tumor selectivity is achieved by conditional replication of XVir-N-31 in cancer cells displaying nuclear localization of the human oncogenic cellular transcription factor YB-1. 16 YB-1 is a multifunctional oncoprotein that is highly overexpressed in a variety of tumor types. 17,18 In bladder cancer, high YB-1 expression correlates with invasiveness, tumor grade, and metastasis and is associated with decreased overall survival. Consequently, YB-1 appears to be a promising target for oncolytic virotherapy in bladder cancer. 19,20 Adenovirus infection of tumor cells induces the translocalization of YB-1 into the nucleus, where it is needed for adenoviral replication. 21 The YB-1 dependency of XVir-N-31 was established by deleting the 13S splice variant of the adenoviral regulatory protein E1A. XVir-N-31 has already been shown to have strong oncolytic activity in vitro and in vivo in other tumor types, and it is unable to replicate in nontumorigenic cells. 16,22 Intravesical instillation of oncolytic viruses in several animal models for the treatment of NMIBC has been described previously. 23,24 In the present study, we analyzed the antitumor activity of the oncolytic YB-1–based adenovirus XVir-N-31 against bladder cancer in vitro. In addition to viral replication and killing capacity, we also evaluated the occurrence of ICD. In order to investigate the efficacy of OVs in an orthotopic MIBC model, we used a new method in which tumor cells are directly injected into the lamina propria under ultrasound guidance. 25 After tumor formation, direct injection of virus was carried out using ultrasound guidance as well.
Methods
Cell culture
The human bladder cancer cell lines J82, T24, and UMUC3 cells were obtained from ATCC (Manassas, VA). The BFTC905 cell line was a kind gift from Dr. Wolfgang A. Schulz, Department of Urology, Düsseldorf, Germany. Cells were cultured in RPMI1640 supplemented with 10% FCS, 1% nonessential amino acids, and 1% penicillin/streptomycin or DME-medium supplemented with 10% FCS and 1% penicillin/streptomycin (Biochrom, Berlin, Germany) at 5% or 10% CO2. To analyze the release of proteins into the cell culture supernatant, cells were slowly adapted to be maintained with 2% FCS and were cultured in protein-free medium during the experiment.
Adenoviral vectors
AdWT is the wild type of human serotype 5 adenovirus. Dl703 contains a deletion in E1 which makes it replication defective at low to medium doses. 26 XVir-N-31 is an oncolytic adenovirus and has previously been described as Ad-Delo3-RGD. 16 It has a deletion in the CR3-region of E1A, which makes its replication YB-1 dependent. 27 Furthermore, it contains a deletion in the E1B19k gene as well as in the E3 region. The fiber knob contains an RGD-motif.
Immunoblot
Cells were lysed using RIPA buffer (150 mM NaCl; 50 mM Tris/HCl; pH 7.2; 1% Triton X-100; 0.05% SDS; 5 mM EDTA; protease inhibitor cocktail; Roche Diagnostics, Penzberg, Germany). Lysates were centrifuged for 30 min at 30,000 g and protein concentrations were quantified using Pierce BCA Protein Assay (Thermo Scientific, Planegg, Germany). Proteins were loaded on 10%–12% polyacrylamide gels for SDS-PAGE and blotted on PVDF-membranes (GE-Healthcare, Chicago, IL). The following antibodies were used for immunoblotting: Hsp70 (Enzo Life Sciences, Lörrach, Germany), Calnexin, GAPDH (Cell Signaling, Danvers, MA), CD63 (Santa Cruz Biotechnology, Inc., Santa Cruz, CA), and a self-made antibody against the N-terminal region of YB-1 (Eurogentec, Lüttich, Belgium). Secondary antibodies were horseradish peroxide conjugated (Dianova, Hamburg, Germany). Immunoreactive proteins were visualized using Amersham ECL-reaction (GE-Healthcare, Chicago, IL).
ELISA
Hsp70-ELISA (Enzo Life Sciences, Lörrach, Germany) and HMGB1-ELISA (IBL International, Hamburg, Germany) were performed with cell-free cell culture supernatant according to the manufacturers' protocols and recommendations.
Quantitative real-time PCR
To investigate viral replication in bladder cancer cells, 1 × 105 cells were seeded on six-well plates and infected with 10 MOI 24 h later. Cells were harvested after 4, 24, 48, and 72 h, and DNA was isolated using DNeasy-Blood & Tissue Kit (Qiagen). For detection of viral copies following specific primers were used: Hexon-fw, GGCCATTACCTTTGACTCTTC; Hexon-rev, GCATTTGTACCAGGAACCAGTC or fiber-fw, AAGCTAGCCCTGCAAACATCA; fiber-rev, CCCAAGCTACCAGTGGCAG TA; beta-actin-fw, TAAGTAGGTGCACAGTAGGTCT GA; beta-actin-rev, AAAGTCCAAXAACACGGCTAAG. Quantitative PCR was performed on 96-well plates with SYBR Green Mastermix (Eurogentec, Lüttich, Belgium) and the following cycling conditions: 95°C for 2 min and 45 cycles 94°C for 15 s, 60°C for 15 s, and 72°C for 15 s. Gene expression was calculated using the ΔΔCT-Method. 28
Quantification of infectious viral particle formation
J82, UMUC3, and T24 cells (1 × 105) were seeded on six-well plates. Twenty-four hours after seeding, cells were infected with 10 MOI of indicated viruses in 400 μL of serum-free media. After 72 h, cells and supernatant were harvested and viruses were released from cells by three thaw-freeze cycles. Finally, samples were centrifuged for 5 min at 1,000 g. To quantify viral particles in the supernatant of cell lysates, low passage HEK293 cells were seeded on a 24-well plate at a concentration of 2 × 105 cells in 500 μL per well. Cell-free supernatant of infected bladder cancer cells was added to HEK293 undiluted and in dilutions ranging from 10−1 to 10−5 in a volume of 10 μL and 50 μL. After 40–48 h, HEK293 cells were fixed and stained according to the AdEasy Viral Titer Kit instruction manual (Agilent Technologies, Santa Clara, CA). Immuno-stainings were performed using a goat-anti-hexon antibody (Chemicon) and a rabbit-anti-goat antibody (P0449, Dako A/S, Glostrop, Denmark). Visualization was carried out by DAB solution (Dako A/S, Glostrup, Denmark).
FACS
Cells were collected with cell scratchers and pelleted by centrifugation at 500 g for 5 min. Primary CAR-antibody (Clone RmcB; Merck Millipore, Burlington, MA) was diluted 1:40 in FACS-Buffer (1% BSA in PBS). Secondary anti-mouse PE-conjugated antibody (Dianova, Hamburg, Germany) was diluted 1:50. Stained cells were fixed in 1%–2% paraformaldehyde for 20 min at 4°C. Stained cells were analyzed using BD FACS Calibur Flow Cytometry System (BD Bioscience, Heidelberg, Germany) by counting 10,000 events and FlowJo-software (FlowJo LLC).
Immunofluorescence
UMUC3, T24, and J82 cells (7 × 103) were grown directly on coverslips and infected with 30, 100, and 40 MOIs of XVir-N-31, respectively. Forty hours after infection, cells were fixed using ice-cold methanol:acetone for 15 min at −20°C. Fixed cells were blocked and stained for YB-1 (Eurogentec, Lüttich, Belgium, self-made, rabbit) and for E1A (Santa Cruz, M73, mouse) followed by incubation with secondary anti-rabbit Alexa Fluor 488-conjugated antibody (Cell Signaling, Danvers, MA) and secondary anti-mouse Cy3-conjugated antibody (Life Technologies, Carlsbad, CA). Slides were mounted with ProLong Gold Antifade Mountant with DAPI (Life Technologies, Carlsbad, CA), and pictures were taken with Microscope AxioVert.A1 and Camera AxioCam ERc5s (Zeiss, Oberkochen, Germany).
Cytotoxicity assay
Sulforhodamine B (SRB) assay was performed as previously described. 16 Cells at 5 × 104 per well were seeded on 12-well plates and infected with various MOI of the indicated virus 24 h after seeding. After 4 days, cells were fixed with 10% cold trichloroacetic acid at 4°C overnight and stained with 0.5% SRB in 1% acetic acid for 30 min. To quantify the cytopathic effect, the bound SRB was dissolved with 10 mM Tris buffer (pH 10). Absorbance was measured at 590 nm.
Isolation and analysis of exosomes
Tumor cells were infected with 20 MOI of the indicated virus and grown in serum-free medium. Exosomes were isolated 72 h after infection of bladder cancer cells from cell culture supernatant using ExoQuick-TC Exosome precipitation solution (System Biosciences) according to the manufacturer's protocol and recommendations. Exosomes were suspended in RIPA buffer and analyzed by Western blotting. Exosomes were also isolated from cell culture supernatant using an established protocol with multiple centrifugation steps as described by Thery et al. 29 In brief, cells and cell debris were removed by successive centrifugation at 300, 2,000, and 10,000 g. Exosomes were isolated by ultracentrifugation at 100,000 g for 70 min, and the exosome containing pellet was washed with PBS and again pelleted by ultracentrifugation at 100,000 g for 70 min. The pellet was suspended in 100 μL of PBS and analyzed by dynamic light scattering (DLS), transmission electron microscopy (TEM), and Western blotting.
For DLS, 10 μL of solution containing exosomes was added to 10 mM NaCl (H2O for latex beads) to a final volume of 800 μL. Measurements were performed using Malvern Nano ZS zetasizer (Malvern Instruments, Malvern, United Kingdom) as recently described. 30
Ultrasound-guided injection of UMUC3 and monitoring of tumor growth
Seven-week-old athymic nude mice were purchased from Charles River. All animal experiments were performed in accordance with German law. The protocol was approved by the District Government of upper Bavaria. Mice were anesthetized with 100 μL of MMF (medetomidine, 0.5 mg/kg; midazolam, 5 mg/kg; fentanyl, 0.05 mg/kg). Ultrasound-guided intramural tumor cell injection was performed according to Jäger et al. 25 Therefore, 2 × 105 UMUC3 cells suspended in 50 μL of Matrigel (Sigma Aldrich, St. Louis, MO, E1270) was used. Briefly, the bladder was visualized by ultrasound. An artificial space was created in the bladder wall using a 30-G needle filled with PBS. Subsequently, tumor cells were injected into this space. Tumor growth was monitored by three-dimensional ultrasound twice a week. Mice were anesthetized using isoflurane, and the bladder was scanned as a whole in 0.08-mm steps. The tumor volume was calculated using the Visual Sonics imaging software package by analysis of every fifth picture according to the user manual.
Treatment
According to their tumor volume, mice were randomized into two groups. Treatment started at a mean tumor volume of 25 mm3 per group. Mice were treated three times by ultrasound-controlled intratumoral injections of PBS or XVir-N-31. A 30-G needle was filled with 10 μL of virus (2 × 109 viral particles) or PBS and stored on ice until use. The needle was inserted in the center of the tumor, and the correct position was verified by ultrasound. 25 Injection of virus or PBS was performed slowly and under controlled retraction of the needle to ensure a good distribution of the virus within the tumor. At 17–29 days after tumor cell injections mice were euthanized, and the bladder was used for further immunohistochemical analysis.
Histology and immunohistochemistry
Animals were sacrificed around 20 days after injection of tumor cells for animal welfare reasons. The bladder was removed from each animal, fixed in 3.5% paraformaldehyde, embedded in paraffin, and cut in 3.5-μm sections. Sections were used for hematoxylin and eosin (H&E) staining. For the immunohistochemistry staining, antigen retrieval was performed in a pressure cooker using citrate buffer (pH 6). Endogenous peroxidase activity was blocked by incubation in 6% H2O2 for 6 min. Slides were washed with TBS prior to blocking with 5% FBS-TBS (for Ki-67, F4/80) or 1% BSA-PBS (for Hexon) for 2 h at room temperature. Sections were incubated in a humidifying chamber with primary antibodies Ki-67 (MIB-1, Dako, 1:100), Hexon (AB1056, Chemicon, 1:500), and F4/80 (D2S9R, Cell Signaling Technology, 1:250) for 2 h at room temperature. To visualize the Ki-67 and F4/80 staining, the Dako REAL EnVision Detection system (K5007) was used according to the manufacturer's manual. To visualize the Hexon staining, sections were incubated with a horseradish peroxidase–conjugated rabbit-anti-goat antibody for 30 min prior to adding the Dako DAB+Substrate System (K3468). Staining was monitored under the microscope and stopped with TBS. Counterstaining was performed by using hematoxylin.
Statistics
The statistical analysis was performed with Microsoft Excel software. Statistical significance was determined with the two-tailed Student's t-test, with p ≤ 0.05 considered statistically significant.
Results
YB-1 expression and localization in various bladder cancer cell lines
The multifunctional protein YB-1 has been described as promoting tumor growth, invasion, and drug resistance in bladder cancer cells. 19 To investigate XVir-N-31–based virotherapy in bladder cancer cell lines, we analyzed YB-1 expression on the protein level by using immunoblotting in UMUC3, T24, J82, and BFTC905 cells (Fig. 1A). All four cell lines showed strong expression of YB-1. Since the subcellular localization of YB-1 plays a crucial role for the replication efficiency of XVir-N-31, we used immunofluorescence staining for YB-1 and the adenoviral protein E1A in infected and uninfected cells (Fig. 1C). In uninfected cells, the majority of YB-1 was perinuclear in the cytoplasm, with a weak nuclear appearance of YB-1. In contrast, infection of cells with XVir-N-31 (positive for E1A) led to a distinct localization of YB-1 in the nucleus.
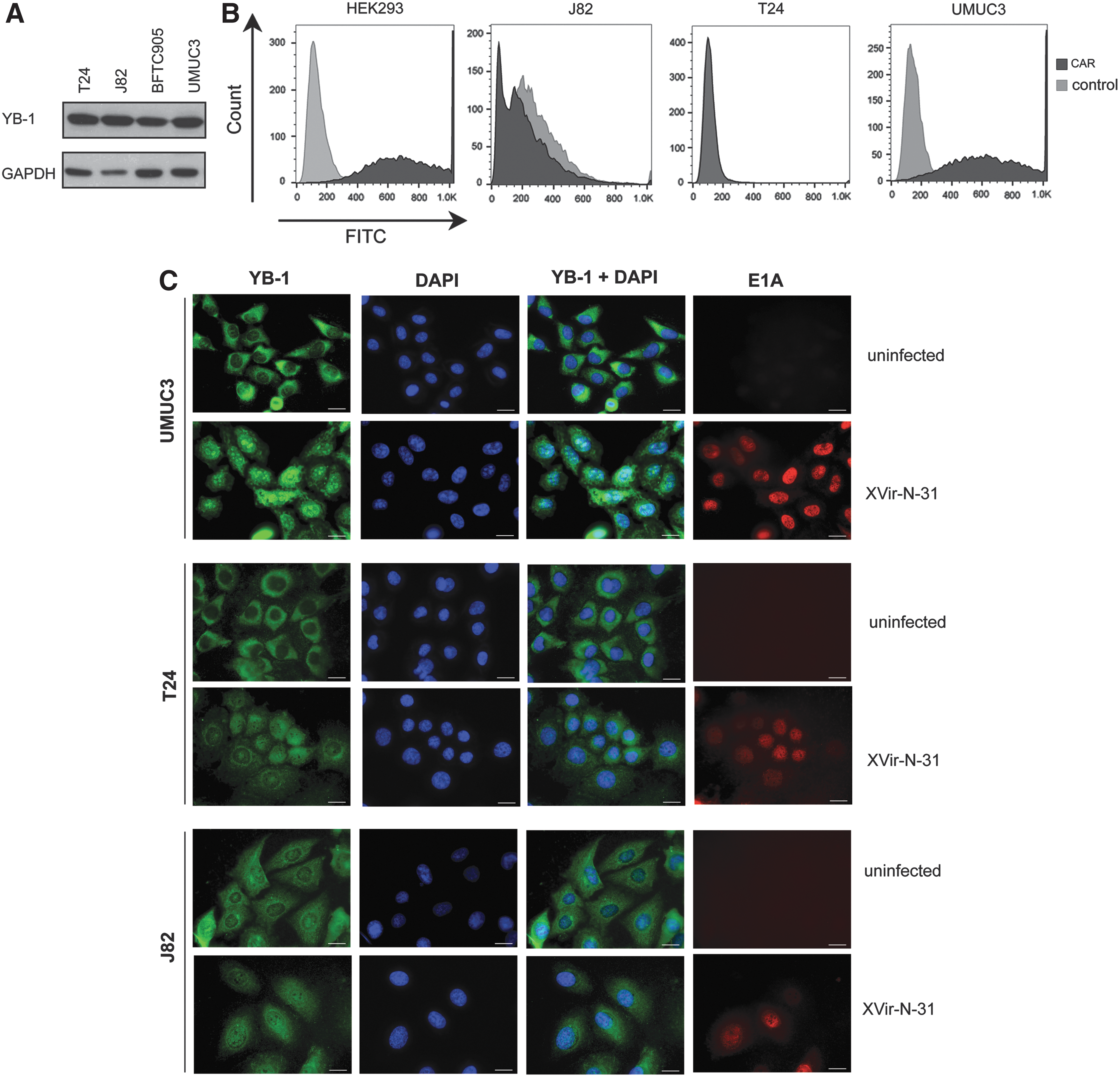
YB-1 and Coxsackie-adenoviral receptor (CAR) expression in bladder cancer cell lines.
Coxsackie-adenoviral receptor expression in various bladder cancer cell lines
Adenoviruses are known to bind to the Coxsackie-adenoviral receptor (CAR) and integrins to infect cells. 31 Although XVir-N-31 contains the integrin binding motif RGD, which increases CAR-independent infection, 32 virus infectivity still plays a pivotal role in its cell-killing capacity. Therefore, it appeared necessary to examine CAR expression in bladder cancer cells. FACS analysis showed clear differences in membrane-bound CAR levels in different bladder cell lines (Fig. 1B). While UMUC3 cells displayed high levels of CAR expression (like HEK293 cells), J82 and T24 cells showed low CAR expression. 33
Cell-killing capacity of XVir-N-31 in T24, UMUC3, and J82 cells
To quantify the cell-killing capacity, we performed an SRB assay on UMUC3, T24, and J82 cells. Cell killing of XVir-N-31 was analyzed 4 days after infection with indicated MOI and compared to wild-type adenovirus (AdWT) and the E1-deleted, replication-defective adenoviral vector dl703. The dl703 served as a negative control and had little effect on cell viability at MOIs higher than 100, showing that active replication is decisive in inducing tumor cell killing. As shown in Fig. 2, XVir-N-31 had an MOI-dependent cell-killing effect that varied slightly between J82, UMUC3, and T24 cells. XVir-N-31 caused cell death in J82, T24, and UMUC3 cells at concentrations of 5, 25, and 10 MOI, respectively. AdWT induced cell death at 10 MOI in J82, 50 MOI in T24, and 1 MOI in UMUC3 cells. The low cell-killing effect of AdWT in T24 cells is in accordance with the low CAR expression of these cells. Since the RGD-armed XVir-N-31 also killed the CAR negative cell line T24 efficiently, bladder cancer cells are suitable for targeting by XVir-N-31. Furthermore, XVir-N-31 showed a higher oncolytic effect in the CAR-negative cell lines J82 and T24, compared to AdWT, which we ascribe to a better infection with the RGD motif and the deletion of the anti-apoptotic protein E1B19k in XVir-N-31.
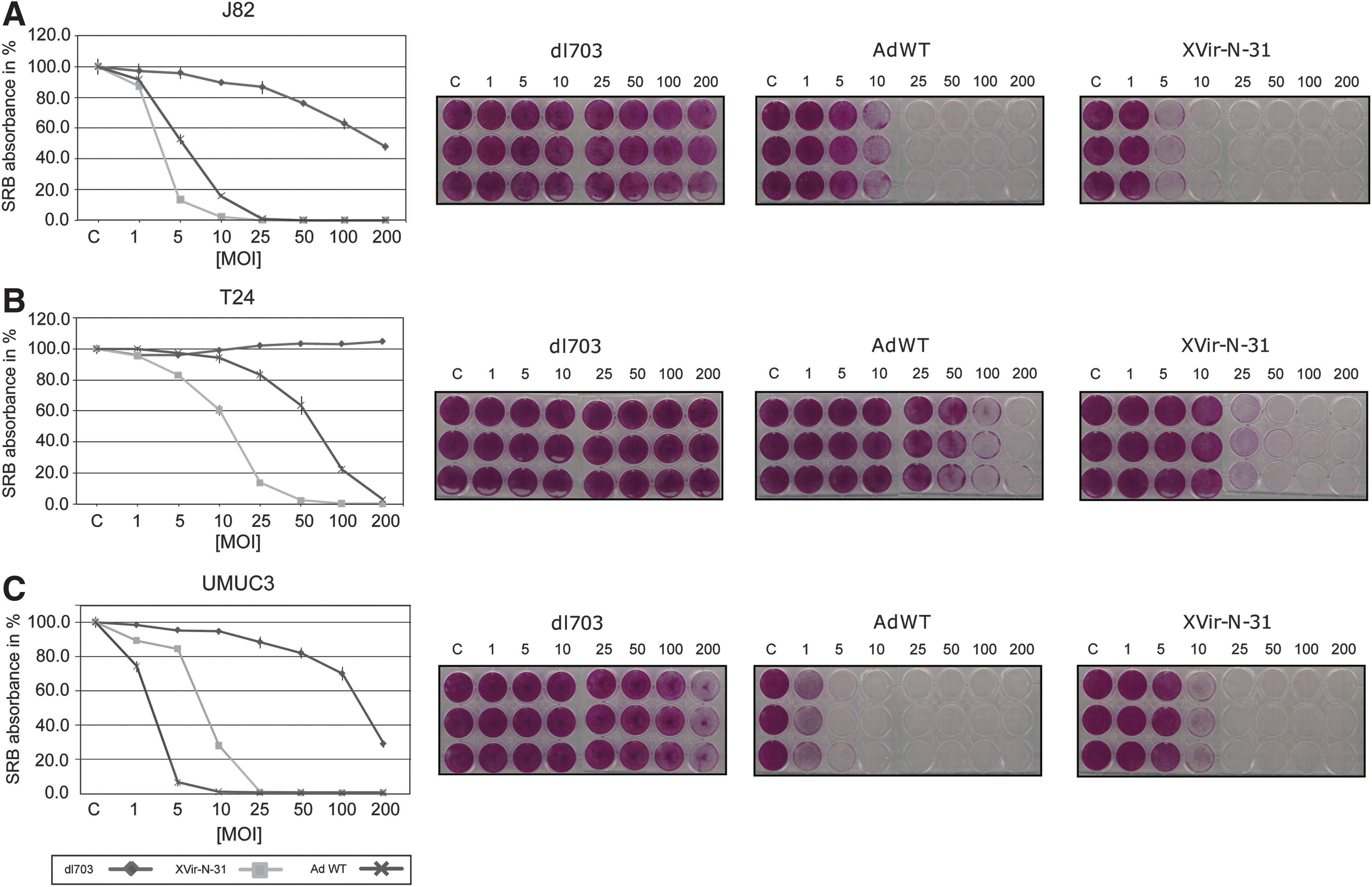
Cell-killing capacity of XVir-N-31, AdWT, and dl703 in bladder cancer cell lines.
Replication and infectious particle formation of XVir-N-31
Strong replication of XVir-N-31 in infected cells leading to the production of new viral particles to re-infect nearby cancer cells is a key factor in the principle of virotherapy. We analyzed replication of XVir-N-31 by qPCR and compared it to AdWT as a positive control and dl703 (Fig. 3). XVir-N-31 showed high replication levels in all three cell lines. Hexon and fiber DNA expression increased around 100–1,000 fold between 4 and 48 h post infection. However, the replication level of AdWT was higher in all cell lines compared to XVir-N-31. This finding can be explained by the E1A13S-deletion of XVir-N-31, which makes viral replication specific to cancer cells but leads to a reduced replication rate. In addition, a titer test based on expression of the hexon protein was used to quantify viral particles in T24, J82, and UMUC3 cells. Efficient viral particle formation was measured in all three cell lines with levels higher than 2 × 108 pfu/mL in J82 and UMUC3 cells. In T24 cells, particle formation was lower, with titers of 4 × 107 pfu/mL (Fig. 3).
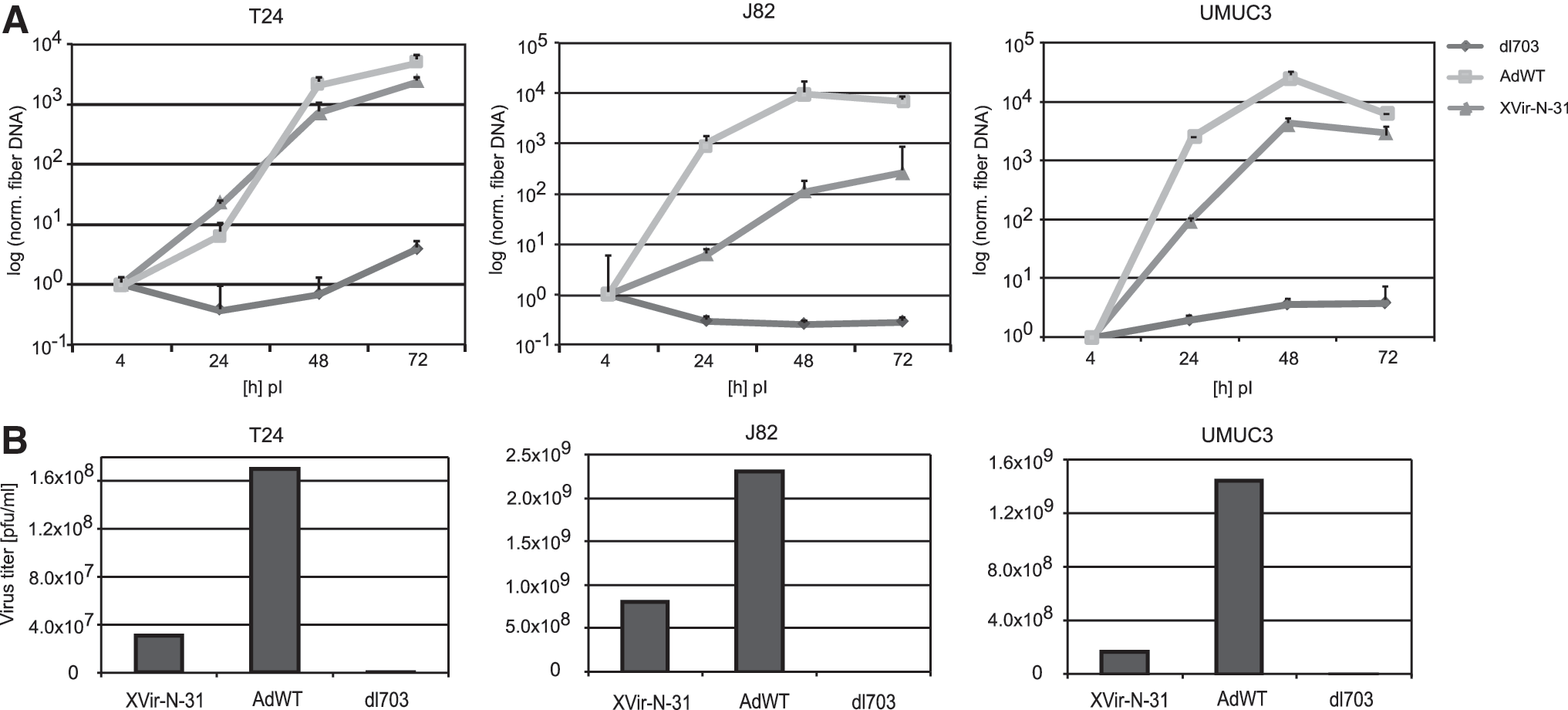
Replication and particle formation of XVir-N-31, AdWT, and dl703 in bladder cancer cell lines.
Analysis of cell death
To investigate if XVir-N-31 induces an ICD, we measured the release of HMGB1 and Hsp70 after viral infection. HMGB1 and Hsp70 are DAMPs and serve to characterize the induction of ICD. 34,35 Our experiments showed an elevated release of HMGB1 and Hsp70 upon XVir-N-31 infection in comparison to the control and AdWT-infected cells in all three cell lines, although the effect was minor in UMUC3 cells (Fig. 4A). Interestingly, doxorubicin, which is known as a potent ICD inducer, 36,37 showed lower values of HMGB1 and Hsp70 release than XVir-N-31, demonstrating the strength of XVir-N-31 to trigger ICD.
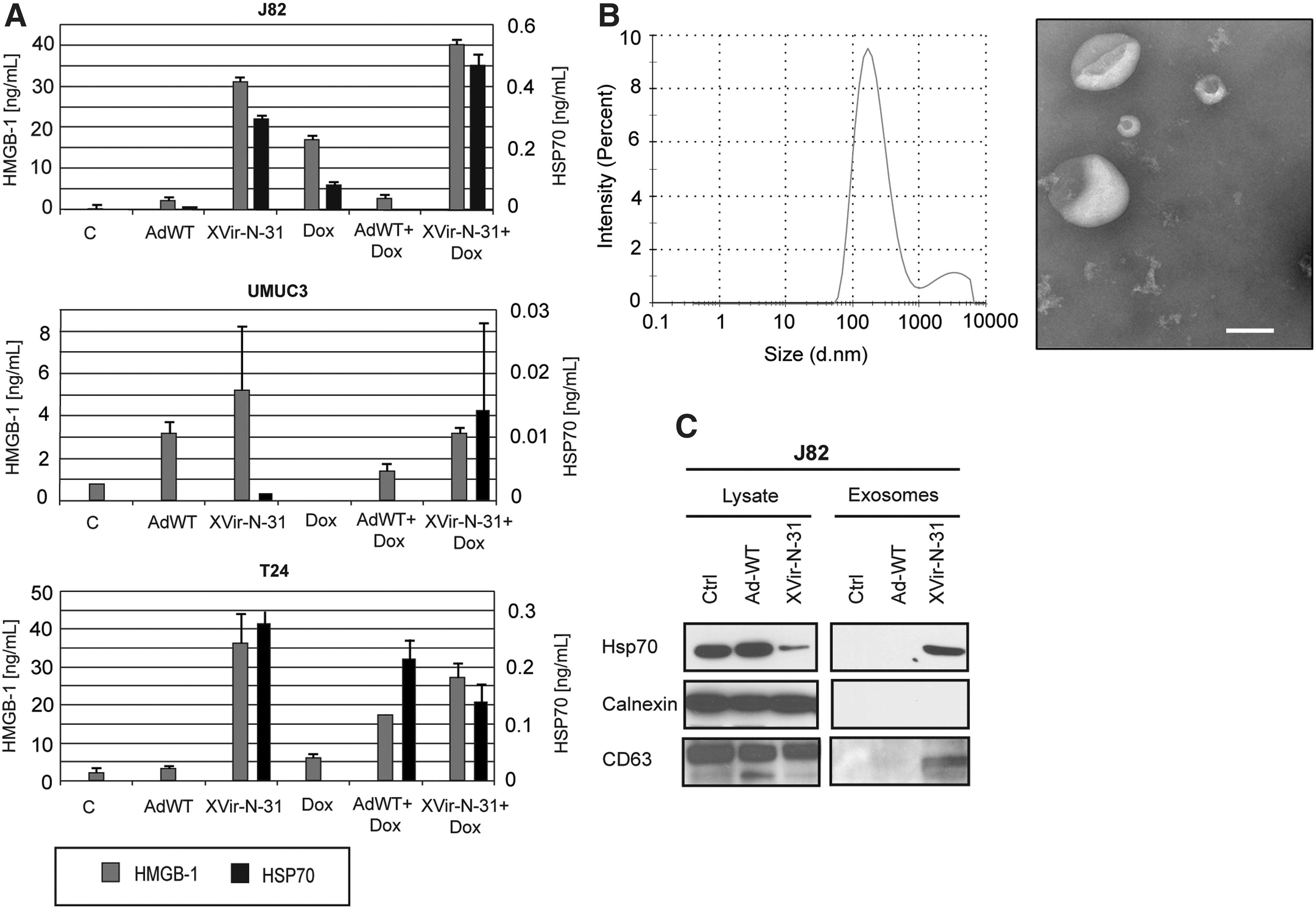
Induction of immunogenic cell death.
We next isolated exosomes and analyzed them for Hsp70 by immunoblotting. As indicated by the presence of the exosomal marker CD63, only infection with XVir-N-31, but not with AdWT, led to sufficient formation of exosomes that were Hsp70 positive (Fig. 4C). Interestingly, Hsp70 expression was increased in cellular lysate upon AdWT infection. To investigate the presence of exosomes in more detail, we expanded our analysis using DLS and TEM. DLS showed an average size of 180–190 nm (Fig. 4B). The discrepancy from the expected size of 100 nm for exosomes can be explained by the polydispersity index values of 0.3, which indicate a slightly polydispersed suspension. Larger particles scatter more light and strongly influence the measured particle size. 38 The size of vesicles measured by TEM was around 100 nm (Fig. 4B). Importantly, the very low number of exosomes in AdWT-infected cells, as seen by TEM, confirmed the absence of exosomes by immune blotting.
XVir-N-31 affects tumor growth in an orthotopic mouse model
Based on our encouraging in vitro results, we generated an orthotopic bladder tumor model for therapy with XVir-N-31. We established an ultrasound-guided, intramural bladder cancer model. Representative ultrasound images showing the steps of the intramural tumor cell injection starting with the visualization of the bladder (1), injection of the needle into the mouse (2), and injection of PBS into the lamina propria to generate an artificial space (3), followed by injection of tumor cells into the generated space (4–6) (Fig. 5A). At day 4 after inoculation, all animals showed tumors detectable by ultrasound. H&E sections from tumor samples revealed tumors that were covered by the urothelium showing correct intramural injections (Fig. 5B). Furthermore, we also detected invasive growth in and through the muscle layer (Fig. 5C). When the tumors reached a size of roughly 20 mm3, mice were randomized into two groups: a virus group that was treated with 3 × 109 viral particles of XVir-N-31, and a control group treated with PBS as depicted in Fig. 6A. The ultrasound images (Fig. 6B) refer to various steps of the procedure: (1) empty bladder; (2–4) bladder tumor growth on day 4, 6, 10; (5) intratumoral virus application; and (6) remaining tumor at day 28 after virus application. Virotherapy led to a significant reduction of the mean tumor growth rate compared to PBS-treated control at day 17 (mean tumor volume of control group [214.6 mm3] and virus-treated group [124.1 mm3], p = 0.047), corresponding to a tumor growth inhibition of 42% (Fig. 6C). Figure 6D shows H&E-stained sections of a remaining tumor 11 days after virotherapy.
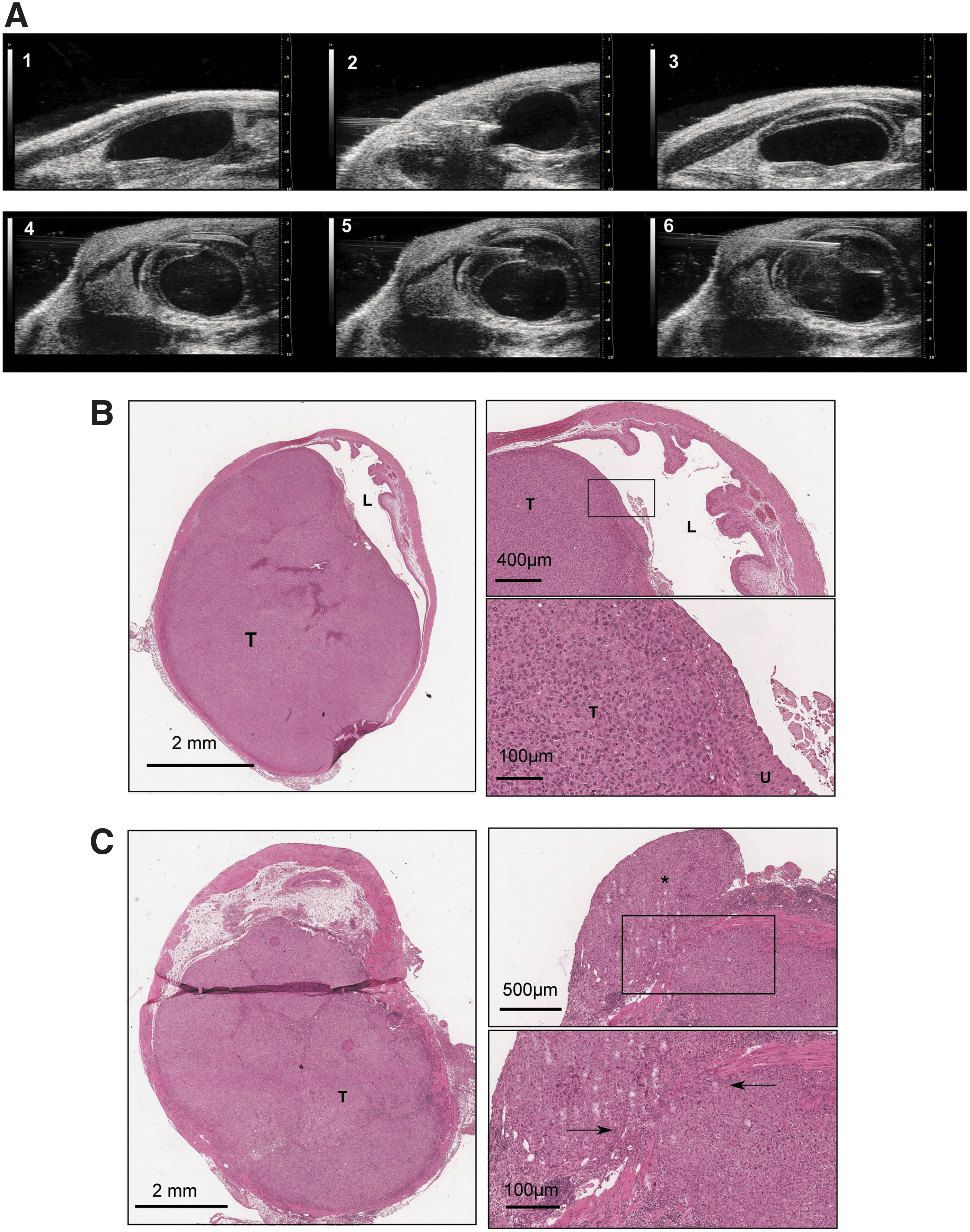
Establishment of an ultrasound-based orthotopic bladder cancer model.

Establishment of an ultrasound-guided injection of XVir-N-31.
Additionally, we performed immunohistochemistry staining for adenoviral hexon, Ki-67, and F4/80. Immunohistochemistry for adenoviral hexon was positive in virus-treated tumors, but not in PBS-treated tumors, confirming correct application and presence (and replication) of XVir-N-31. The Ki-67 staining was positive for the majority of tumor cells, while tumor stroma and muscle cells were Ki-67 negative, indicating the high proliferation rate of the tumor cells. In addition, the invasive capacity of this tumor model is demonstrated by single Ki-67–positive tumor cells migrating into the muscle layer (Supplementary Fig. S1; Supplementary Data are available online at
Discussion
Virotherapy has recently shown impressive results in clinical studies with an excellent safety profile. 39 Particularly in combination with immunotherapeutic approaches, virotherapy could be a novel therapeutic option for the treatment of bladder cancer and has potential for clinical success. 12,13 The possibility of direct virus injection into the tumor by ultrasound guidance or by endoscopic procedure allows high local concentrations and a low systemic exposure, 23 making oncolytic virotherapy particularly attractive as a new treatment strategy for MIBC. Currently, multiple clinical trials are investigating the use of oncolytic viruses constructed from a wide range of viruses in urological malignancies, 40 including the oncolytic adenovirus CG0070 expressing GM-CSF that has completed a human phase I study and is now being tested in a clinical phase II/III study for NMIBC. 41,42 Therefore, XVir-N-31 can be suggested for use in intravesical instillation for treatment of NMIBC. Furthermore, even wild-type vaccinia virus can be administered safely into the bladder. 43 Besides mild local toxicity, no serious treatment-related side effects were reported. 23,43
To ensure safety and efficiency, an oncolytic virus used against bladder cancer should show high specificity and strong oncolytic activity. Our results indicate that YB-1–based virotherapy fulfills these requirements, and they support the existing view that YB-1 is a novel molecular target in bladder cancer and can be used in context of an YB-1–based virotherapeutic approach. 17 –19 However, little attention has been given to the induction of ICD by OVs, although ICD possesses the capacity to generate a systemic immune response 11 and is likely important to achieve the optimal efficacy of OV-based cancer immunotherapies. 7 –9 It is widely accepted that OVs induce ICD mainly through induction of autophagy, 44 –46 which plays a crucial role in antigen processing. 47 Since AdWT showed no significant release of HMGB1 and ATP or calreticulin surface exposure, suggesting that AdWT has developed mechanisms to counteract ICD, 46,48 we analyzed XVir-N-31 and AdWT in the context of ICD induction.
To characterize ICD by XVir-N-31, we measured the release of the DAMPs HMGB1 and Hsp70. 34,35 These molecules are released from dying cancer cells and bind to TLR-449 or CD91-receptors 50 on dendritic cells, which then can activate T lymphocytes to kill tumor cells. 10 Our data demonstrate that infection with AdWT did not result in an increase in HMGB1 and Hsp70 release after infection, whereas infection with XVir-N-31 did. It is obvious to suggest that AdWT suppresses release of DAMPS, thereby causing an ICD inconspicuous phenotype in order to prevent host immune response. E1A-mediated induction of Hsp70 post virus infection has previously been demonstrated. 51 This observation is in line with our results obtained with immunoblotting, which show increased levels of intracellular Hsp70 after infection with AdWT, but not with XVir-N-31. This result indicates that E1A13S is involved in regulation of Hsp70 expression (data not shown).
Further analyses showed that Hsp70 is released in exosomes upon XVir-N-31 infection. This finding might be of importance since several studies have shown increased immune activation by exosomal Hsp70 that activates NK cells. 52 –54
Thus, these results suggest that treatment with XVir-N-31 could cause a stronger anticancer immune response than AdWT and thereby should suppress tumor growth more efficiently. This concept is supported by a recent study from Workenhe et al. 7 that demonstrates this strategy well. However, further studies in an appropriate (e.g., syngeneic mouse model) are certainly required to verify if this observation helps activate the immune system against the tumor. Finally, since adenoviral proteins modulate the level of apoptosis/autophagy, 46 the present deletions in XVir-N-31 (E1A13S and E1B19k) could be suggested as contributing to the observed increase of ICD.
Mouse models for human bladder cancer are crucial in translational research. They are applicable in the investigation of new therapeutic approaches. Several animal models for bladder cancer have been described. 55 Heterotopic xenograft models are generated by non-orthotopic injection of tumor cells under the skin. This method is fast and easy in handling but does not represent the organ-specific physiology and natural environment within the mouse. Development of orthotopic bladder cancer models needs surgical implantation, instillation, or ultrasound-based injection of tumor cells. Surgical injection is time consuming, risky, and not reliable. Instillation of tumor cells needs pretreatment of the bladder, and the tumor uptake is unpredictable. 24 The ultrasound-based model allows injection of tumor cells into the lamina propria and leads to the formation of invasive bladder tumors. It simulates human pathology and tumor biology within the natural environment most closely, and tumor uptake rates are almost 100%. 25 In addition, this method allows direct injection of the virus into the established tumor. Immunohistochemistry staining showed massive F4/80-positive cells in the tumor, demonstrating the presence of (probably activated) macrophages. Although we did not observe differences in treated and untreated tumors, this presence is of great importance since infiltration of macrophages together with the vascularization architecture of the tumor demonstrates that this model closely reflects the physiological characteristics of human bladder cancer.
Our results in this sophisticated orthotopic model showed that XVir-N-31 as monotherapy slowed tumor growth significantly compared to the control, although we used a relatively small number of animals per group (Fig. 6C). We noticed massive cell death in areas with adenoviral hexon-positive cells, demonstrating the oncolytic activity of XVir-N-31 in vivo (Fig. 6E). However, a complete tumor regression in vivo was not achieved. This outcome is in line with published data, demonstrating the difficulty of completely eradicating the highly proliferating xenograft tumor in nude mice in which an immune response against the tumor is not present. 56,57 Accordingly, a functioning immune system is necessary to achieve complete tumor remission. Furthermore, the nature of solid tumors, including their extracellular matrix, increased intratumoral pressure, and the meshwork of secreted proteins, represents a natural barrier that complicates even virus dispersal and widespread tumor cell infection. 58 In addition, since nuclear YB-1 is a prerequisite for viral replication of XVir-N-31, combination treatments with chemotherapy or radiation therapy that translocate YB-1 into the nucleus 59,60 are warranted and can help to further improve the therapeutic activity.
Considering the capacity of XVir-N-31 to increase ICD, it would be highly exciting to verify our strategy of YB-1–based induction of ICD in an immuno-competent mouse model. Regarding the latest results from Lichty et al. 6 and Ribas et al., 12 we hypothesize that a combination of XVir-N-31 with an immune checkpoint blockade would be beneficial and might help to further increase the immunological effects. For this purpose, we have already equipped XVir-N-31 with an anti-PD-L1 antibody and are currently focusing on establishing a syngeneic orthotopic bladder cancer mouse model that allows the investigation of immunological effects of this highly promising combinatorial approach.
Footnotes
Acknowledgments
This work was supported by the German Research Foundation (DFG) under grant no. HO-1482/7-1.
Author Disclosure
Dr. Per Sonne Holm is CEO and co-founder of XVir Therapeutics GmbH, Munich, Germany. The authors declare that they have no competing interests.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.