
Abstract
Adeno-associated viral vectors are showing great promise as gene therapy vectors for a wide range of retinal disorders. To date, evaluation of therapeutic approaches has depended almost exclusively on the use of animal models. With recent advances in human stem cell technology, stem cell–derived retina now offers the possibility to assess efficacy in human organoids in vitro. Here we test six adeno-associated virus (AAV) serotypes [AAV2/2, AAV2/9, AAV2/8, AAV2/8T(Y733F), AAV2/5, and ShH10] to determine their efficiency in transducing mouse and human pluripotent stem cell–derived retinal pigment epithelium (RPE) and photoreceptor cells in vitro. All the serotypes tested were capable of transducing RPE and photoreceptor cells in vitro. AAV ShH10 and AAV2/5 are the most efficient vectors at transducing both mouse and human RPE, while AAV2/8 and ShH10 achieved similarly robust transduction of human embryonic stem cell–derived cone photoreceptors. Furthermore, we show that human embryonic stem cell–derived photoreceptors can be used to establish promoter specificity in human cells in vitro. The results of this study will aid capsid selection and vector design for preclinical evaluation of gene therapy approaches, such as gene editing, that require the use of human cells and tissues.
Introduction
A
The availability of pluripotent stem cells (PSC) that can be effectively differentiated into three-dimensional (3D) retinal organoids has brought many new opportunities for accelerating the development of novel therapies for wide range of retinal conditions, including inherited retinal dystrophies caused by defects in either photoreceptors or the RPE. Numerous reports have described the differentiation of both mouse and human pluripotent stem cells into photoreceptors and RPE, for transplantation 18 –25 and in vitro disease modeling studies, using patient-derived induced pluripotent stem cells (iPSCs). 26 –29 The combined use of relatively new stem cell technologies together with the more established use of recombinant AAV vectors for gene therapy represents a unique opportunity for advancing our ability to understand and treat inherited retinal disorders.
A growing number of studies are utilizing PSC-derived retinal organoids and efficient gene transfer to these tissues is of increasing interest. We have previously observed that AAV vectors are more effective at mediating gene transfer than either lentiviral vectors or physical methods such as transfection or electroporation and have therefore previously used AAV vectors to mark mouse and human PSC-derived photoreceptor precursors in order to isolate and purify them for use in transplantation studies. 25,30,31 Human PSC-derived retinal organoids provide access to large numbers of normal and diseased samples and also offer a new platform to evaluate gene correction by gene therapy specifically in human retinal cells. The precise modification of the genome can be achieved by well-described gene editing technologies, such as zinc fingers, TALENS, and CRISPR/Cas9. 32,33 The in vivo delivery of these editing tools to the mouse retina using AAV has now been demonstrated in a number of recent studies (for review see Peddle and Maclaren, 2017 34 ). However, as a prelude to in vivo gene editing, double strand breaks efficiency, cutting specificity, and efficiency of gene integration when a donor DNA is used need to be tested first in cells in vitro. While various cells lines can be used, mouse and human retinal organoids provide more appropriate tools to evaluate gene editing than primary cell lines. Furthermore, in the case of patient-derived human organoids, these provide unlimited number of diseased cells to develop and test the most efficient gene correction technology. While AAV vectors are promising tools for mediating gene editing in retinal organoids, it is first necessary to establish which AAV serotypes most efficiently transduce these tissues in vitro. This study therefore assesses the tropism of AAV vectors in stem cell–derived retinal organoids. We sought to determine the relative transduction efficiencies and toxicities of six AAV serotypes [AAV2/9, AAV2/8, AAV2/8T(Y733F), AAV2/5, AAV2/2, and ShH10(Y445F)] in mouse and human PSC-derived photoreceptor and RPE cells. These six serotypes were chosen for comparison in vitro because of their widespread use in vivo. AAV2/8, AAV2/2, and AAV2/5 have been used in ocular gene therapy trials, while AAV2/9 and the two engineered capsids, ShH10 and AAV2/8T(Y733F), mediate particularly efficient in vivo transduction.
Materials and Methods
Mouse embryonic stem cell retinal differentiation culture
A CCE mouse embryonic stem cell (ESC) line (129/SvEv; a kind gift of Professor E. Robertson, University of Oxford, Oxford, UK) was maintained as previously described. 35 For 3D retinal differentiation, 3 × 104 dissociated ESCs were resuspended per milliliter of differentiation medium (Glasgow minimum essential medium containing 1.5% knockout serum replacement, 0.1 mM non-essential amino acids solution, 1 mM pyruvate, 0.1 mM 2-mercaptoethanol), plated into 96-well, low-binding plates (Corning) and incubated at 37°C, 5% CO2. Embryoid body cell aggregates (EBs) formed within 24 hours; on day 1 of culture, growth factor reduced Matrigel (GIBCO) was added to each well to give a final concentration of 2%. For whole EB retinal differentiation toward photoreceptor cell fate, EBs were transferred into retinal maturation medium (Dulbecco's modified Eagle's medium [DMEM]/F12 Glutamax containing N-2 supplement and penicillin/streptomycin [pen/strep]) at day 9, plated in low-binding plates at a density of 6 EBs/cm2, and incubated at 37°C, 5% CO2. The media was changed every 2–3 days, with the addition of 1 mM Taurine (Sigma) and 500 nM retinoic acid (Sigma) from day 14 of culture onwards. For whole EB retinal differentiation towards RPE cell fate, EBs were treated with 500 nM working solution of 6-bromoindirubin-3′-oxime (BIO; Sigma), from day 5 onward. For monolayer cultures of RPE, pigmented regions of the BIO-treated EBs were manually dissected with a 21G needle and platted these on laminin-coated chamber slides.
Human ESC maintenance and retinal differentiation culture
The human embryonic and iPS stem cell lines (H9 and IRM90-4 from Wicell) were maintained on feeder free conditions on E8- and geltrex-coated 6-well plates. RPE differentiation protocol was adapted from previous published protocols.
19,36
Briefly, hPSCs were dissociated using a dispase and collagenase solution. PSC clumps were collected and resuspended in E8 media. The cell clumps were then transferred to a low-binding 10-cm plate to form floating EBs. Differentiation media was changed to EB media (DMEM/F12 (1:1), knockout serum replacement, minimum essential medium (MEM) nonessential amino acids,
For retinal neuroepithelia differentiation we used a previously described protocol. 25 Briefly, human PSCs were maintained until confluent when media without fibroblast growth factor (FGF) was added to cultures for 2 days. Proneural induction media (advanced DMEM/F12, MEM nonessential amino acids, N2 supplement, 100 mM glutamine, and pen/strep) was added until optic vesicles were observed. Vesicles were manually excised and kept in 96-well plates in retinal differentiation media (DMEM, F12, pen/strep, and B27 without retinoic acid). At 6 weeks of differentiation, medium was supplemented with FBS, taurine, and glutamax and at 10 weeks retinoic acid was added.
Production of recombinant AAV serotypes and in vitro transduction analysis
A pD10/cytomegalovirus (CMV) promoter–green fluorescent protein (GFP) construct containing AAV-2 inverted terminal repeat was used to generate AAV2/9, AAV2/8, AAV2/8(Y733F), AAV2/5, AAV2/2, and ShH10(Y445F) CMV.GFP viruses. A pD10/Rhodopsin promoter–GFP and a pD10/2.1PRM/L-opsin promoter–GFP was used to generate viruses. Recombinant AAV2/2 serotype particles were produced through a previously described triple transient transfection method HEK293T cells. 37 AAV2/8, AAV2/8(Y733F), AAV2/5, AAV2/2, and ShH10(Y445F) serotypes were bound to an AVB Sepharose column (GE Healthcare) and eluted with 50 mM glycine pH 2.7 into 1 M Tris pH 8.8. AAV2/9 was purified by size separation on a Sephacryl S300 column, followed by anion exchange chromatography using a POROS 50 HQ column, eluting the vector in 20 mM Bis-Tris propane, 20 mM Trizma Base and 0.24 M NaCl pH9. Vectors were washed in 1 × phosphate-buffered saline (PBS) and concentrated to a volume of 100–150 μL using Vivaspin 4 (10 kDa) concentrators. Viral genome titers were determined by quantitative real-time PCR using a probe-based assay binding the SV40 poly-adenylation signal. Amplicon-based standard series of known amounts were used for sample interpolation. Final titers were expressed as vg/mL.
SV40 Forward primer: 5′-AGCAATAGCATCACAAATTTCACAA-3′.
SV40 Reverse primer: 5′-AGATACATTGATGAGTTTGGACAAAC-3′.
SV40 Probe: FAM-5′-AGCATTTTTTTCACTGCATTCTAGTTGTGGTTTGTC-3′-TAMRA.
The same number of EBs or RPE cells per well were used in all experiments (approximately 1.5 × 106 EB cells and 2 × 106 RPE cells per well) and infected with 1.2 × 1011 viral particles per well in appropriated retinal medium. Estimated gMOI of 8000 and 6000.
Flow cytometry analysis
For flow cytometry experiments, mouse whole EBs (wEBs) were dissociated at day 29 of culture into a single cell suspension via a modified protocol using reagents from the papain-based Neurosphere Dissociation Kit (Miltenyi Biotec). Cells were counted and resuspended in 1% bovine serum albumin (in PBS) to a concentration of 1 × 107 cells per mL, and staining was performed with 100 μL aliquots. For surface marker analysis, an antibody against mouse CD73 (APC-conjugated rat immunoglobulin 1, clone TY/11.8; Miltenyi Biotec) was added at a 1:75 dilution and incubated for 30 minutes at 4°C. Cells were washed once in 1 × binding buffer (eBioscience). For subsequent viability analysis, Annexin V-eFluor 450 (eBioscience) was added to samples at a 1:20 dilution (in 1 × binding buffer) and incubated for 15 min at room temperature. Cells were washed once in 1 × Binding Buffer and resuspended in PBS. DRAQ7 (BioStatus) was then added to the samples at a final concentration of 50 ng/mL for 5 min at room temperature before analysis.
Cells were analyzed using FlowJo software. Background fluorescence was measured using unstained cells, and single-stained controls were used to set gating parameters between positive and negative populations. Small debris, cell fragments, and aggregates were excluded from analysis on the basis of Annexin V and live-dead dye nonfluorescence (double negative population), followed by forward and side scatter (measuring cell size and granularity respectively). For cell surface marker analysis, dead cells were excluded from analysis on the basis of live-dead dye fluorescence as well.
For cell sorting experiments fluorescence-activated cell sorting was performed on a BD Influx Cell Sorter™ (BD Biosciences) fitted with a 200 mW 488 nm blue laser to excite GFP. GFP was collected using the 488-530/40 nm detector. A 70 micron nozzle at 30 psi was used and cells were collected on a 1:1 FBS/ EBSS solution.
Immunohistochemistry
For wEB and BIO-treated wEB experiments, titer-matched viruses were added at day 22 of differentiation and wEBs were collected for immunohistochemistry analysis at day 29 of culture. For adherent cultures of RPE, viruses were added between day 20 and 22 and chamber slides were fixed 7 days later. Whole EBs and eye cups were fixed for 1 hour in 4% paraformaldehyde and embedded in optimal cutting temperature compound (RA Lamb). Cryosections were cut (18 μm thick) and all sections were collected for analysis. For immunohistochemistry, sections were blocked in 5% goat serum and 1% bovine serum albumin in PBS. Primary antibody (Supplementary Table S1; Supplementary Data are available online at
Image acquisition
Images were acquired by confocal microscopy (Leica DM5500Q). A series of XY optical sections, approximately 1.0 μm apart, throughout the depth of the section were taken and built into a stack to give a projection image. Leica Microsystems LAS AF image software was used.
Statistical analysis
All means are presented ± standard deviation, unless otherwise stated; N, number of animals or independent experiments performed; n, number of eyes or EBs examined, where appropriate. For quantification assessment by flow cytometry and cell counting of transduction efficiency, statistical analysis is based on at least three independent experiments. Statistical significance was assessed using Graphpad Prism 6 software; *p < 0.05,** p < 0.01, and ***p < 0.001. Appropriate statistical tests were applied, including t-test and ANOVA with Tukey's correction for multiple comparisons.
Results
AAV transduction of mouse PSC-retinal cells
AAV transduction of mouse ESC-derived whole embryoid bodies (wEBs)
First, we sought to determine how efficiently we could transduce retinal organoids using AAV vectors. We have described previously the 3D differentiation of retinal cells from mouse ESC-derived (mESC) wEBs. These structures contain both neural and glial cells, as well as retinal neuroepithelium containing mostly photoreceptors and fewer less well-developed retinal interneurons. 30 We tested six different AAV serotypes, each driving enhanced GFP under the control of a CMV promoter. A schematic of the 3D differentiation and vector transduction protocol is shown in Fig. 1a. At day 22 of differentiation, titer-matched AAV vectors (1.2 × 1011 vg/well) were added to wEB cultures (6 wEBs in 0.3 mL medium) and the presence of GFP+ cells was assessed 7 days later. The AAV serotypes exhibited different patterns of tropism in vitro (Fig. 1b–g). Low magnification images of wEB sections at day 29 of culture showed GFP+ cells not only in regions of retinal neuroepithelium (Fig. 1b, arrow head), but also in other regions of the wEBs (Fig. 1b, arrow). The number of GFP+ transduced cells was analyzed by flow cytometry. The percentage of GFP+ cells was significantly higher (53% ± 8.7) following transduction by ShH10 than by the other serotypes tested. AAV2/9 (16% ± 7.5), AAV2/8 (20% ± 4), AAV2/8T (16% ± 1.3), and AAV2/5 (12% ± 5.3) serotypes provided similar transduction efficiency, and AAV2/2 transduced just 6% (± 2.6) of cells (Fig. 1h; p < 0.001, ANOVA; n = 3 independent differentiation cultures and Supplementary Fig. S1a and b). Furthermore, ShH10.CMV.GFP+ labelled cells were not only more numerous, but also had a stronger and brighter GFP expression (see Supplementary Fig. S1c for normalized median fluorescence intensity fold-values). Using a high vector titer can be toxic to target cells, 38 and thus it is also important to establish cell viability following transduction. To do this, we quantified by flow cytometry the percentage of early and late apoptotic (live dead−/AnnexinV+ and live dead+/AnnexinV+, respectively), necrotic (live dead+/AnnexinV−), and live cells (live dead−/AnnexinV−) in cultures transduced with each of the different vectors. No significant differences in cell viability were noted, regardless of the serotype used (Supplementary Fig. S1d).
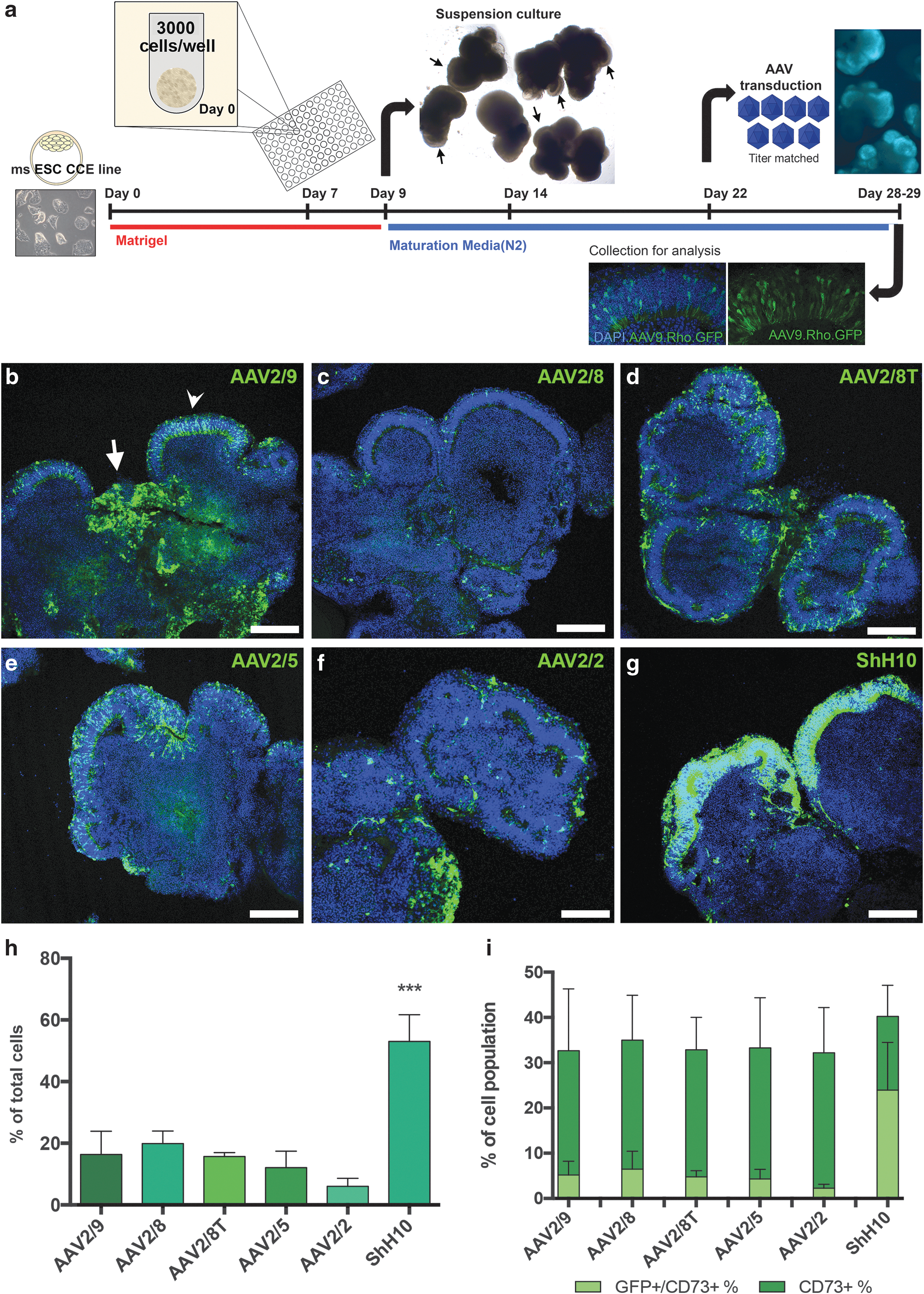
Establishing AAV tropism in mouse ESC-derived wEBs.
Targeting photoreceptor cells with AAV vectors is of particular interest, as these cells contain the primary defect in the majority of retinal degenerations. We therefore investigated the ability of the six serotypes to transduce mESC-derived photoreceptors specifically. We demonstrated previously that, similar to photoreceptors from the postnatal retina, 39,40 mESC-derived photoreceptors express a CD73 surface marker. 41 The percentage of CD73+ photoreceptor cells did not vary significantly between the different transduced cultures (Fig. 1i). However, the number of CD73+/CMV.GFP+ photoreceptor cells was significantly greater when cells were transduced with the ShH10 (24% ± 10.5%), compared with any of the other five serotypes (Fig. 1i ANOVA, p < 0.0001; n = 3 independent differentiations and Supplementary Fig. S2a–c). Furthermore, no significant difference in the number of viable CMV.GFP+/CD73+ photoreceptors was detected between any of the six serotypes (Supplementary Fig. S2d).
ShH10 has been previously reported to specifically target astrocytes and Müller Glia following intravitreal (i.v.) injections. 42 We performed i.v. injections of the six CMV.GFP viruses into wild-type postnatal day 6 pups, a similar stage to days 22–24 of differentiation, to assess the potential of these viruses to transduce different cell types in the postnatal neural retina (Supplementary Fig. S3). Similar to previous studies, 42 we found that ShH10.CMV.GFP specifically transduces astrocytes and Müller glia following i.v. injections in vivo (Supplementary Fig. S3f). However, immunohistochemistry analysis of ESC-derived wEBs transduced with ShH10.CMV.GFP demonstrated that neuroepithelium regions contained not only CRALBP+/GFP+ Müller glia cells, but also many Recoverin+/GFP+ photoreceptors (Supplementary Fig. S4). This is most likely due to increased viral vector accessibility in vitro. In the 3D wEB cultures, the presumptive inner nuclear layer is thinner compared with the postnatal retina, and these structures may lack astrocytes and the inner limiting membrane, both present at the vitreal surface of the postnatal retina. Our results demonstrate that ShH10 vectors are able to mediate significantly improved transduction of mouse ESC-derived cells, including photoreceptors, compared with the other serotypes tested.
AAV transduction of mouse ESC-derived rods and cones using cell-specific promoters
To address questions of retinal and photoreceptor development using mouse ESC-derived retinal organoids it is often necessary to target rod and/or cone photoreceptor cells specifically. To determine the transduction efficiency of rods and cones in vitro, we used either a bovine rhodopsin promoter 43 to obtain rod-specific expression or a human L-opsin cone promoter (PR2.1opsin.GFP) 44 to obtain cone-specific expression. We compared the transduction efficiency of an AAV2/9 vector with ShH10. Both AAV2/9.Rhop.GFP and ShH10.Rhop.GFP robustly and specifically labelled rod photoreceptors in 3D cultures, as demonstrated by GFP colocalization with the rod-specific protein Rhodopsin (Fig. 2a). In our mouse 3D retinal organoid cultures we do not observe M/L opsin positive cones. 31 However, subretinal injection of AAV2/8.PR2.1opsin.GFP virus into the mouse adult wild-type retina results in labeling of S cones, as well as M/L cones (Supplementary Fig. S5). We therefore examined the efficiency of AAV2/9 and ShH10.PR2.1opsin.GFP transduction of S cones by colocalization with S-opsin (blue opsin) (Fig. 2b). In wEBs transduced with either vector, GFP fluorescence colocalized with S-opsin staining. The percentage of Rhop.GFP+ rods and PR2.1opsin.GFP+ cones was significantly higher in wEBs transduced with ShH10 (24% ± 6.8 and 7% ± 1.3, respectively), compared with AAV2/9 (17% ± 5.0 and 4% ± 1.0, respectively) (Fig. 2c and Supplementary Fig. S6; unpaired t-test; rods p < 0.05, n = 9; cones p < 0.01, n = 4 independent differentiations). No significant differences in viability were detected between any of the vectors tested (Fig. 2d).

Testing promoter specificity in mouse ESC-derived photoreceptors.
AAV transduction of mouse ESC-derived RPE
To investigate the tropism of AAV serotypes with respect to mESC-derived RPE, we used a 3D suspension differentiation protocol in which the Wnt agonist BIO was added from day 5 of culture. A schematic of RPE differentiation is shown in Fig. 3a. Using this method, neuroepithelia that under non-BIO control conditions 20 would normally generate thick neuroepithelium of retinal origin and a small number of pigmented RPE cells (Fig. 3b, black and white arrows respectively), instead differentiate exclusively into thin, convoluted RPE layers (Fig. 3b, white arrows). First, we analysed the ability of the six AAV serotypes to transduce RPE-derived from BIO-treated wEBs cultured in suspension. Sections of BIO-treated wEBs, positive for the early RPE marker Mitf, demonstrated the presence of CMV.GFP+ pigmented cells in all six serotypes tested (Fig. 3c). To quantify the proportion of RPE cells transduced, RPE regions were manually dissected from BIO-treated wEBs at day 14 of culture and plated to form adherent RPE monolayers that were positive for actin-label phalloidin and demonstrated the typical cobblestone-like RPE cell morphology (Fig. 3d). RPE monolayers were then transduced and the percentage of GFP+ RPE cells was quantified by manual cell counting. ShH10 and AAV2/5 achieved similar levels of transduction, with around 70% of RPE cells being GFP+ (71% ± 11.2 and 71% ± 26 respectively). These two serotypes were significantly more efficient at transducing mESC-derived RPE compared with the other four serotypes tested (Fig. 3e; p < 0.001, ANOVA; n = 3 independent differentiations).

Establishing AAV tropism in mouse ESC-derived RPE.
AAV transduction of human PSC-retinal cells
AAV transduction of human PSC-derived RPE
Pluripotent stem cells provide a renewable source of human RPE cells that can be used to study normal and disease mechanisms in vitro. Patient-derived iPSC-derived RPE cells offer a disease modelling platform that may allow the development of new treatments. Here, to assess which AAV serotypes most efficiently transduce human pluripotent stem cell (hPSC)-derived RPE in vitro, we differentiated human ES and iPS cells into RPE using a previously described protocol. 19,36 A schematic of the differentiation and virus infection protocol is shown in Fig. 4a. Titer-matched vectors were added between days 50 and 60 of culture and the number of GFP+ RPE cells was quantified as before, 7 days following addition of vector. Similar to mouse ESC-derived RPE, hPSC-derived RPE cultures formed typical pigmented and polygonal adherent cultures that expressed phalloidin (Fig. 4b). All serotypes were able to transduce adherent cultures of hPSC-derived RPE in vitro, albeit with different efficiencies. The proportion of ShH10 CMV.GFP-transduced hPSC-derived RPE (76% ± 30) was significantly greater than for AAV2/9 (6% ± 3), AAV2/8 (6% ± 6), AAV2/8T (3% ± 2), and AAV2/2 (11% ± 6), but not compared with AAV2/5 (37% ± 18) (Fig. 4c; p < 0.01, ANOVA; n = 3 independent differentiations). Usually, hPSC-derived RPE fields are picked manually from a heterogeneous population of retinal cells and as a result the final population is not always composed entirely of RPE cells. Therefore, to ensure specific targeting of RPE cells in vitro, specific promoters are needed. We have previously described that an optimised RPE65 human promoter can drive GFP expression in hPSC-derived RPE cells. 45 Here, we assessed if the same was true for the well-described human vitelliform macular dystrophy 2 (VMD2) promoter. We found that this promoter can also be used to target RPE in vitro and the percentage of GFP+ RPE cells transduced with the ShH10.VMD2.GFP (79% ± 5%) was not significantly different from cells transduced with ShH10.CMV.GFP (92% ± 8) (Fig. 4d; ANOVA; n = 3 independent differentiations). Furthermore, despite greater levels of GFP expression present in RPE cells transduced with vector containing a CMV promoter than in cells transduced with vector containing the RPE-specific VMD2 promoter, no differences in the viability of the cells were observed (Fig. 4e). Our results demonstrate that AAV vectors can readily transduce human RPE cells in vitro, with ShH10 and AAV2/5 performing significantly better than other commonly used serotypes.
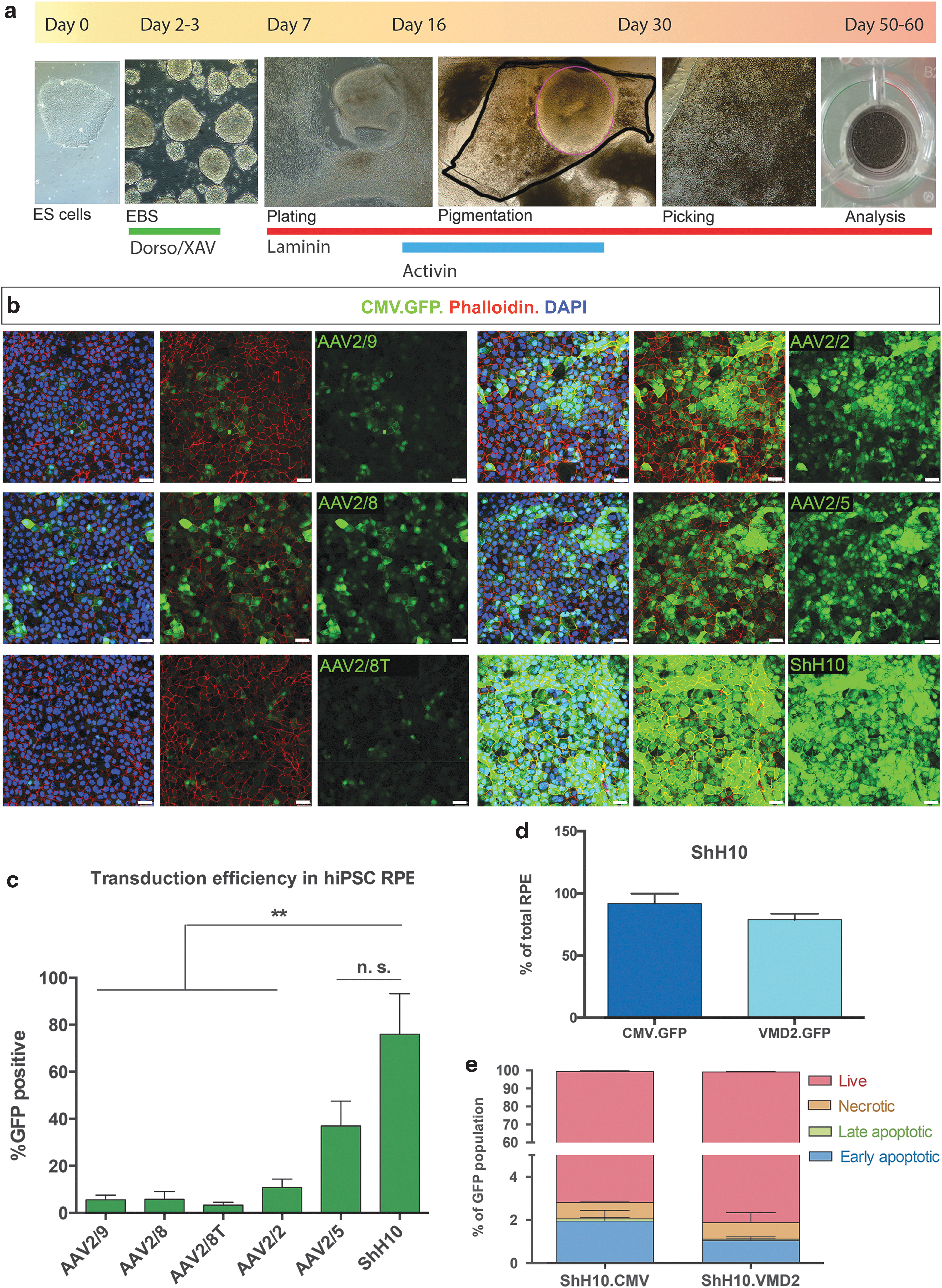
Establishing AAV tropism in human iPS-derived RPE.
AAV transduction of human ESC-derived retinal organoids and photoreceptors
A number of protocols have been reported for the differentiation of hPSCs into retinal tissues and cell types. We have adapted previously described protocols 24,46 to generate retinal neuroepithelia containing a layered neuroretina, including photoreceptors that exhibit rudimentary outer segments and synaptic structures. 25 A schematic of photoreceptor differentiation and virus infection protocol is shown in Fig. 5a. Briefly, human ESCs were kept in maintenance conditions to confluence, before adding FGF negative medium. Islands of pigmented RPE were observed 3–4 weeks later. and continuous retinal neuroepithelium vesicles surrounded by RPE were observed from 4 to 7 weeks of culture. These retinal vesicles were then excised from the 2D cultures and cultured further in suspension. We sought to establish the most efficient AAV serotype to transduce hESC-derived photoreceptors in vitro by testing the same six vectors described above. Titer-matched CMV.GFP AAV vectors were added to hESC-derived organoids at 13 weeks of culture and analysed for the presence of GFP +2 weeks later. All six vectors were able to transduce hESC-derived retinal vesicles. A low magnification image of an AAV2/8.CMV.GFP-transduced vesicle shows a number of GFP+ cells within the vesicle (Supplementary Fig. S7a,a′). Strikingly, however, none of these appeared to be photoreceptors, as identified by costaining for RECOVERIN (Fig. 5b–h; n = 3 independent experiments, N > 18 vesicles). This was the case for all six serotypes, including ShH10, which was able to transduce mouse mESC-derived photoreceptors with very high efficiency. However, when viral transduction was performed 4 weeks later (17 weeks) CMV.GFP+/ RECOVERIN+ photoreceptors were observed in the ONL-like region (Fig. 5i–m). In organoids transduced with ShH10.CMV.GFP, other retinal cells, possibly Müller glia, were also GFP+ (Fig. 5l). Flow cytometric analysis of hESC-derived organoids demonstrated equivalent transduction using AAV2/2 and ShH10 (19% ± 3 and 21% ± 4, respectively), which were significantly greater than all the other vectors tested (Fig. 5n; 4% ± 3 AAV2/9, 4% ± 3 AAV2/8, 5.5% ± 3 AAV2/8T, 4% ± 2.5 AAV2/5; p < 0.001, ANOVA; n = 3 independent differentiations). Therefore, the transduction efficiency of human PSC-derived photoreceptors by AAV vectors appears to depend on the developmental stage of the host photoreceptor cells.

Establishing AAV tropism in human ESC-derived retinal organoids.
Testing promoter specificity in human ESC-derived photoreceptors
One requirement for many retinal gene therapy approaches is the specific targeting of photoreceptor cells that avoids off-target expression. There may also be a requirement to further discriminate between rod and cone photoreceptors. We therefore assessed the ability of our retinal organoid system to test the specificity of expression after AAV-mediated transduction of photoreceptor cells using cone- and rod-specific promoters. Titer-matched vectors containing the cone-specific PR2.1opsin promoter driving enhanced GFP were added between weeks 13 and 14 of culture, and retinal organoids were collected 2 weeks later for immunohistochemistry and flow cytometry. Despite the addition of vectors at early stages in culture (13 weeks), we observed a number of GFP+ cells displaying typical cone morphology, with large inner segments and presumptive synaptic pedicles (Fig. 6a). ShH10.PR2.1.GFP+ cells coexpressed the cone-specific protein, RXRγ, but not the rod photoreceptor-specific photopigment, RHODOPSIN (Fig. 6b and c, respectively), demonstrating that all the transduced cells were indeed cone photoreceptors. To establish the transduction efficiency of the different serotypes and promoter specificity for the different cone photoreceptors subtypes, we tested four different AAV serotypes (ShH10, AAV2/9, AAV2/5 and AAV2/8) in hESC-derived organoids. GFP+ cells were observed in all transduced hESC-derived organoids and, in keeping with their normal position in the human outer nuclear layer (ONL), positive cells were mostly observed at the apical edge of the neuroepithelium. The efficiency of the ShH10.PR2.1 vector (15% ± 5) was similar to AAV2/8.PR2.1 vector (13% ± 2) and was significantly greater than AAV2/9 (3% ± 1), AAV2/2 (2% ± 1), or AAV2/5 (2% ± 1) capsids (Fig. 6d and Supplementary Fig. S7; p < 0.001, ANOVA; n = 4 independent differentiations). To determine in which cone photoreceptor subtypes the PR2.1 promoter is active, we checked for colocalization of GFP with red and green (L+M) opsins (M/L OPSIN) and blue opsin (S-OPSIN). Colocalization of M-OPSIN and GFP+ cells was observed in the neuroepithelium (Fig. 6e, arrow). Colocalization of GFP+ and S-OPSIN cones was also observed, but only in a small proportion of the S cones. These PR2.1opsin.GFP+/S-OPSIN+ cones were observed only in ShH10 and AAV2/8 transduced organoids, but not in AAV2/9, AAV2/2, and AAV2/5 transduced organoids (Fig. 6f, arrow). Next, to test whethher a previously described human rhodopsin promoter 47 is active and drives expression of GFP in PSC-derived rod photoreceptors, we used an ShH10 capsid (ShH10.hRho.GFP). Similar to serotypes bearing the CMV promoter, no GFP+ cells were observed following transduction at 13 weeks, but when administered at 17 weeks in culture, this vector readily transduced rod photoreceptors as observed by widespread GFP expression and colocalization with the rod-specific marker α-rod transducin (GNAT1) (Fig. 6g and g′). Finally, we also observed photoreceptor-specific labelling when using an shh10 vector containing a human rhodopsin kinase promoter fragment 48 (ShH10.hRK.GFP). Despite the sporadic labelling at the time point tested (17 weeks), our results confirm that this promoter fragment is active in both ARRESTIN3+ human cone (Fig. 6h and h′) and GNAT1+ rod photoreceptors (Fig. 6i and i′) and is therefore an appropriate promoter for gene therapies targeting both types of photoreceptors.
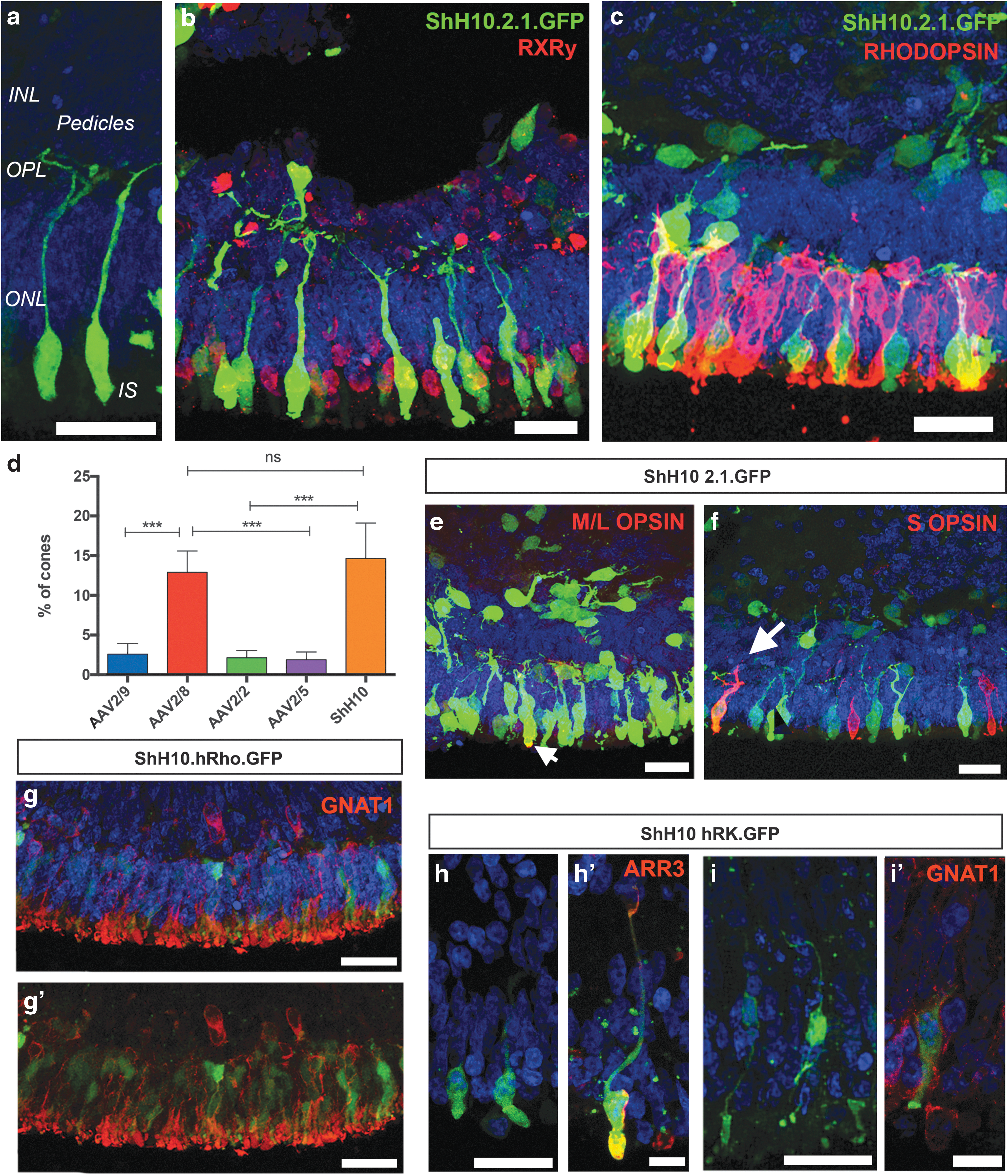
Testing promoter specificity in human ESC-derived photoreceptors.
The results presented here confirm that the PR2.1.opsin and hRhodopsin promoters mediate robust and specific expression in human cone and rod photoreceptors, respectively. The PR2.1 promoter is active predominantly in M/L-cones, but also in some blue S-cones when transduction efficiency is high. Again, the ShH10 capsid was the most efficient serotype for the transduction of human cone photoreceptors in vitro. Surprisingly, the AAV2/8 serotype, which transduced mouse ES-derived photoreceptors poorly, transduced human ESC-derived cones efficiently in vitro. Most importantly, our results demonstrate that cone photoreceptors can be transduced at early time points in culture, while GFP+ rod photoreceptors were only observed following transduction at later time points.
Discussion
Numerous studies have demonstrated successful gene supplementation to the RPE and photoreceptor cells via AAV vectors in animal models 49 and the translation of some of these findings into clinical trials has established the feasibility of AAV ocular gene therapy. 5,6,50,51 A number of parameters must be fulfilled by preclinical studies prior to starting a gene therapy clinical trial. These include establishing vector tropism, vector transduction efficiency, expression of the transgene specifically in the target cells, safety, and finally, rescue of a disease phenotype. These provide proof of concept and are usually performed in mouse and larger animal models. In the last few years, with the advent of efficient gene editing techniques, there may also a requirement for additional preclinical proof-of-concept studies involving human retinal cells. Stem cell technology has evolved rapidly in the past decade and is now able to provide human retinal organoids for studying development and disease in vitro and for the development of therapeutic approaches involving gene editing. In order to facilitate the development of such therapies we compared the efficacy of various AAV viral vectors in mouse and human stem cell–derived RPE and photoreceptor cells.
Much of our understanding about AAV tropism has been established in the mouse retina, together with larger animals including pigs and dogs. To our knowledge, this is the first time the efficiency of AAV serotypes have been compared in stem cell–derived mouse and human retinal cells. We report a marked difference in transduction efficiency, dependent upon the serotype used. Overall, the ShH10 serotype, an AAV6 variant generated by directed evolution methodology, 42 performed superiorly compared with the other AAV serotypes tested. However, some serotypes performed similarly to ShH10, dependent on the cell type; AAV2/5 and AAV2/8 were equivalent to ShH10 at transducing mouse and human RPE and human cone photoreceptors respectively. Although it is very useful to selectively target Müller glia in mice via intravitreal injections, our studies demonstrate that this serotype can also be used in vitro to target mouse and human ESC-derived photoreceptors and RPE with high efficiency. Our findings underline the importance of selecting the appropriate AAV serotype for efficient transduction of the target cells for in vitro studies.
Pluripotent stem cell–derived human retinal cells can also be used to test promoter specificity, which would otherwise be tested in nonhuman primates. Here we show that AAV vectors carrying GFP under the control of the CMV promoter drive robust expression in mouse ESC-derived photoreceptors. Interestingly, the same vectors failed to drive expression following administration to 13-week-old human ESC-derived photoreceptors, whereas transduction of 17-week-old neuroepithelia resulted in robust GFP expression in photoreceptors. This data is in agreement with recent reports demonstrating that the developmental stage of the retina influences AAV transduction efficiency in photoreceptor cells. 7 A recent study by Petit et al., (2017) suggested that outer segments are necessary for viral transduction of rod photoreceptors. 52 Indeed, our data supports these findings, as 13-week-old neuroepithelia contains photoreceptor inner segments but not outer segment–like structures, while the first rudimentary outer segment structures are observed from 16 weeks of development. 25 In contrast, transduction of cones by vectors carrying PR2.1opsin.GFP was observed even in early stages of culture. This is also consistent with the study by Petit et al. which showed that cone, but not rod photoreceptors, were efficiently transduced at early postnatal stages in mice. 52 These results show that care should be taken when assessing vectors in retinal organoids, taking into account that the developmental stage of the organoid could potentially affect the transduction efficiency and expression levels. This explains why serotypes, such as AAV2/5 and 2/9, that are highly efficient in in vivo animal studies and mature retinal explants, do not appear to transduce a large proportion of rods in the retinal organoids.
While many animal studies have been performed to evaluate the specificity and efficiency of rod- and cone-specific promoters, there is little data regarding their efficacy in human photoreceptors. Here, we tested RPE-, cone- and rod-specific promoters in human ESC-derived retinal cultures. Although the use of human ESC-derived cells is valuable for establishing viral vector tropism for AAV gene therapy in vitro as well as promoter specificity, such in vitro cultures have their limitations and should not be used as a sole method to aid the choice of AAV vectors to be used in human trials. Similar caution should be applied when using ex vivo human retinal explants. Measurements of transduction efficiency and biodistribution need to be examined with caution, as these cultures cannot fully replicate the complex environment of the eye. The concentrated administration and subsequent spread of viral particles within the confined environment of the subretinal space cannot be replicated readily in vitro. For this reason, such in vitro studies should be regarded as complementary to in vivo work using animal models for the evaluation of viral transduction. Furthermore, it is important to note that in vitro differentiation systems do not currently reach later stages of development, equivalent to the adult retina. Toxicity is another important issue that can only be assessed in animal models, as assessments of transgene toxicity in cultures are limited to the cellular level. Systemic toxicity regarding immune related reactions to viral vectors can only be studied in vivo.
In summary, we demonstrate the utility of PSC-derived retinal tissues as a test system for the development of gene therapy vectors. Furthermore, we have used this system to assess the tropism of six AAV serotypes for mouse and human photoreceptors and RPE cells and we demonstrate that PSC-derived cells can be used to test promoter specificity in human cells. A likely next step is the use of iPSC-derived retinal cells generated from patients with inherited retinopathies as in vitro models to test a variety of gene therapy strategies, including those involving gene editing.
Footnotes
Acknowledgments
This work was supported by grants from the Medical Research Council UK (MR/J004553/1), European Research Council (ERC-2012-ADG_20120314), RP Fighting Blindness (GR576), Fight for Sight (1448/1449), the Macular Vision Research Foundation, The Miller's Trust, the Special Trustees of Moorfields Eye Charity, and a generous donation by Mr. Otto van der Wyck. A.G.C is a UCL Sensory systems and Therapies Fellow; D.G. is a Singapore A star PhD student; R.A.P. is a Royal Society University Research Fellow. R.R.A is partially funded by the Department of Health's National Institute for Health Research Biomedical Research Centre at Moorfields Eye Hospital. R.A.P is part-funded by Alcon Research Institute.
Author Disclosure
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.