
Abstract
Targeted therapy produces objective responses in bladder cancer patients, although the responses can be short. Meanwhile, response rates to immune therapy are lower, but the effects are more durable. Based on these findings, it was hypothesized that urothelial carcinoma associated 1 (UCA1)-targeted therapy could synergize with programmed cell death 1 (PD-1) blockade to enhance antitumor activity. To test this hypothesis, the effects of CRISPR-Cas9 targeting of UCA1 and PD-1 were assessed in vitro and in vivo. It was found that gRNA/cas9-targeted UCA1 induced apoptosis of 5637 bladder cancer cells, whereas PD-1 gene knockout could be achieved by electroporation of gRNA/cas9 targeting PD-1, as detected by polymerase chain reaction. In 5637 cell-xenografted humanized SCID mice, stimulation with CRISPR-Cas9 systems, immune phenotypes, and cytokine expression of human dendritic cells (DCs) was detected by flow cytometry, and polymerase chain reaction, respectively. The results of these assays suggested that the gRNA/cas9 treatment upregulated expression of CD80, CD83, and CD86 and significantly increased interleukin (IL)-6, IL-12, and IL-23 and tumor necrosis factor alpha mRNA levels. Co-administration of anti-PD-1 and anti-UCA1 treatment suppressed tumor growth and markedly improved survival of 5637 xenografted mice. Additionally, the combination treatment increased interferon gamma production by T cells that subsequently enhanced the expression of Th1-associated immune-stimulating genes to reduce transcription of regulatory/suppressive immune genes and reshape the tumor microenvironment from an immunosuppressive to a stimulatory state. Finally, anti-UCA1 treatment was shown to induce interferon gamma–dependent programmed cell death ligand 1 expression within 5637 xenograft tumors in vivo. Together, these results demonstrate potent synergistic effects of a combination therapy using LncRNA UCA1-targeted therapy and immune checkpoint blockade of PD-1, thus supporting the translational potential of this combination strategy for clinical treatment of bladder cancer.
Introduction
B
To explore the possible benefits of combining PD1-blockade and UCA1-suppression for bladder cancer treatment, this study disrupted expression of PD-1 and UCA1 using the CRISPR-Cas9 genome editing tool in a bladder cancer model. The data support that therapies involving combined targeted therapy and immunotherapy could have synergistic effects that enhance treatment efficacy for bladder cancer.
Methods
gRNA plasmids
gRNA expression plasmids were constructed using the Qiagen Endofree Plasmid Kit (Qiagen, Hilden, Germany) according to the manufacturer's protocol. 11 Dual gRNA-target UCA1 exon 1, -target-UCA1 promoter (CACCGTGGATCTCTTCACGGAATG, CACCGAGGGGCAGCTTTATAGGGCT), and PD-1 (5′-ggcttctaga-3′, 5′-ggtccccaat-3′) were synthesized by Sangon Biotech Co. (Shanghai, China). The recombinant plasmids were verified by DNA sequencing (Fig. 1).

Effect of CRISPR-Cas9-mediated knockdown of urothelial carcinoma associated 1 (UCA1) expression.
Bladder cancer cell culture and transfection
The bladder cancer cell line 5637 was purchased from the Chinese Type Culture Collection of the Chinese Academy of Sciences (Shanghai, China), and 5637 cells were cultured in RPMI 1640 medium (Invitrogen, Carlsbad, CA) supplemented with 10% fetal bovine serum at 37°C in a humidified chamber containing 5% CO2. The 5637 cells were seeded in a six-well plate and transfected with gRNA/Cas9 using Lipofectamine 2000 (Invitrogen).
Preparation of primary human peripheral blood mononuclear cells and electroporation
Apheresis blood samples were collected from healthy donors. Peripheral blood mononuclear cells (PBMCs) were isolated using a Ficoll density gradient centrifugation and suspended in AIM-V medium (Gibco, Grand Island, NY).
Apoptosis
Apoptosis was detected by an Annexin V/FITC and propidium iodide (PI) binding assay according to the manufacturer's instructions (BD Biosciences, Franklin Lakes, NJ).
Western blot
Total protein was extracted with RIPA buffer containing protease inhibitors. Protein lysates were subjected to sodium dodecyl sulfate polyacrylamide gel electrophoresis and transferred to polyvinylidene difluoride membranes that were incubated overnight with primary antibodies, including anti-human MMP2 and MMP9 (Abcam, Cambridge, United Kingdom; 1:500) and Bax, Bcl-2, and PD-1 (Cell Signaling Technology, Beverly, MA; 1:500) followed by incubation with a horseradish peroxidase–conjugated secondary antibody. The blots were then detected using enhanced chemiluminescence reagents (Pierce, Rockford, IL) with a chemiluminescence imaging system (Bio-Rad, Richmond, CA).
Establishment and characterization of hu-PBL-SCID model mice
SCID mice (4–5 weeks old) weighing 18–20 g were purchased from the Center of Experimental Animals of the Academy of Military Medical Science (Beijing, China). The mice were caged individually under specific pathogen-free (SPF) conditions. All animal experiments and protocols were performed strictly in accordance with the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health. Hu-PBL-SCID mice were established, as previously described. 12 Briefly, the SCID mice were treated with cyclosporine (40 mg/kg/day) to inhibit hemocytopoiesis. With successive 4-day intraperitoneal injections, human PBMCs (1 × 107 cells/mouse) were engrafted into SCID mice to generate hu-PBL-SCID. Enzyme-linked immunosorbent assay was used to detect human immunoglobulin G (IgG) concentrations in mouse peripheral blood.
Treatment of tumor-bearing mice
The 5637 cells (2 × 106 cells/mouse) were injected subcutaneously into the humanized SCID mice. Treatments were initiated when the xenograft tumors reached a volume of ∼200 mm 3 . The tumor-bearing mice were randomized into different experimental groups: 40 μg cas9 + 40 μg empty gRNA (group A), 40 μg cas9 + 40 μg UCA1 (group B), 40 μg cas9 + 40 μg PD-1 (group C), and 40 μg cas9 + 20 μg UCA1 + 20 μg PD-1 (group D). For plasmid transfer, 80 μg plasmid in 50 μL KPBS was injected percutaneously into the mouse tissues using a 27-gauge needle. Five minutes after the plasmid injection, the tumors were pulsed with a T820 square-wave electroporator (BTX, San Diego, CA) fitted with a 0.5 cm diameter array of six-needle electrodes. Eight square-wave pulses were delivered at a frequency of 1 Hz, with a pulse length of 50 ms and 33 V (nominal field strength: 33/[0.5] = 66 V/cm). The tumor volume was calculated by measuring the diameters, which were input into the formula V = a × b 2 /2, where V is the volume and a and b are the larger and smaller diameter, respectively.
Flow cytometry of DCs
Lymph nodes or splenocytes were digested into single-cell suspensions using DNase I and Liberase TL (Roche, Basel Switzerland), re-suspended in FACS buffer, and stained with fluorescein isothiocyanate-conjugated CD80, phycoerythrin-conjugated CD83, and allophycocyanin-conjugated CD86 (BD Biosciences, San Jose, CA). The immunophenotype was analyzed with a FACS flow cytometer (BD Biosciences).
Expression of cytokines
Total RNA was isolated using TRIzol reagent (Invitrogen) according to the manufacturer's instructions. mRNA expression of human IL-12a (hIL-12a), IL-12b, IL-6, IL-23, and tumor necrosis factor alpha (TNF-α) were detected with SYBR Premix Ex Taq II (Perfect Real Time; Takara, Dalian, China) on a real-time polymerase chain reaction (PCR) system (Applied Biosystems, Foster City, CA). The expression levels of target genes were normalized to β-actin. The primer sequences are listed in Table 1.
Primers used in real-time PCR
PCR, polymerase chain reaction; IL, interleukin; TNF, tumor necrosis factor; IFN, interferon.
Evaluation of PD-L1 within tumors
Xenograft tumors were cut into small pieces, disaggregated with collagenase (0.5 mg/mL), and filtered through strainers to generate single-cell suspensions. Cells (2 × 106) were stained with APC-conjugated anti-PD-L1 (eBioscience, San Diego, CA), and PD-L1 expression was analyzed by flow cytometry. In addition, 5637 tumor cells cultured in six-well plates were treated with peritoneal lavage fluid (20%, v/v) prepared from control or anti-UCA1-treated mice for 24 h, and PD-L1 expression was then analyzed by flow cytometry. In some experiments, 5637 tumor cells treated with peritoneal lavage fluid from anti-UCA1 treated mice were cultured in the presence of isotype or anti-interferon gamma (IFN-γ) neutralizing antibody.
T7 endonuclease 1 cleavage assay and DNA sequencing
Primers for PCR are listed in Table 1. The PCR products were subjected to heteroduplex formation after denaturing 200 ng of amplified DNA at 95°C for 5 min followed by slow cooling to 35°C. The samples were treated with 5 IU T7 endonuclease 1 in 1 × NEB Buffer 2, incubated at 37°C for 30 min and then analyzed by agarose gel electrophoresis. Tanon electrophoretic software (Tanon Science & Technology Co. Ltd., Shanghai, China) was used to measure band intensities.
Statistical analysis
All analyses were performed using IBM SPSS Statistics for Windows v19.0 (IBM Corp., Armonk, NY), with significance defined as p < 0.05. Survival rates were analyzed by the Kaplan–Meier method, and differences in survival between the groups were analyzed by the log-rank test.
Results
Cas9-mediated efficient LncRNA-UCA1 knockdown in bladder cancer cells
It was previously found that levels of UCA1 expression in 5637 cells are higher than those in T24 cells. 13 Therefore, 5637 cell lines were chosen to examine the effect of CRISPR-Cas9 on UCA1. Dual gRNA-target UCA1 exon 1 (CACCGTGGATCTCTTCACGGAATG) and gRNA-target-UCA1 promoter (CACCGAGGGGCAGCTTTATAGGGCT) were designed in a previous study. 14 UCA1 expression in 5637 cells transfected with CRISPR-Cas9-targeted UCA1 was reduced by 93% compared to the control, thus indicating effective targeting of UCA1 by the gRNA. Flow cytometry showed that cells with suppression of UCA1 expression had significant apoptosis (Fig. 1B) as well as inhibited migration and invasion (Fig. 1C and D). These results suggested that UCA1 acted as an oncogenic lncRNA in bladder cancer.
Cas9-mediated PD-1 knockdown in T cells of patients
Freshly isolated PBMCs were activated for 3 days. gRNA (GGCTTCTAGAGGTCCCCAAT) was co-transfected into PBMCs separately with the Cas9 expression vector to test transfection efficiency. The selected gRNA worked effectively with Cas9 on human genomes, as shown by PCR (Fig. 2).
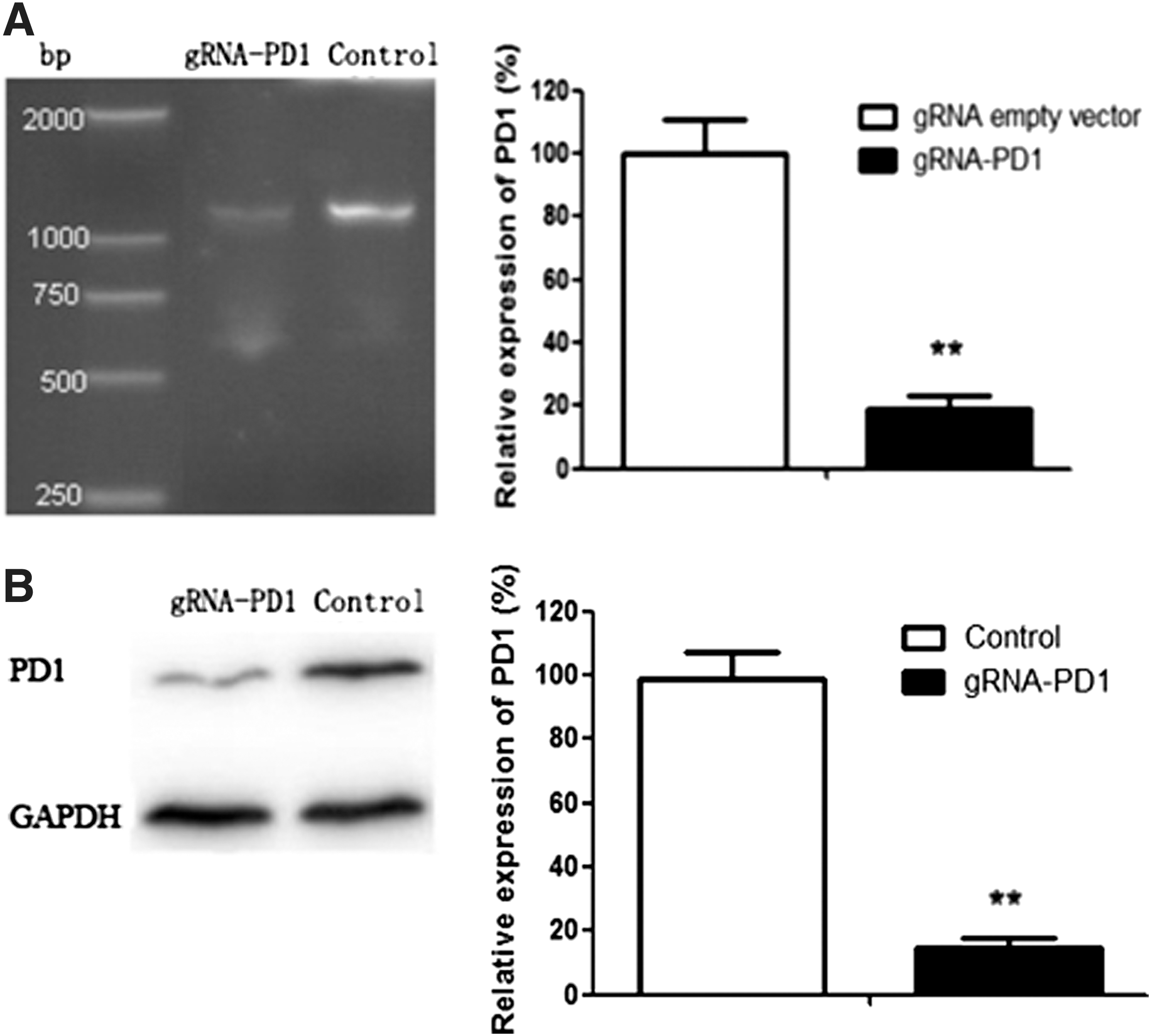
Knockdown of programmed cell death 1 (PD-1) gene expression following transfection of 5637 cells with CRISPR-Cas9. PD-1 expression was determined by
Reconstitution of hu-PBL-SCID mice and the antitumor effect of combined anti-UCA1 and anti-PD1 CRISPR-Cas9 treatment in vivo
Similar to previous reports, the serum concentrations of human IgG were variable, even among SCID mice that were reconstituted with PBL in the same experiment (Fig. 3A).
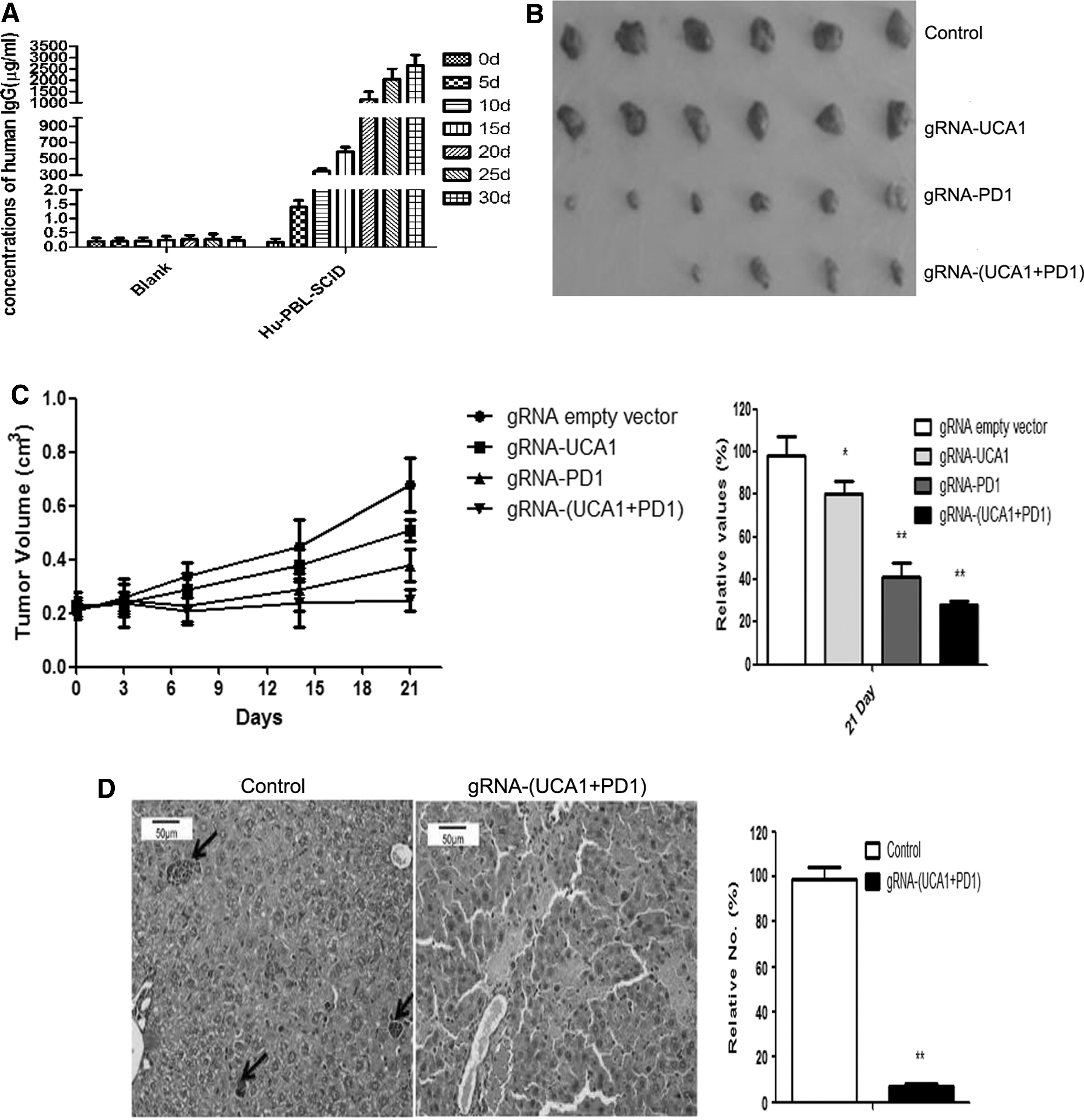
Antitumor activity of gRNA/cas9 on 5637 xenografts in a hul-mouse xenograft model. A total of 1 × 106 5637 cells were injected subcutaneously into the left flanks of mice. When the tumor volumes reached∼200 mm3, the mice were randomly divided into four groups. To observe antitumor effects, gRNA/cas9 was administered by electroporation.
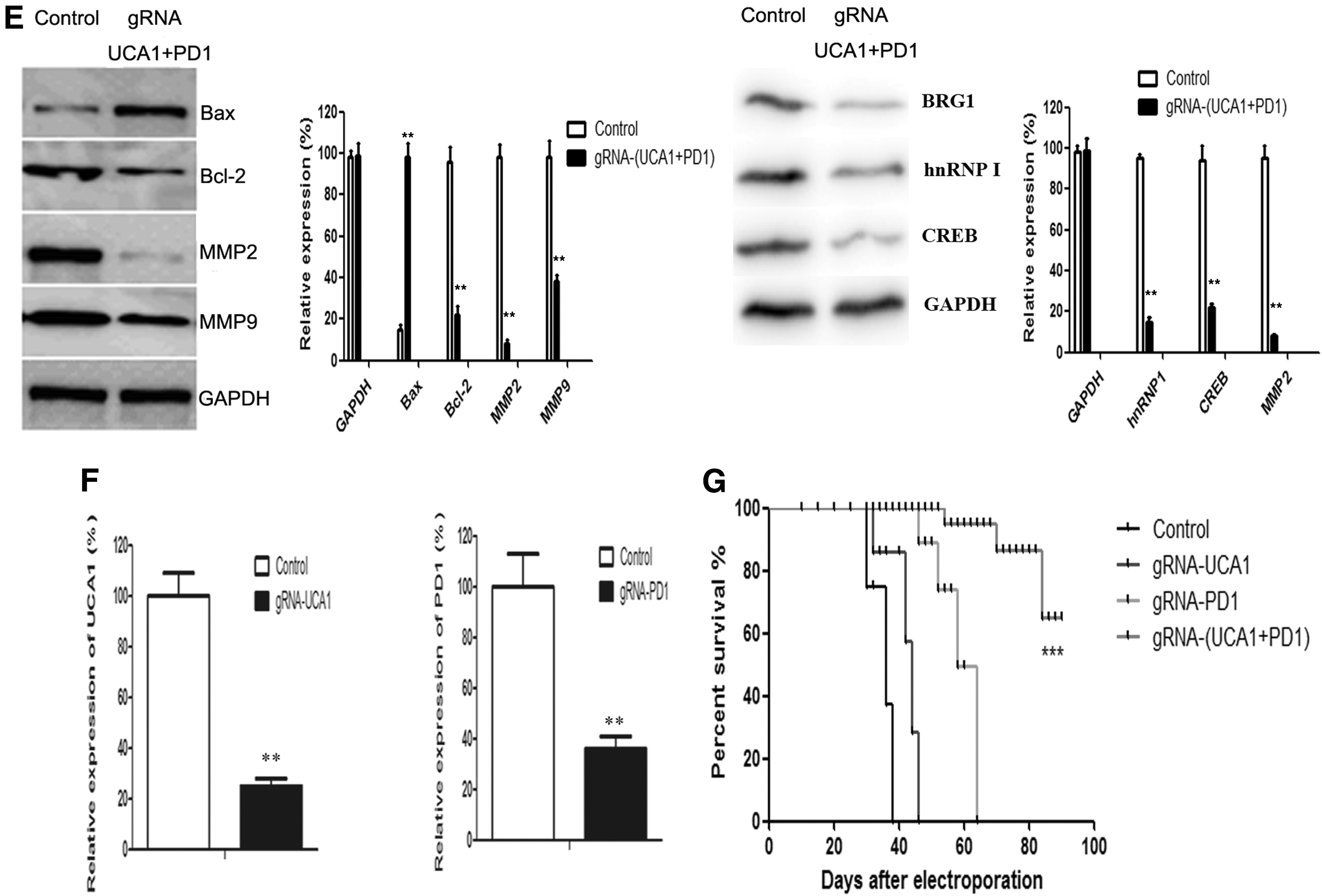
The mice were treated with electroporation of sgRNA and cas9. At day 21 after treatment, the tumor volume of the control group increased rapidly. Combined anti-UCA1 and anti-PD1 CRISPR-Cas9 exhibited a stronger antitumor effect than either treatment alone (Fig. 3B and C). Moreover, the control group showed tumor metastasis in the liver by day 21 after treatment, but the combined gRNA-(UCA1+PD1) CRISPR-Cas9-treated group did not (Fig. 3D). MMP2, MMP9, Bcl-2, BRG1, hnRNP I, and CREB expression levels were significantly decreased in tumors from the combined treatment group, whereas that of pro-apoptotic Bax was substantially increased (Fig. 3E).
Also on day 21 after treatment, UCA1 expression in tumors and PD-1 expression in DCs could still be detected in the singly treated groups, but UCA1/PD-1 expression in the combined treatment group was lower than that for the control group (Fig. 3F).
Another group of mice was used to test the antitumor efficacy of the combined treatment, as defined by prolonged overall survival of single or combined anti-(UCA1+PD-1) in hu-PBL-SCID mice. The group receiving the combination treatment showed an overall survival of >90 days after treatment (Fig. 3G).
Targeted disruption of UCA1/PD-1 sequences in vivo
A T7 endonuclease assay (T7E1) was performed to confirm the presence of gene sequence disruption. Multiple site-specific bands were detected (Fig. 4A), and the sequence of UCA1/PD-1 DNA confirmed that the intended target sites were mutated (Fig. 4B). These results suggested that the cloned UCA1/PD-1 DNA contained various mutations compared to DNA from mice that received the gRNA empty plasmid (wild type).
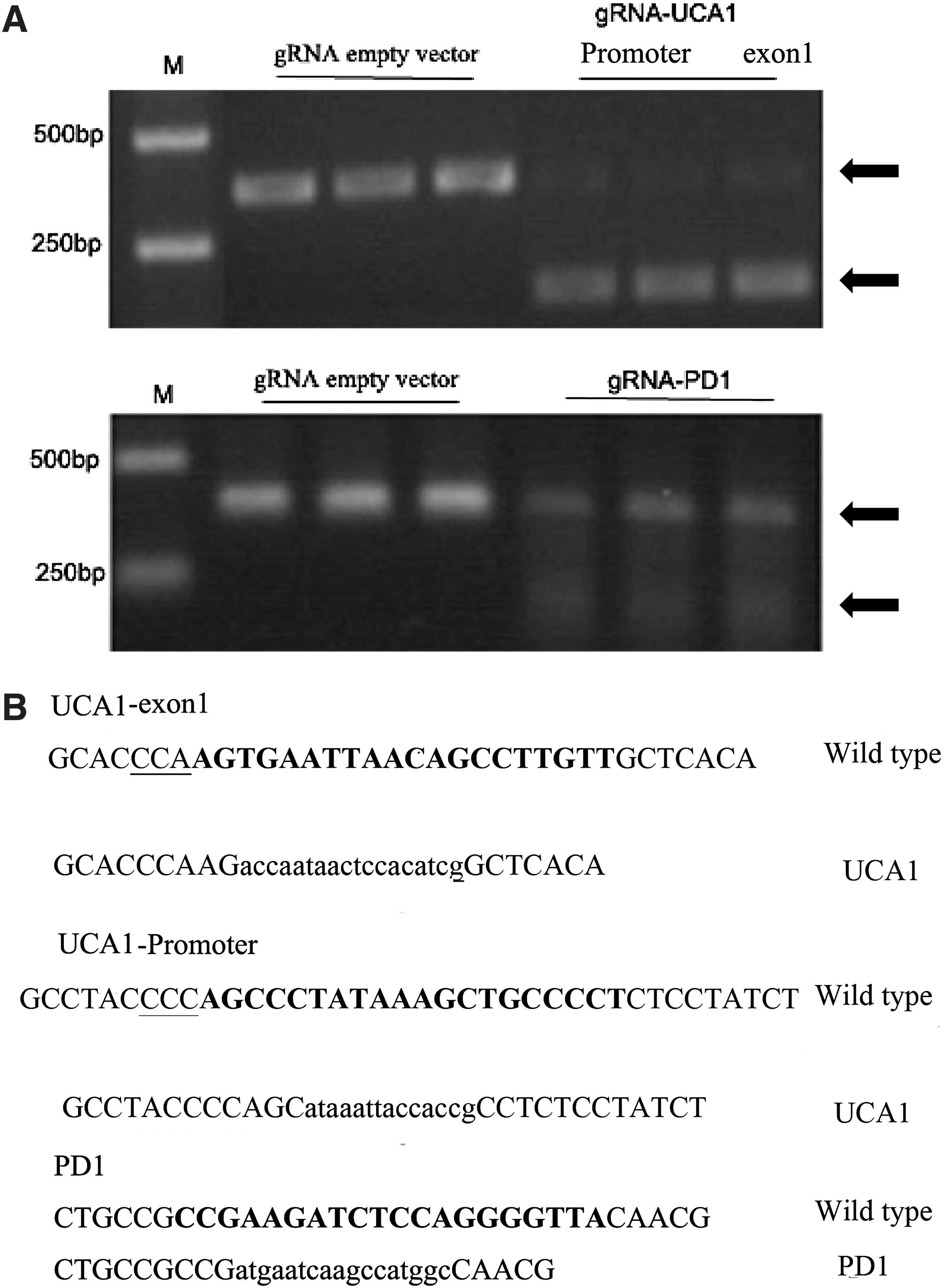
Targeted disruption of sequences in vivo.
Combined anti-UCA1 and anti-PD-1 CRISPR-Cas9 treatment synergistically induced a durable antitumor effect that is dependent on DC maturation
Previous studies demonstrated that PD-L1 expression is strongly associated with neoplastic progression and prognosis of bladder cancer. In particular, T cells and DCs are reported to express PD-1 and PD-L1, respectively. 15,16
Flow cytometry indicated that the proportion of DCs that were CD80-, CD83-, and CD86-positive among total DCs in lymph nodes was larger in the gRNA-(UCA1 + PD1) group relative to the other groups (Fig. 5A). These results demonstrated that the CRISPR-Cas9 was designed could increase the population of mature DCs.

Immune activation of human mononuclear cell-derived DCs in vivo. DCs were collected and stained with fluorescein isothiocyanate (FITC)-conjugated anti-CD80 antibody, phycoerythrin (PE)-conjugated anti-CD83 antibody, and allophycocyanin (APC)-conjugated anti-CD86 antibody, and the immune phenotypes were analyzed by FACS.
Enhanced cytokine production of DCs stimulated with gRNA/cas9
Next, IL-12, IL-23, IL-6, and TNF-α expression was detected in stimulated DCs (iDCs). It was found that DC stimulation with gRNA/Cas9-transfected cells was associated with upregulated expression of IL-6 and IL-12 but not IL-23 and TNF-α. Meanwhile, compared to cells treated with gRNA empty vector, cells transfected with PD-1 gRNA/Cas9 had enhanced expression of IL-6, IL-12, IL-23, and TNF-α (p < 0.01; Fig. 5B).
Combined anti-UCA1 and anti-PD1 treatment shaped a local immunostimulatory microenvironment in vivo
Expression profiling of immune-associated genes in tumors by real-time PCR showed that the combined anti-(UCA1 + PD-1) treatment induced a marked change in the expression of immune-associated genes (i.e., increased levels of CXCL9 and CXCL10 transcripts) and functionality (i.e., increased levels of IFN-γ and IL-12p40 transcripts; Fig. 6).
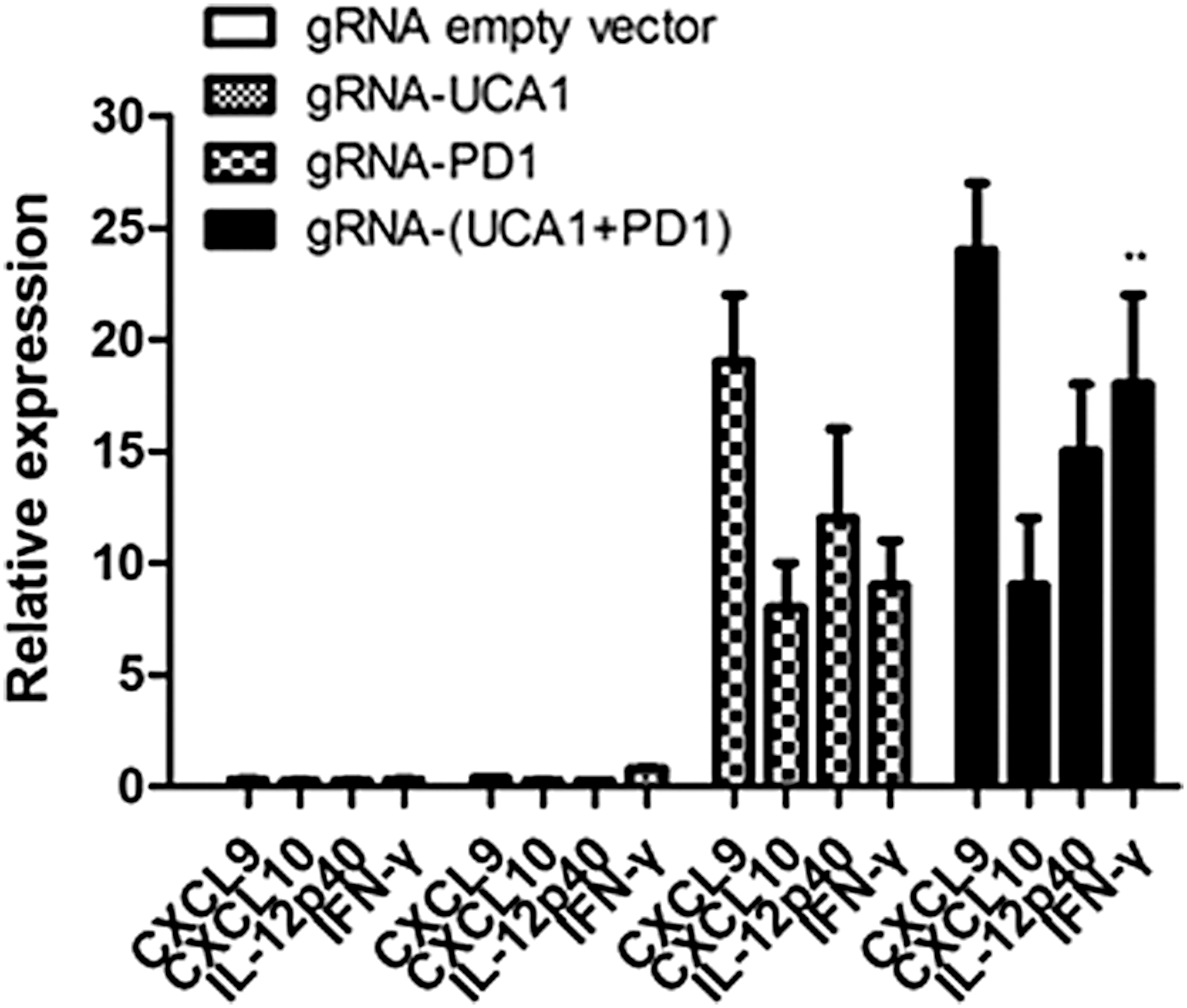
Expression profiling of immune-associated genes in tumors from treated mice. Mice were inoculated intraperitoneally (i.p.) with 1 × 106 5637 cells. RNA was extracted from the excised tumors 21 days after injection and used for analysis of immune-associated gene expression by real-time PCR. Data shown are the mean ± SD based on three independent experiments. **p < 0.01.
In vivo anti-UCA1 induction of PD-L1 expression
The presence of pre-existing, tumor antigen–specific T cells that have disabled tumor PD-L1 expression is required for successful α-PD-1 monoclonal antibody treatment. 17 It was speculated that anti-UCA1 treatment could induce tumor PD-L1 expression through IFN-γ and thus provide a target for anti-PD-1 therapy. It was further speculated that IFN-γ released from anti-UCA1 could induce PD-L1 expression. Treatment of cultured 5637 cells with lavage fluid from anti-UCA1 treated mice induced PD-L1 expression to a larger degree than did fluid from control mice, and this induction could be largely prevented by the addition of an IFN-γ-neutralizing antibody (Fig. 7A and B). These results support a key role for IFN-γ release in PD-L1 induction. This finding, together with the ability of anti-UCA1 treatment to induce PD-L1 expression in vivo, may constitute the basis for a synergistic effect of combined anti-UCA1/anti-PD-1 treatment in BC.
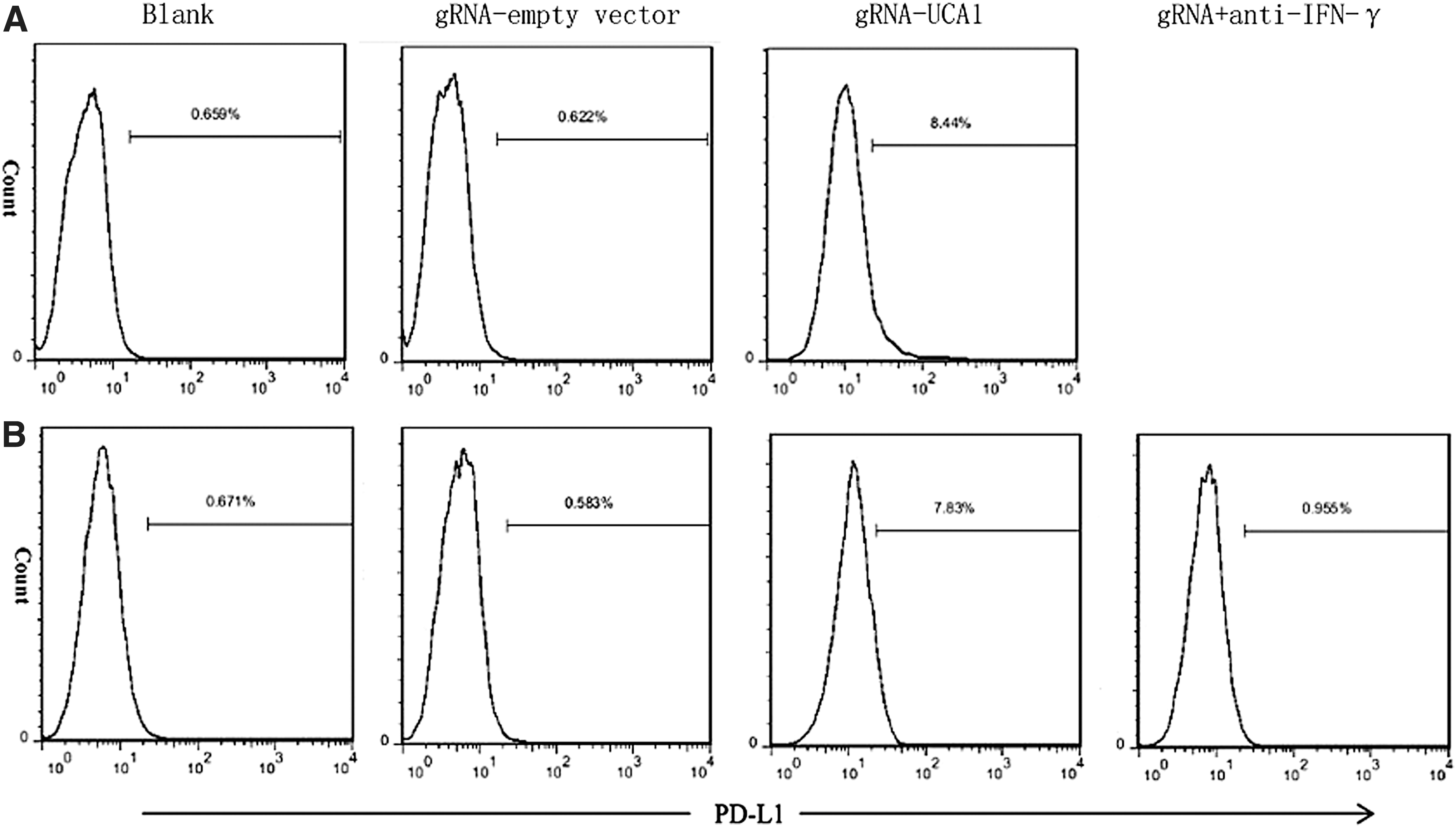
In vivo anti-UCA1 treatment induced programmed cell death ligand 1 (PD-L1) expression within 5637 tumors. Mice were inoculated i.p. with 1 × 106 5637 cells and treated with gRNA empty vector or anti-UCA1-gRNA. Tumor cells harvested from the treated mice were stained with APC-conjugated anti-PD-L1 or isotype control antibody to evaluate PD-L1 expression by flow cytometry.
Discussion
Recent advances in the understanding of antitumor immunity and immune escape have facilitated the design of new immunotherapy agents for bladder cancer treatment. 18 The clinical development of these agents increased long-term survival for a subset of patients and opened the door to combinatorial therapies that could further improve outcomes. In parallel, the discovery of oncogenic driver genes in different tumor types signified another major development in cancer treatment. Hence, there is considerable interest in testing combined treatment modalities in ongoing early stage clinical trials.
Modern immunotherapy strategies can produce durable responses but only in some patients. Accordingly, research efforts have focused on examining the effects of treatment modalities that combine targeted therapy and immunotherapy in several different histological subtypes of cancer.
Prior to the discovery of thousands of lncRNAs, the conventional view of gene regulation in biology centered on protein coding genes. Subsequent research showed that lncRNAs can act as oncogenes or tumor suppressors and can be used for diagnostic and prognostic purposes. 19 However, a major obstacle for lncRNA research is the lack of effective tools to inhibit their transcription. Many lncRNAs are localized in the nucleus and function during transcription processes rather than through the transcripts themselves, and thus the effects are difficult to knock down by RNAi. 20 Recently, a novel technology called CRISPR-Cas9 emerged that requires only changes in the sequence of the guide RNA (gRNA) that is subcloned into the specific CRISPR-Cas9 system. 21 –24 In the human genome, the CRISPR-Cas9-based system can target not only the protein-coding genes but also noncoding RNAs. 25,26 However, knockout of non-coding genes can be challenging in that a small deletion or insertion generated using the standard CRISPR-Cas system may not necessarily lead to functional loss of a given non-coding gene due to the lack of an open reading frame (particularly in polyploid human cell lines) or effects on neighboring or overlapping genes. 27 To overcome these challenges, in this study, a dual gRNA was constructed that can make two cuts simultaneously at designated sites to allow deletion of a large fragment of the UCA-1 sequence. Similar long and short ncRNAs have been employed as targets in precision oncology and indicated that CRISPR-Cas9-mediated ncRNA editing may provide an excellent way to treat cancer.
Previous characterization of UCA1 in human bladder TCC cells and BLZ-211 cells suggested that UCA1 acts as an oncofetal gene. 13 In an earlier study, specific dual gRNA that effectively inhibited UCA1 expression was designed, and it was shown that multiple gRNAs can be used to increase the targeting efficiency of lncRNA-UCA1. Thus, the CRISPR-Cas9 system can be used to modulate the expression of lncRNAs, and it can be further employed as a therapeutic approach for clinical cancer therapy. 14
This study concentrated on understanding the potential synergy between immune checkpoint blockade and UCA1-targeted therapy. Several experiments suggested that interactions between PD-1 and PD-L1 could limit, terminate, and attenuate T-cell activation, as well as induce T-cell apoptosis by reducing the production of IL-2, IFN-γ secretion, and stimulation of IL-10 production. PD-1 was inhibited by directly disrupting expression of genomic PD-1 in human T cells using gRNA/Cas9 with subsequent generation of effective gRNA for PD-1.
Recent reports revealed a role for the PD-1/PD-L1 pathway in balancing protective immunity. 28 Bladder cancer is regularly infiltrated by antigen-specific immune cells and is considered to be an ideal model for immunotherapy. 29,30 DCs have a crucial role in the induction of antigen-specific T-cell responses. Cancer patients nearly always have DC deficiencies, in both number and functional activity. 31 –33 As a classical immune checkpoint, the interaction of anti-PD1 with PD-L1 could effectively promote cytokine expression and cellular immune responses.
In testing whether combining a targeted therapy with immunotherapy has a synergistic effect that exceeds that seen by the singular agents, it was observed that combined anti-UCA1/PD1 gRNA/cas9 displayed a significantly stronger antitumor effect than either approach alone to promote long-lasting survival of mice. Targeted approaches aim to inhibit molecular pathways that are crucial for tumor growth and maintenance, whereas the goal of immunotherapy is to stimulate a host immune response that effectuates long-lived tumor destruction. Targeted therapies and cytotoxic agents also modulate immune responses, which raises the possibility that these treatment strategies might be effectively combined with immunotherapy to improve clinical outcomes. These findings support that combined anti-UCA1/PD1 CRISPR-Cas9 treatment can mount a potently durable antitumor response.
Although the mechanisms for the synergy between anti-PD-1 and anti-LncRNA require further characterization, the data suggest that anti-LncRNA UCA1 induction of PD-L1 expression may be partially responsible for the synergistic antitumor effect of the combined treatment anti-LncRNA UCA1. Several previous studies supported diverse roles for LncRNAs in immune system function. 34 –37 A number of LncRNAs have been shown to affect the magnitude of the IFN response strongly through regulation of NF-kB function, and also can potentiate or induce PD-L1 expression in tumor cells in an IFN-γ-dependent manner. The study results provide evidence that several LncRNAs could induce PD-L1 expression, which may itself mediate drug resistance on the one hand, and provide a rationale for combined cancer treatment on the other hand.
This study provides a basis for the use of a combined targeted and immune-therapy strategy for bladder cancer. The results demonstrate a potent synergistic effect of combination therapy that uses CRISPR-Cas9 to target LncRNA UCA1 and in turn block PD-1 function. These results have important translational implications.
Footnotes
Acknowledgments
This study was supported by grants from the National Natural Science Foundation of China (grant no. 81372151 to X.L.; grant no. 81602295 to S.Z.)
Author Disclosure
The authors have no conflicts of interest.