
Abstract
Sickle cell disease (SCD) is an inherited blood disorder caused by a single amino acid substitution in the β-globin chain of hemoglobin. Gene therapy is a promising therapeutic alternative, particularly in patients lacking an allogeneic bone marrow (BM) donor. One of the major challenges for an effective gene therapy approach is the design of an efficient vector that combines high-level and long-term β-globin expression with high infectivity in primary CD34+ cells. Two lentiviral vectors carrying an anti-sickling β-globin transgene (AS3) were directly compared: the Lenti/βAS3-FB, and Globe-AS3 with and without the FB insulator. The comparison was performed initially in human BM CD34+ cells derived from SCD patients in an in vitro model of erythroid differentiation. Additionally, the comparison was carried out in two in vivo models: First, an NOD SCID gamma mouse model was used to compare transduction efficiency and β-globin expression in human BM CD34+ cells after transplant. Second, a sickle mouse model was used to analyze β-globin expression produced from the vectors tested, as well as hematologic correction of the sickle phenotype. While minor differences were found in the vectors in the in vitro study (2.4-fold higher vector copy number in CD34+ cells when using Globe-AS3), no differences were noted in the overall correction of the SCD phenotype in the in vivo mouse model. This study provides a comprehensive in vitro and in vivo analysis of two globin lentiviral vectors, which is useful for determining the optimal candidate for SCD gene therapy.
Introduction
S
A definitive cure is represented by hematopoietic stem-cell transplantation (HSCT) with bone marrow (BM), mobilized peripheral blood stem cells (mPBSC), or cord blood. 11 However, allogeneic HSCT is limited by the availability of well matched donors and the immunological complications of graft rejection and graft-versus-host disease, especially for the >80% of patients who lack a human leukocyte antigen–identical sibling donor. Thus, despite advances with novel approaches to allogeneic HSCT, SCD continues to be a significant cause of morbidity and early mortality.
A promising approach to cure SCD is gene therapy using autologous hematopoietic stem cells (HSC). This approach has the potential to afford the benefits of allogeneic HSCT to SCD patients without a matched sibling donor while avoiding immunological complications. Several proof-of-principle and preclinical studies have been published showing a number of different strategies to treat SCD, including the development of lentiviral vectors (LV) carrying an “anti-sickling” beta-globin cassette, 12,13 –15 the γ-globin gene, 16 –18 and overexpression of the β-globin gene. 19 –21 While many preclinical studies have been reported, only a single case of successful gene therapy clinical trial has been reported so far for SCD. 22 The major challenge of this approach is the optimization of a vector that allows efficient gene transfer into hematopoietic stem and progenitor cells while maintaining high-level, long-term, and lineage-specific expression of the therapeutic gene. In particular, the β-globin gene has been shown to require several regulatory elements, including introns and portions of the β-globin locus control region (LCR), as well as 3′ enhancer, in order to express at sufficient levels. Nonetheless, inclusion of all of these regulatory elements in the LV backbone, while beneficial for expression of the therapeutic gene, can cause a drop in viral titer and a decrease of infectivity into primary hematopoietic cells. The main reason of this poor infectivity is believed to be the increased length of the provirus.
The authors' group has previously shown the proof of principle 12 and preclinical studies 23 using a LV carrying an anti-sickling globin gene (Lenti/βAS3-FB). The mutated β-globin was previously shown to inhibit HbS polymerization efficiently while conferring a competitive advantage over βs-globin for tetramer formation with α-globin chains. 14
Using the same βAS3-globin gene, a Globe-AS3 LV has now been developed based on the previously described Globe vector. 24 The main differences between the Lenti/βAS3-FB and the Globe-AS3 are (1) the absence in Globe of the hypersensitive site 4 (HS4) LCR sequence and the woodchuck hepatitis virus post-transcriptional regulatory element (WPRE), and (2) a shorter (528 bp) 3′ untranslated region (UTR) globin sequence and a shorter (218 bp) β-globin intron 2 in Globe-AS3. A version of Globe-AS3 including the FB insulator, also present in the Lenti/βAS3-FB, was cloned and tested (Globe-AS3-FB). The goal of this project was to compare the vectors head-to-head and analyze transduction efficiency in primary CD34+ cells, as well as β-globin expression in vitro and in vivo, with an emphasis on in vivo long-term expression and correction of hematologic parameters in a SCD mouse model.
Methods
LV production and titration
The cloning strategy to produce the Lenti/βAS3-FB was previously described in Romero et al. 12 In the lentiviral pCCL-GLOBE1 plasmid, 24 the three anti-sickling amino acids (G16D, E22A, and T87Q) were introduced by in vitro site-directed mutagenesis (QuikChange Multi Site-Directed Mutagenesis Kit; Agilent, Santa Clara, CA) in order to obtain the Globe-AS3 plasmid. To include the FB insulator in the Globe-AS3 vector, the 3′ long terminal repeat (LTR) containing the FB element was isolated by polymerase chain reaction (PCR) amplification from the Lenti/βAS3-FB using the forward primer 5′-GAAGGGCTAATTCACTCCCA-3′ and the reverse primer 5′-ATAATAAGATGACATGAACTACTACT-3′. The GLOBE-AS3 plasmid backbone was amplified with the forward primer 5′-TGGGAGTGAATTAGCCCTT-3′ and the reverse primer 5′-AGTAGTAGTTCATGTCATCTTATTAT-3′. Homology of the two fragments allowed them to be joined seamlessly using NEBuilder HiFi (New England Biolabs, Ipswich, MA).
LVs were packaged by transient transfection of 293T cells with the vector plasmid and plasmids encoding HIV GAG/POL, REV, and the VSV-G protein. Then they were processed and concentrated by tangential flow filtration, as previously described. 25 To determine the titers of the vectors, the HT-29 human colon carcinoma cell line was transduced with different dilutions of the vectors tested. About 60 h after transduction, cells were harvested, and genomic DNA was extracted using the PureLink Genomic DNA Mini Kit (Invitrogen, Waltham, MA). Droplet Digital PCR (ddPCR) was performed, as described in Urbinati et al. 23 in order to determine vector copy number (VCN), and titers of the vectors were calculated. Briefly, for VCN determination, HIV-1 psi region and human SDC4 primers and probes were used. Droplet generation was performed, as described in Hindson et al. 26 The droplet emulsion was then amplified in a conventional thermal cycler (T100 Thermal Cycler; Bio-Rad, Hercules, CA). Thermal cycling conditions consisted of 10 min at 95°C (30 s at 94°C and 1 min 60°C) for 55 cycles, 10 min at 98°C for one cycle, and 12°C hold. Afterwards, PCR acquisition and analysis of the ddPCR data was performed with the QuantaSoft software (Bio-Rad), provided with the droplet reader.
BM CD34+ isolation and LV transduction
CD34+ cells were isolated from healthy donor or SCD patients' BM aspirates using Ficoll-Hypaque gradient separation and a CD34 MicroBead Kit (Miltenyi Biotec, Bergisch Gladbach, Germany). The isolated cells were cryopreserved in liquid nitrogen with 10% dimethyl sulfoxide (Sigma–Aldrich, St Louis, MO).
After thawing at 37°C, BM CD34+ cell samples from SCD and HD were plated at 1 × 10 6 cells/mL in non-tissue culture-treated plates precoated with RetroNectin (20 μg/mL; Takara Shuzo Co., Otsu, Japan). Cells were pre-stimulated for 18–24 h in X-Vivo 15 medium (Lonza, Basel, Switzerland) containing 1 × glutamine, penicillin, and streptomycin (Gemini Bio-Products, Sacramento, CA) and cytokines at the following concentrations: 50 ng/mL human SCF (hSCF), 50 ng/mL human hFlt3 ligand (hFlt3-l), 50 ng/mL human thrombopoietin (hTPO), and 20 ng/mL human interleukin-3 (hIL-3; all cytokines from PeproTech, Rocky Hill, NJ). CD34+ cells were transduced with concentrated viral supernatants of the Lenti/βAS3-FB LV, Globe-AS3, or Globe AS3-FB vectors. Twenty-four hours after initiation of transduction, portions of the CD34+ cells were plated in methylcellulose for colony-forming unit (CFU) assay, and other portions were plated into in vitro erythroid differentiation culture following the protocol previously described 12 and/or used for NOD SCID gamma (NSG) in vivo mouse experiments.
%AS3 mRNA by ddPCR
Reverse transcription PCR to detect %AS3-globin mRNA/total β-globin transcript (%AS3) was performed, as described in Urbinati et al. 23 Briefly, isolation of RNA was done using the RNeasy Plus Mini Kit (Qiagen, Valencia, CA), followed by reverse transcription. Two different TaqMan hydrolysis probes were used to obtain the ratio of target (HBB AS3) over reference (HBB Total—all variants of endogenous and AS3 β-globin-like) mRNA transcripts for the quantification of gene expression by ddPCR. The assay sequences were: GGAGAAGTCTGCCGTTACTG (HBB AS3/Total F2), CACTAAAGGCACCGAGCACT (HBB AS3/Total R2), 5′-FAM-ACAAGGTGA-ZEN-ACGTGGATGCCGTTG-3′ Iowa Black (HBB AS3 probe), 5′-HEX-AACCTCTGG-ZEN-GTCCAAGGGTAGACCAGCAG-3′ Iowa Black (HBB Total probe).
Transplantation of transduced human BM CD34+ cells into immunodeficient mice
Human BM CD34+ cells from healthy donors were transduced with the LV vectors or mock transduced, and 7 × 10 5 cells were transplanted by retro-orbital injection into 9- to 12-week-old NOD.Cg-PrkdcscidIl2rgtm1Wjl/SzJ (NSG) mice (Jackson Laboratories, Davis, CA) previously irradiated (250 cGy) as described. 12 After 6–10 weeks, mice were euthanized, and BM was analyzed for VCN by ddPCR normalizing vector sequences to a human-specific gene (Syndecan 4 sequence). Engraftment of human cells in murine BM was analyzed by flow cytometry using allophycocyanin-conjugated anti-human CD45 versus fluorescein isothiocyanate–conjugated anti-murine CD45 (BD Biosciences, San Jose, CA). The percentage of engrafted human cells was calculated as follows: %huCD45+/(%huCD45+ + %muCD45+). Portions of the BM from engrafted mice were enriched of human CD45 cells using immunomagnetic separation (CD45 MicroBeads, human; Miltenyi Biotec). The human CD45+ cell fraction was cultured in vitro in an erythroid differentiation assay to analyze HBBAS3 mRNA expression.
In vivo experiment in SCD mouse model
The Townes (hα/hα::βA/βS, hα/hα::-383 γ-βA/-1400 γ-βS) mouse model for SCD was used for the in vivo transplant experiment (JAX stock #013071). SCD homozygous mice were euthanized between 8 and 12 weeks, and Lin– cells were isolated using a lineage cell depletion kit for murine cells (Miltenyi Biotec). Lin– cells were pre-stimulated in culture with StemSpan (Stem Cell Technologies, Vancouver, Canada), and the following cytokines were added to the medium: mSCF (100 ng/mL), hIL-11 (100 ng/mL), m-IL3 (20 ng/mL), and hFlt-3L (100 ng/mL) (Peprotech). After 24 h of pre-stimulation, cells were transduced with the individual LV at the concentrations indicated in the text. Cells were harvested 24 h later, counted, and injected intra-orbitally into lethally irradiated (1,075 cGy) C57/Bl6 mice. Between 1 and 2 × 10 6 Lin– cells per recipient mouse were injected in all experiments. A portion of the cells (2.5 × 10 5 cells) was cultured in vitro for 2 weeks to analyze the VCN by ddPCR. Monthly peripheral blood (PB) collections were performed to follow VCN by ddPCR and HbAS3 levels by high-performance liquid chromatography (HPLC) over time. Complete blood counts (CBCs) were also performed on PB samples at every time point on a GENESIS™ Hematology Analyzer (Oxford Science, Oxford, United Kingdom). Primary mice were euthanized 4–7 months after transplant, and BM was harvested for DNA analysis and CFU assay. At the time of euthanasia, spleens were collected, weighed, and photographed. Whole unfractionated BM from primary mice (30–40 × 10 6 cells) were transplanted into secondary mice (C57/Bl6 mice) after lethal irradiation (1,075 cGy). Secondary mice were euthanized 4 months post transplant and analyzed as described for primary mice. All work with mice performed at UCLA was done under protocols approved by the UCLA Institutional Animal Research Committee.
CFU progenitor assay
CFU assay for human CD34+ cells was performed as previously described in Romero et al. 12 Briefly, 100, 300, and 900 BM CD34+ cells (non-transduced or transduced) were plated per 35 mm gridded cell culture dish in duplicate using methylcellulose medium (Methocult H4435; Stem Cell Technologies). After 14 days of culture at 5% CO2, 37°C, and humidified atmosphere, the different types of hematopoietic colonies (CFU-GM, BFU-E, and CFU-GEMM) were identified and counted. CFU were then plucked for genomic DNA isolation (NucleoSpin Tissue XS; Clontech Laboratories, Inc., Mountain View, CA) for determination of VC/cell by ddPCR.
CFU assay was also performed from murine BM from the SCD Townes mouse model with minor modifications. The numbers of whole BM cells plated were: 10,000 cells, 40,000 cells, and 120,000 cells. Methylcellulose medium was used that supports murine BFU-Es to grow (Methocult SF M3436; Stem Cell Technologies).
HPLC
HPLC to quantify human hemoglobin tetramers, including HbA, HbS, HbF, and HbAS3, was performed from in vitro erythroid differentiated cells, as previously described in Urbinati et al. 23 Briefly, erythroid cells (1 × 10 6 ) were harvested on day 18 of the erythroid culture and lysed in hemolysate reagent (Helena Laboratories, Beaumont, TX) for 5–10 min at room temperature. Cell lysates were diluted 1:10 in mobile phase A and characterized by HPLC (Infinity 1260; Agilent) using a weak cation-exchange column (PolyCAT A™; PolyLC, Inc., Columbia, MD). FASC Reference Material (Trinity Biotech, Bray, Ireland) was used to assign the elution time of common hemoglobins (HbF, HbA, HbS, and HbC). Analysis and peak integration was performed using OpenLAB CDS Chemstation software. The relative percentage of HbAS3 produced for each sample was calculated based on the sum total of areas under the curve for each of the primary hemoglobin peaks, which included fetal hemoglobin, HbF; adult hemoglobin, HbA; sickle hemoglobin, HbS; as well as the anti-sickling hemoglobin, HbAS3. The same HPLC protocol was used to quantify hemoglobin tetramers from murine PB samples. As a control, a murine PB sample from a C57 mouse was used to determine elution times of murine βmaj and βmin hemoglobins.
Results
LVs
All the vectors tested are self-inactivating (SIN) LV carrying a human β-globin gene with three amino acid modifications (AS3) that confer an anti-sickling activity to the vector. 14 The β-globin gene in the Lenti/βAS3-FB LV vector, previously described in Romero et al., 12 is driven by the endogenous β-globin promoter and enhancers, with the expression regulated by a mini LCR comprised of portions of the hypersensitive sites HS2, HS3, and HS4, as shown in Fig. 1.

Schema of the proviruses. A schema of the three vectors tested is shown. ΔU3, self-inactivating (SIN) deletion in the U3 portion of the viral long terminal repeat (LTR); FB, FII-BEAD insulator; R, R region of the viral LTR; U5, U5 region of viral LTR; Y, packaging signal; RRE, Rev responsive element; cPPT, central polypurine tract; 3′-UTR, 3′ untranslated region; βp, β-globin promoter; HS-2, HS-3, HS-4, hypersensitive sites 2, 3, and 4; dWPRE, deleted woodchuck hepatitis virus post-transcriptional regulatory element; T87Q, G16D, and E22A, the three amino acid substitutions present in AS3-globin. The main differences among the vectors are indicated.
The Globe-AS3 and Globe-AS3-FB were constructed using the backbone of the Globe vector previously described in Miccio et al., 24 substituting the β-globin gene with the engineered βAS3 globin gene. The main differences with the Lenti/βAS3 vectors are: the absence of the HS4 and WPRE sequences in Globe-AS3 and Globe-AS3-FB; a shorter intron 2 (−218 bp), and shorter 3′-UTR (−528 bp) in the β-globin gene. Thus, the Globe-AS3 vector provirus is 6 3 kb in length, and the Lenti/βAS3-FB vector is 8.9 kb. Nonetheless, the sequence homology for the common shared components is 99%.
The vectors were packaged in 293T cell line, and the titers of unconcentrated vectors were tested for all the constructs in parallel on the HT-29 cell line (Supplementary Fig. S1; Supplementary Data are available online at
Transduction efficiency in BM CD34+ cells
To analyze the efficiency of transduction of Lenti/βAS3-FB and Globe-AS3 vectors in clinically relevant target cells, CD34+ cells isolated from BM aspirates of healthy donors or SCD patients were transduced with the two vectors. Transduction efficiency was examined using two assays: droplet-digital PCR (ddPCR) to detect proviral integration in the pool of target cells and ddPCR on CFUs to detect the presence of the LV in single colonies, allowing calculation of the percentage transduction in the population of CD34+ cells tested. After isolation, CD34+ cells were pre-stimulated for 24 h and then transduced with Lenti/βAS3-FB or Globe-AS3 LV at a concentration of 2 × 10 7 TU/mL (multiplicity of infection [MOI] of 40) for 18–24 h. After transduction, cells were washed, and portions of the cells were cultured in an in vitro erythroid differentiation system for 3 weeks. Cells were harvested 14 days after transduction, and ddPCR was performed to determine VCN.
In four independent experiments, shown in Fig. 2A, the average VCN was 1.2 ± 0.7 versus 2.9 ± 0.9 for Lenti/βAS3-FB versus Globe-AS3, respectively, resulting in a 2.4-fold difference among the two vectors, which was statistically significant (p < 0.01). Therefore, despite transducing the CD34+ cells with the two vectors at equal MOI, the infectivity of the Globe-AS3 vector was higher.
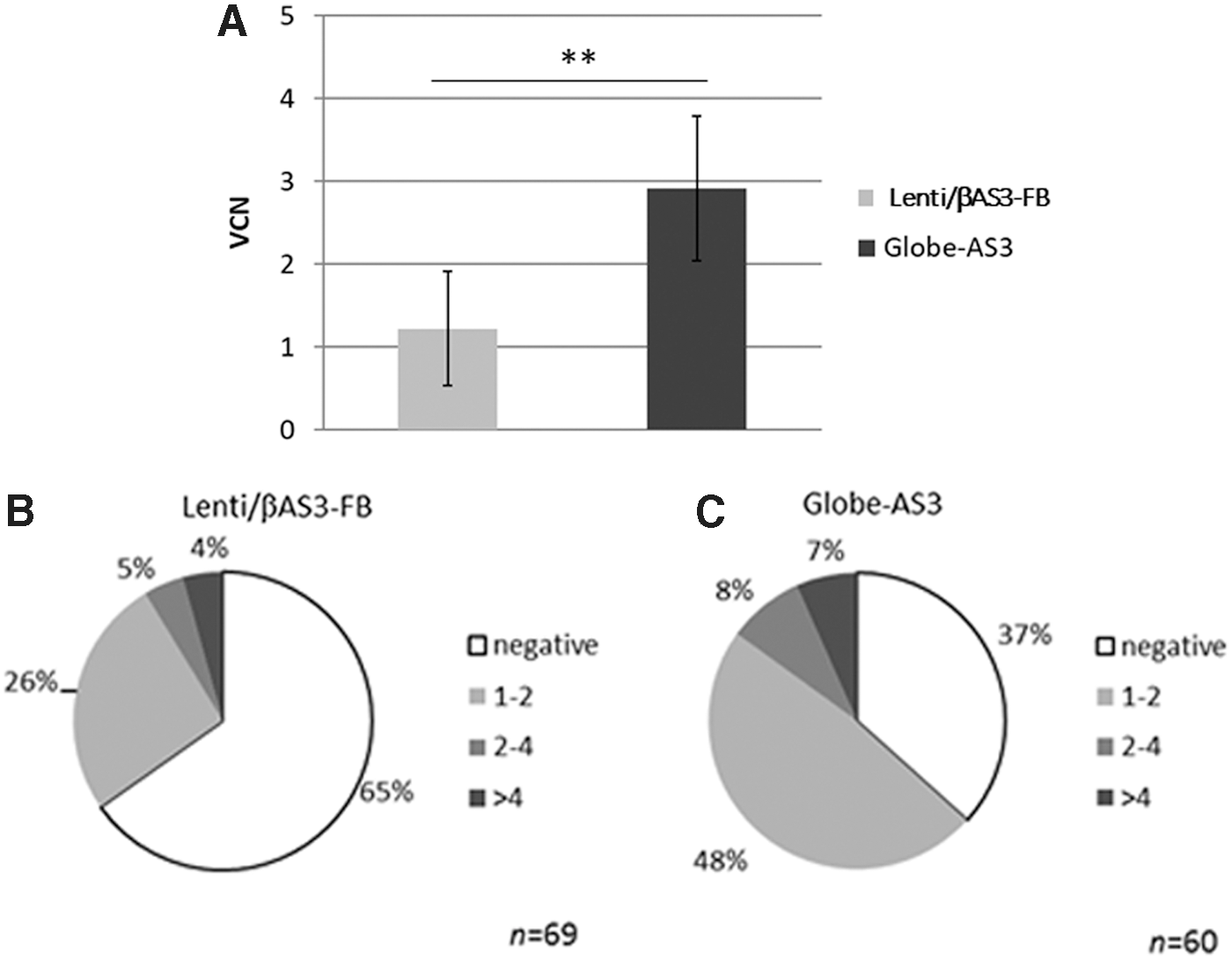
Transduction efficiency in sickle-cell disease (SCD) bone marrow (BM) CD34+ cells.
To confirm these results, analysis of transduction efficiency was also performed on single colonies. After transduction, a portion of the cells were plated in methylcellulose to allow growth of CFUs. Two weeks after plating transduced cells, colonies were scored for their lineage differentiation (BFU-E, CFU-E, CFU-GM, and CFU-GEMM) by manual count and morphologic criteria. Single colonies were picked, genomic DNA (gDNA) extracted, and ddPCR performed to determine presence or absence of the vector (%PCR + CFU) and also quantify the VCN in each transduced colony. Colonies were grouped depending on their VCN: colonies were defined as negative for the LV if the VCN was <0.3; colonies with a VCN >0.3 were considered positive and classified depending on their VCN (1–2, 2–4, or >4). The percentages of transduced colonies were higher when cells were transduced with Globe-AS3 compared to Lenti/βAS3-FB in all experiments performed (n = 4). One representative experiment is shown in Fig. 2B and C where totals of 69 and 60 colonies were analyzed for Lenti/βAS3-FB and Globe-AS3, respectively. A total of 35% of the colonies were positive for Lenti/βAS3-FB, while 63% were positive for Globe-AS3. Among the positive colonies, there was no difference in the VCN distribution among the two vectors tested: around 75% of the positive colonies contained one to two integrants, around 13% contained two to four integrants, and around 12% contained more than four integrants. Furthermore, no differences were found in the lineage differentiation of the CFUs transduced by either vector (not shown). These results indicate that at the same MOI, the Globe-AS3 is able to transduce a higher number of CD34+ cells with the same range of integrants per cell without skewing the differentiation potential of the cells when compared to the Lenti/βAS3-FB.
βAS3-globin expression in vitro
One of the main issues to consider when designing a LV for hemoglobinopathies such as SCD is the ability to achieve a level of expression of the therapeutic protein that will be sufficient to ameliorate the disease's symptoms in patients. It has been shown that the HS4 sequence in the LCR is responsible for high-level and stable globin expression. 28,29 Since the Globe-AS3 does not have the HS4 sequence, it was considered necessary to analyze any differences in expression levels from the two vectors. A preliminary analysis was performed in vitro using SCD BM CD34+ cells. To be able to detect the erythroid lineage-restricted β-globin expression, transduced SCD BM CD34+ cells were differentiated in vitro into RBC following a previously described protocol. 12,30 βAS3-globin levels were analyzed by reverse transcription PCR on RNA extracted from the cells 14 days after transduction, measured as % of β-AS3-globin RNA over % of total β-globin RNA, as described in the Methods, and by HPLC on cell lysates at the end of the differentiation culture (day 21), measured as a percentage of total globin peaks. Non-transduced cells will only present HbS and HbF peaks, while cells transduced with the LV vectors will also show a peak corresponding to βAS3-globin. Cells were transduced with different amounts of vectors, and the VCN ranged from 0.2 to 3.8. As shown in Fig. 3A, a linear relationship between VCN and βAS3-globin RNA levels was found for both vectors, as expected. A slight decrease in RNA levels was noticed for the Globe-AS3 vector compared to the Lenti/βAS3-FB vector, as shown by the elevation of the fitted line in Fig. 3A, but this difference was not statistically significant (p = 0.06).
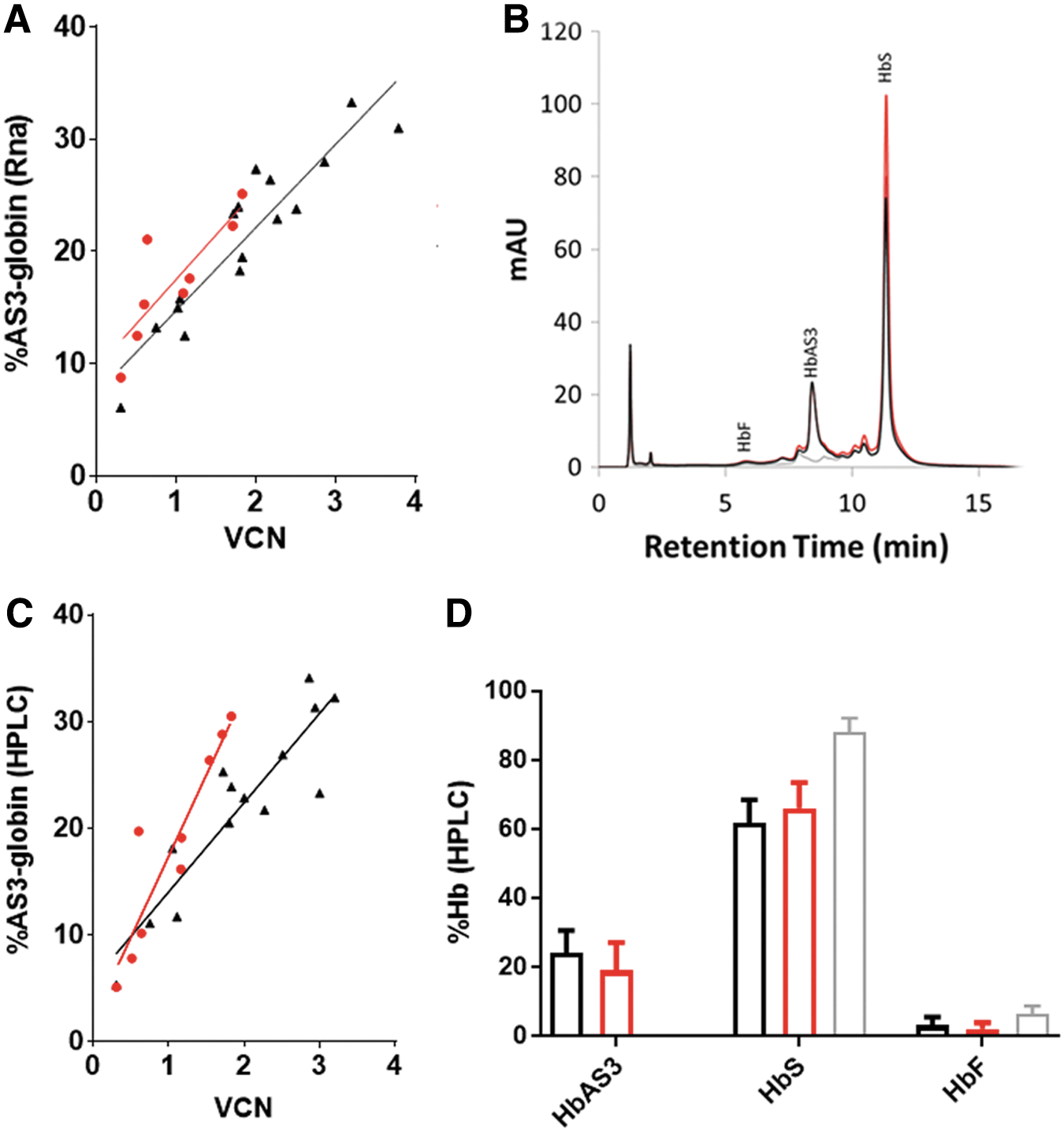
AS3-globin expression after in vitro erythroid differentiation of SCD BM CD34+ cells.
HPLC analysis to detect globin tetramers (HbF, HbS, and HbAS3) was performed, as previously described 23 on RBC lysates at the end of the in vitro erythroid differentiation (day 21). Figure 3B shows a representative experiment, where overlapping chromatograms of mock-transduced and cells transduced with the two tested vectors are shown. A new peak is present representing the βAS3-globin hemoglobin in the samples with either vector (overlapping black and orange peaks) but not in the untransduced control (gray baseline). Protein analysis (Fig. 3C) revealed a difference in βAS3-globin protein production from the two vectors tested; a decrease in slope (p = 0.01) was seen for the Globe-AS3 vector compared to the Lenti/βAS3-FB, suggesting a decrease in protein production by Globe-AS3, especially in the higher VCN range (approximately 20% HbAS3 reduction for Globe-AS3) in this short-term in vitro assay (Fig. 3C). On average, both Lenti/βAS3-FB and Globe-AS3 vectors were able to decrease the %HbS of >20% (Fig. 3D) when compared to a mock-transduced sample.
To better assess better if the difference seen here can be relevant in a clinical scenario, in vivo studies were performed in NSG mice and in a mouse model of SCD. These two mouse models allow better analysis of long-term expression of the HbAS3 protein and evaluation of any differences in ameliorating the SCD phenotype. For the in vivo studies, the Globe-AS3 vector including the FB insulator (Globe-AS3-FB) was utilized. Bridging studies were performed to show that the FB insulator does not alter the transduction efficiency and HbAS3 expression in vitro (Supplementary Fig. S2) in BM CD34+ cells. As expected, no differences (p > 0.05) were seen in VCN 2 weeks after transduction of BM CD34+ cells with Globe-AS3 and Globe-AS3-FB (Supplementary Fig. 2A), and no differences were detected (p > 0.05) in %HbAS3 produced per VCN in mature RBC after in vitro erythroid differentiation of BM CD34+ cells (Supplementary Fig. 2B).
In vivo assessment of βAS3 expression in NSG mouse model
To assess the capacity of human cells transduced by the two tested vectors (Lenti-βAS3-FB and Globe-AS3-FB) to engraft and carry out long-term expression of the therapeutic protein β-globin AS3, a humanized immunodeficient mouse model NOD.Cg-PrkdcscidIl2rgtm1Wjl/SzJ (NSG) was used. Two independent experiments were performed where healthy donor human BM CD34+ cells were transduced with the vectors at 2 × 10 7 TU/mL. Cells were harvested 24 h after transduction and injected into NSG mice (0.7 and 1.1 × 10 6 cells/mouse in experiment I and experiment II, respectively) by retro-orbital injections. A total of six mice were injected with mock-transduced BM CD34+ cells, 11 mice were injected with Lenti/βAS3-FB transduced cells (six in experiment I and five in the Experiment II), six mice were injected with Globe-AS3 vector transduced cells (experiment I), and five mice were injected with Globe-AS3-FB transduced cells (experiment II).
A portion of the cells were kept in culture for 2 weeks to analyze the average VCN of the cells before transplant. VCNs of 0.5 and 1.5 were obtained in the first experiment for LentiβAS3-FB and Globe-AS3, respectively. VCNs of 2.2 and 1.7 were obtained in vitro in the second experiment for Lenti/βAS3-FB and Globe-AS3-FB, respectively. Between 14 and 16 weeks after transplantation, mice were euthanized, and the BM was analyzed for human cell engraftment, VCN, and HbAS3 expression.
Figure 4A shows human engraftment (%hCD45) in the BM for the four groups analyzed. There were no statistically significant differences seen among the groups, as expected. gDNA was extracted from the isolated BM (a mixture of murine and human cells), and VCN was analyzed in human cells by ddPCR for the vector and an endogenous human gene (Syndecan 4; Fig. 4B). A higher VCN was seen in Globe-AS3 (M = 0.6 ± 0.1) and Globe-AS3-FB (M = 0.5 ± 0.1) transduced cells compared to the VCN in Lenti/βAS3-FB (M = 0.3 ± 0.1). These results show that the higher transduction previously shown in vitro for the Globe-AS3 (±FB) vectors is maintained in an in vivo setting, demonstrating that the higher transduced cells are indeed stem cells capable of engrafting and repopulating in the NSG xenograft model and not just progenitor cells that can be detected only in a shorter in vitro assay.
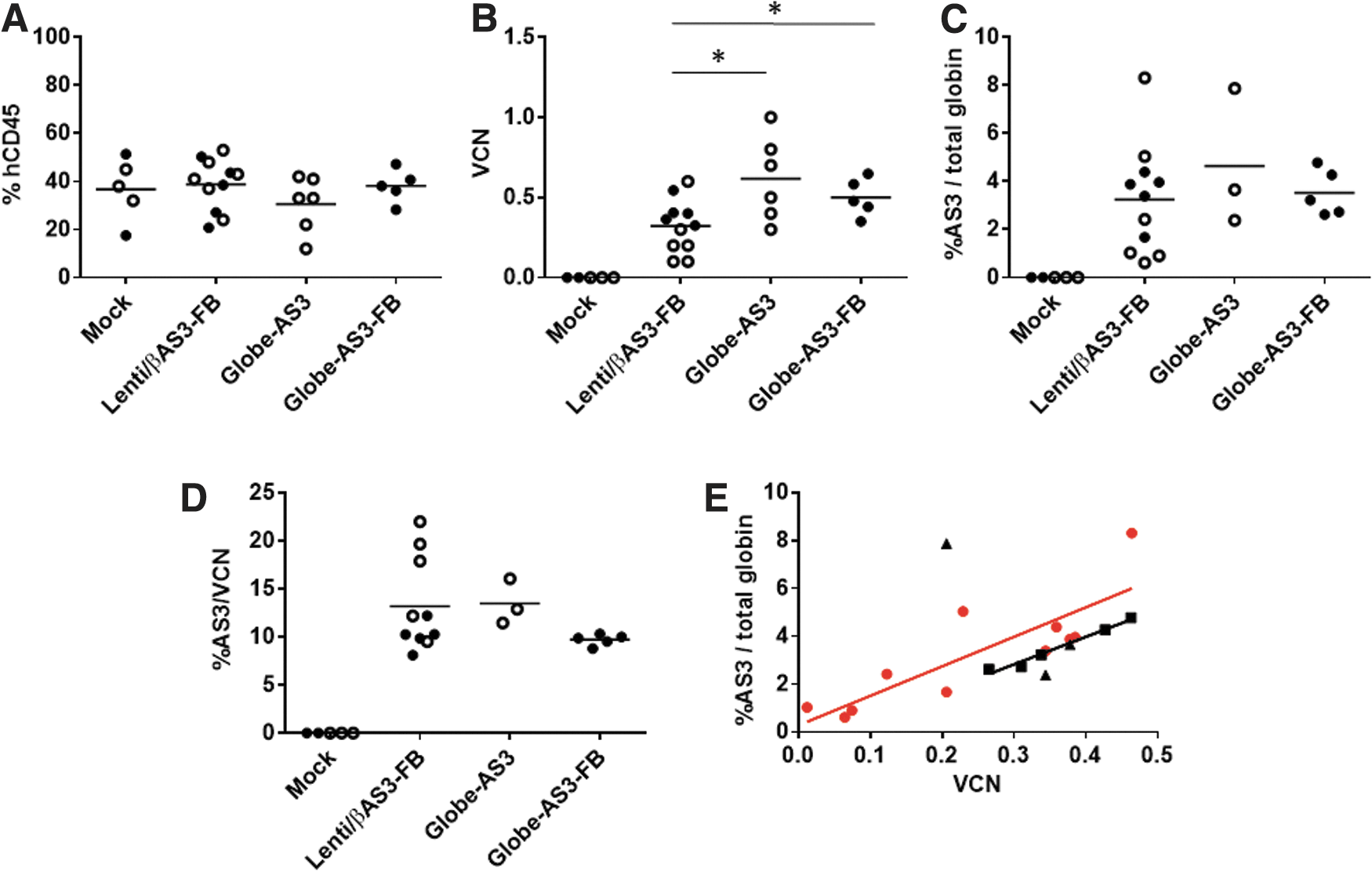
In vivo NOD SCID gamma mouse model.
Because NSG mice do not support human erythroid differentiation, 31 human CD45 cells were isolated form the mouse BM and directed to differentiate into RBC in vitro. This allowed HbAS3 expression to be assessed (Fig. 4C). Although the Globe-AS3 vectors had higher VCNs in the bulk marrow samples, no differences were seen among the three groups tested for their total AS3-globin RNA production. Also, no statistically significant differences were seen in the %AS3 RNA produced/VCN (Fig. 4D) among the vectors. When analyzing the production of AS3 RNA (percentage of βAS3 normalized for total β-globin) in relation to VCN (Fig. 4E), it is notable that the Lenti/βAS3-FB vector had a slightly higher regression line, even if the difference was not statistically significant, indicating a higher percentage of AS3-globin produced per each given VCN of Lenti/βAS3-FB compared to the Globe-AS3 vectors. The higher production of AS3 would then compensate for an average lower VCN in murine BM, resulting in overall the same net production of the therapeutic gene product. Conversely, the higher VCN of Globe-AS3 compensates for the lower expression per VCN.
In vivo phenotypic correction of SCD manifestation
To evaluate the correction of the SCD phenotype following LV transduction, an in vivo SCD mouse model was used: the “Townes mouse.” 32 A schema of the experiment is shown in Fig. 6A. BM was isolated from homozygous sickle mice and Lin–cells isolated using magnetic beads. After pre-stimulation, cells were transduced with the Lenti/βAS3-FB vector, the Globe-AS3-FB vector, or no vector (mock). Based on pilot experiments performed on specific vector batches (not shown), the amount of vector used was adjusted for each vector batch in order to have comparable transduction efficiency in the two arms in each experiment, so that a direct comparison of the efficacy of two vectors would be more appropriate. The amounts of each vector used are shown in Table 1. Cells were washed 18–24 h after transduction and injected into lethally irradiated recipient mice (C57/Bl6). Three independent experiments were performed. VCNs measured in vitro 2 weeks after transduction for the three experiments are shown in Table 1 and ranged from 0.8 to 2.9. Additionally, the percentage of transduced cells, obtained from the %CFU positive colonies, was performed for experiment III and found to be 79% transduced cells for both arms. Thus, VCN and % transduction results indicated similar transduction efficiency in murine Lin– cells for the two vectors in each experiment, allowing for a direct comparison of the two vectors. Reconstitutions of the mice were followed monthly in PB and quantified by HPLC analysis of hemoglobins (human hemoglobins, such as HbS and HbAS3, constitute the donor hemoglobins, while murine hemoglobins, such as βmaj and βmin, constitute the recipient hemoglobins). Only animals with donor hemoglobin >95% at 4 months post transplant were evaluated for hematologic parameters. This high level of human hemoglobins would prevent artifactual amelioration of hematologic parameters due to the presence of residual murine host normal RBCs that would out-compete murine donor cells expressing human HbS. Mice were euthanized 4–7 months post transplant, and BM was analyzed for VCN and RNA expression. At the time of euthanasia, secondary transplants were also performed, transplanting 30–40 × 10 6 whole murine BM cells into secondary lethally irradiated C57/bl6 mice.
In vivo analysis of AS3-globin expression in a SCD mouse model.
VCN of Lin– cells in vitro before transplant
TU, transduction units; VCN, vector copy number.
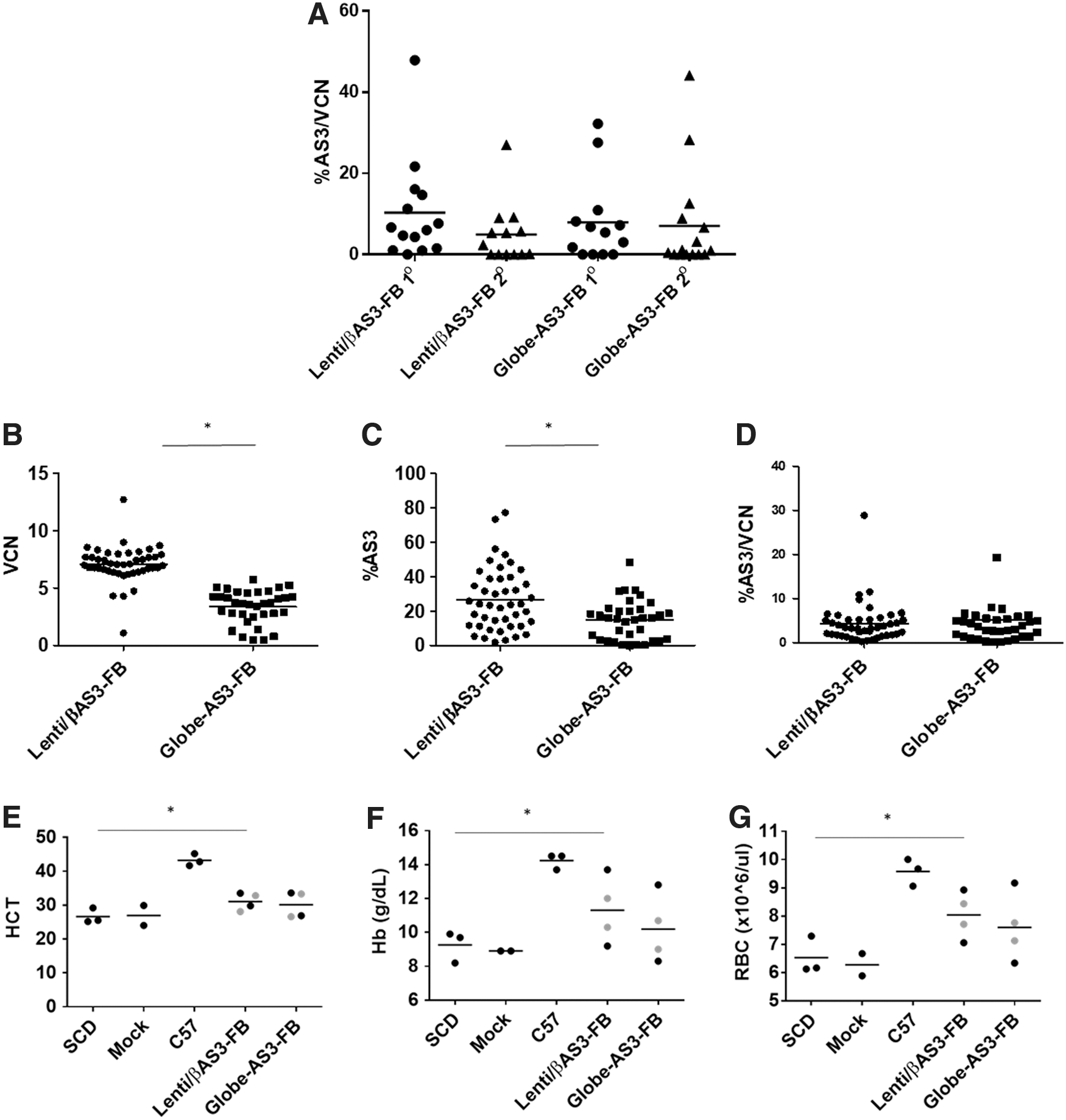
Analysis of VCN in PB at the time of euthanasia is shown in Fig. 5B. As expected, the VCN detected in vivo reflects the VCN of the cells before transplant (lower VCN in experiment II). No overall differences in VCN were seen between Lenti/βAS3-FB and Globe-AS3-FB in PB (p > 0.05) in each experiment. Figure 5C shows the VCN in PB 4–6 months after secondary transplants, and no differences were seen among the groups tested. No overall decrease in the PB VCN was detected from primary to secondary mice, indicating that murine HSCs were stably transduced with both vectors tested and were able to repopulate primary and secondary mice.

In vivo analysis in a SCD mouse model.
Expression of the AS3 transgene was analyzed by HPLC on PB lysates at the time of euthanasia in primary and secondary mice. To be able to compare the expression in the three transplants performed, the %HbAS3 was normalized to the VCN per each mouse, and results are shown in Fig. 6A. An average of 10.3 ± 3.4 and 7.9 ± 2.9 HbAS3/VC was seen in primary mice and an average of 5.0 ± 2.1 and 7.1 ± 3.3 was seen in secondary mice in Lenti/βAS3-FB and Globe-AS3-FB, respectively. No statistically significant differences were found in AS3-globin expression between the two vectors tested or between primary and secondary mice (p > 0.05), indicating the persistence of AS3-globin expression at equivalent levels by the two vectors.
Long-term and persistent transgene expression is one of the desirable features in vectors used for gene therapy. Moreover, one of the vectors (Globe-AS3-FB) is lacking a regulatory element (HS4) that has been shown to be involved in regulating the expression of the transgene. 26 Therefore, a more in-depth analysis of silencing was considered necessary. For this purpose, BM was collected from two primary mice from each group tested and plated in methylcellulose to allow growth of CFU. The VCN analyzed in the whole BM of the mice was 9.6 and 4.1 for Lenti/βAS3-FB and 3.5 and 0.6 for Globe-AS3-FB. Fourteen days after plating the cells, erythroid colonies were picked, and both DNA and RNA were extracted from the same colony. Analysis of VCN and %AS3-globin expression was performed, and results are shown in Fig. 6. A statistically significant difference was found when comparing VCN and %AS3-globin expression between the two groups (Fig. 6B and 6C). These differences reflect the difference in VCN in the whole BM of the two mice analyzed per group (average of 6.8 and 2.1 for Lenti/βAS3-FB and Globe-AS3-FB, respectively). Nonetheless, when the %AS3-globin expression was normalized per VCN (Fig. 6D), no differences were seen between the two groups, showing that the two vectors behave similarly in terms of expression at a clonal level in the mouse model used. Additionally, no evidence of silencing was seen with either vector, where a colony would be detected positive for vector integration but does not express any detectable level of AS3-globin.
Finally, hematologic parameters were analyzed for mice that had >95% donor engraftment (analyzed by HPLC in PB lysates), where no murine hemoglobins would interfere with the hematologic indexes analyzed. In these mice, the percentage of HbS varied from 47% to 96%, depending on the level of engraftment in each mouse. On average, the Lenti/βAS3-FB mice had 76% HbS and 19% HbAS3, Globe-AS3-FB mice an average of 84% of HbS and 12% of HbAS3, and mock an average of 96% HbS (no HbAS3 present). Figure 6 shows Hct, Hb levels, and RBC counts in primary and secondary mice transplanted with Lin– cells from SCD homozygous mice transduced with Lenti/βAS3-FB Globe-AS3-FB vector, or mock transduced cells (negative control). C57 control mice and homozygous SCD mice were included as negative controls.
Recipients of mock-transduced SCD BM had an average Hct of 26.6 ± 1.6. An increase to 31.0 ± 1.3% and 30.1 ± 1.9% on average was seen in Lenti/βAS3-FB and Globe-AS3-FB recipients, respectively (Fig. 6E). An increase in Hb levels was observed from 9.3 g/dL (±0.5) in recipients of mock-transduced SCD BM to 11.3 g/dL (±1.0) and 10.2 g/dL (±1.0) in Lenti/βAS3-FB- and Globe-AS3-FB-treated mice, respectively (Fig. 6F). RBC counts also increased from 6.5 × 10 6 ± 0.4 × 10 6 in recipients of the mock-transduced SCD BM to 8.0 × 10 6 ± 0.4 and 7.6 × 10 6 ± 0.6 in Lenti/βAS3-FB- and Globe-AS3-FB-treated mice (Fig. 6G). Even if a trend of higher HbAS3 and higher Hb levels were noted when the mice were transplanted with Lenti/βAS3-FB versus Globe-AS3-FB, no statistically significant differences were seen between the two vectors tested for the parameters analyzed (p > 0.05). A significant difference (p < 0.05) was seen between Lenti/βAS3-FB and SCD mice (but not between Globe-AS3-FB and SCD mice) for the same parameters analyzed. The average VCN in the mice analyzed was 3.4 and 1.3 for Lenti/βAS3-FB (ranging from 0.1 to 8.5) and Globe-AS3-FB (ranging from 0.1 to 4.9), respectively. The somewhat lesser improvement seen with the Globe-AS3-FB-treated animals compared to the Lenti/βAS3-FB mice could be due to an overall lower VCN that was found in these animals.
Discussion
Gene therapy could be a promising treatment for SCD, and significant progress has been made in this field in the past years. Several clinical trials are ongoing (reviewed in Hoban et al. 19 ), and one study has recently been published 22 where a single pediatric patient was described to have complete clinical remission after treatment with an anti-sickling β-globin LV. One of the main difficulties with this approach has been designing a LV that can be effectively packaged at a high titer with high transduction efficiency into human HSCs while still maintaining all the regulatory elements necessary for high-level and stable β-globin expression.
Until now, several vectors have been designed and tested in several laboratories, but only a few have been directly compared. 33
This study analyzed and compared two LVs for gene therapy of SCD. The Lenti/βAS3-FB was previously described in Romero et al. 12 and Urbinati et al. 23 where first proof of principle was shown in human CD34+ cells, and then IND-enabling studies were described. The vector is currently in use in a gene therapy clinical trial at UCLA (clinicaltrials.gov no. NCT02247843). Globe-AS3 and its variant Globe-AS3-FB are derived from the Globe vector initially described in Miccio et al. 24 The original vector (Globe) is also in use in a gene therapy clinical trial for β-thalassemia (clinicaltrials.gov no. NCT02453477). For the present comparison, the three amino acid modifications present in the AS3 cassette were added into the β-globin gene cassette of the Globe LV vector.
While the two vectors are similar (99% homology between Globe-AS3 and Lenti/βAS3-FB), the Globe vector is smaller (6.3 kb vs. 8.9 kb for Lenti/AS3) due to the absence of the HS4 and WPRE sequences, and its larger deletion in intron 2 and smaller 3′-UTR. Globe had a two- to threefold higher titer after packaging than Lenti/βAS3-FB and infectivity to transduce CD34+ cells when used at equal MOI, which may be the result of its smaller genome size. 27,34 Even these modest improvements in titers and infectivity would be significant if they translated to reduced costs for production on a larger scale and increased efficacy.
The comparison among the vectors presented here was performed in vitro in patient-derived cells (SCD BM CD34+ cells) and in vivo in two models: in NSG mice using human BM CD34+ cells transduced with the vectors and in a SCD murine transplant model. 32 The first model was used mainly to detect transduction efficiency in human cells and persistence of the transgene and its expression. The second model was used to compare the hematologic correction directly provided by the two vectors.
In the in vitro erythroid differentiation model with human CD34+ cells, it was found that the Globe-AS3 vector had on average 2.4-fold higher transduction efficiency but a moderate decrease in β-globin expression compared to the Lenti/βAS3-FB vector. Higher VCN was also found in the PB of NSG mice 4 months post transplantation with BM CD34+ cells transduced with the two vectors. Nonetheless, this increase in VCN did not result in an overall increase in β-globin expression in PB in mice treated with the Globe-AS3 vector.
In the SCD mouse model, it was possible to analyze the effectiveness of the two vectors in correcting the manifestation of the disease. Overall, it was found that both vectors stably transduced murine HSCs, as shown by the similar VCN found in the PB of primary and secondary mice. Additionally, no signs of expression silencing were present with either vector when analyzing CFU expression, and no differences were seen overall in hematological parameters (HCT, Hb, or RBC count) in the PB of primary and secondary mice that were treated with either vector, indicating that the levels of β-globin-AS3 expressed from both vectors were sufficient to ameliorate the SCD phenotype in the murine transplant model used. However, it is to be noted that due to the complexity of the mouse model used, where a minimum of 95% engraftment is necessary to be able to see an effect in SCD manifestation, only a small number of animals were available for analysis. This, together with animal to animal variation, may not be optimal to detect small differences between the vectors.
Overall, this study strengthens the proof of principle of the efficacy of both vectors tested, moving forward their use in gene therapy clinical trials for SCD. Further efforts to improve β-globin vectors, focused mainly on increasing the titer, transduction efficiency in primary CD34+ cells, and globin expression levels, are still necessary to improve the efficiency of these vectors, and accomplishing this would be an important advance in the field.
Footnotes
Acknowledgments
The Flow Cytometry Core of the Eli and Edythe Broad Center of Regenerative Medicine and Stem Cell Research were essential to study performance. We thank the donors with SCD who provided BM samples for these studies. This work was supported by a Sponsored Research Agreement from Biomarin Pharmaceutical, Inc.
Author Disclosure
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.