
Abstract
Gaucher disease (GD) is an autosomal recessive lysosomal storage disorder caused by mutations in the GBA gene. Enzyme replacement treatment is the most effective therapy available for type 1 GD patients, but it is very expensive and does not improve neurologic outcomes in type 2 and 3 GD patients. This study evaluated the effectiveness of an adeno-associated virus 9 (AAV9) vector expressing the Gba gene delivered systemically in GD mouse models. To detect the therapeutic effects of the AAV9-mediated Gba transfer on the systemic symptoms of GD, an inducible whole-body Gba knockout mouse was developed in which tamoxifen effectively induced whole-body Gba gene deletion, and the mice displayed systemic symptoms of GD. The AAV9-CMV-Gba vector, with the expression of Gba driven by the universal CMV promoter, restored GCase activity in multiple organs and prolonged the lifespan in tamoxifen-induced GD mice after intravenous injection. Mice with brain-specific Gba deletion were also included in this study as a model of neuropathic GD (nGD) and injected intraperitoneally on postnatal day 5 with the AAV9-SYN-Gba vector; this improved the GCase activity, ameliorated the neuropathological changes and extended the mean lifespan two-fold. This study demonstrates that AAV9-mediated gene transfer is a potentially effective treatment for GD.
Introduction
G
All GD patients should receive regular monitoring, and once GD has been initiated, treatment must generally be administered for life. There are currently two specific types of treatment for GD: enzyme replacement therapy (ERT) and substrate reduction therapy (SRT). ERT aims at supplying GD patients with a recombinant GCase protein, and SRT is meant to decrease the production of GlcCer by inhibiting the glucosylceramide synthase. 3 Both treatments have proven to be effective in treating type 1 GD, but they are very expensive and do not improve neurologic outcomes in type 2 and 3 GD patients. Full engraftment with hematopoietic stem cell transplantation or bone marrow transplantation has been performed in patients with type 1 and type 3 GD. 4 –7 Some of these patients exhibited prolonged survival, but others did not show any therapeutic effects. Moreover, the lack of human leukocyte antigen–matched donors and complications from graft-versus-host disease need to be seriously considered.
Increasingly, gene therapy is thought to be a potential alternative to traditional pharmacologic approaches since it can directly and permanently correct genetic defects, avoiding the need for repeat treatments. Many attempts have been made to discover the ideal gene delivery vectors, and adeno-associated virus (AAV) and lentiviral vectors have shown great promise for effective treatments of human genetic disorders. 8 Previous research demonstrated that ex vivo gene therapy of hematopoietic stem and progenitor cells by lentiviral vectors could correct a GD phenotype in a mouse model displaying clinical GD1 symptoms in both an early and a late intervention procedure. 9 Compared with lentiviral vectors, AAVs have many advantages, especially their lower immunogenicity, better safety, and higher efficacy for in vivo gene transfer. 10 However, there has been no report about the application of AAV vectors in GD treatment.
Here we investigated the use of AAV serotype 9 (AAV9)–mediated Gba gene delivery as a therapeutic strategy to treat GD in tamoxifen-inducible whole-body Gba knockout mice and brain-specific Gba deletion mice.
Materials and Methods
Animals
The Institutional Animal Care and Use Committee of Fudan University, China approved all protocols. The ubiquitin C promoter-cre recombinase fused with the estrogen receptor mutant T2 (UBC-CreERT2) transgenic mice express a Cre-ERT2 fusion gene under the control of the UBC promoter and the CreERT2 fusion protein consists of Cre recombinase fused to a triple mutant form of the human estrogen receptor, which can gain access to the nuclear compartment only after exposure to tamoxifen and result in deletion of the loxP- flanked sequences. Homozygous Gba-floxed mouse, Gba(flox/flox), (provided by S. Karlsson, Lund University, Sweden) 11 were crossed with UBC-creERT2 mice 12 (The Jackson Laboratory) to generate mice with the genotype Gba(flox/flox);UBC-creERT2, which developed GCase deficiency throughout the whole body when treated with tamoxifen. Neuropathic GD mice were manipulated as described by Enquist et al. 11 by mating Gba(flox/flox) mice with Nestin-cre mice 13 (The Jackson Laboratory) to remove the Gba gene specifically in neurons and neuroglia cells. Primers P1 and P2 were applied for genotyping by PCR with DNA extracted from mouse tails to determine whether the loxP cassette was present or not. The primer sequence is listed in Supplementary Table S1. To detect the UBC-creERT2 or Nestin-cre transgene, we used the primers P3 and P4. The PCR conditions were as follows: 94°C for 4 min; 38 cycles of 94°C for 30 s, 58°C for 30 s, 72°C for 30 s, and finally, 72°C for 2 min. To induce Cre recombination, 8-week-old male mice with the Gba(flox/flox);UBC-creERT2 or Gba(flox/+);UBC-creERT2 genotype were i.p. injected with tamoxifen (Sigma, T5648) solution for 5 days consecutively at a concentration of 0.5 μM tamoxifen was dissolved in corn oil (Sigma, C8267) at a concentration of 20 mg/mL. The body weight was recorded daily, and the phenotypes were monitored as well.
PCR to determine Gba gene deletion
When the tamoxifen-induced Gba(flox/flox);UBC-creERT2 mice developed obvious phenotypes, they and the control mice were sacrificed. The identification of the Gba gene deletion was performed by PCR using two primers, P5 and P6, with DNA extracted from mouse tails. P5 was in Gba exon 8, while P6 was in exon 11. The primers were intentionally designed to determine whether the Gba gene was deleted by measuring the length of the PCR products. The primer sequence is listed in Supplementary Table S1. The shorter product (426 bp) indicated gene deletion, while the larger product (1225 bp) represented the normal gene.
Quantitative real-time PCR
To analyze the Gba gene expression levels, the isolation of the total RNA of different tissues and quantitative real-time PCR (qPCR) proceeded as previously described, 14 and the description was as follows: total RNA was extracted (whole brain, heart, liver, spleen, lung, and kidney of tamoxifen-induced GD mouse; and cortex, liver, and spleen of nGD as well as the respective controls) with TRIzol reagent (Thermo Fisher), according to the manufacturer's instructions. RNA was reverse-transcribed using a PrimeScript RT reagent kit with gDNA Eraser (Perfect Real Time; TaKaRa), and the integrity of the synthesized cDNA was confirmed using glyceraldehyde 3-phosphate dehydrogenase (Gapdh) as the endogenous control. qPCR was performed in an ABI 7900 real-time PCR system with SYBR Premix Ex TaqTM II (Perfect Real Time; TaKaRa). Reactions were performed in triplicate and analyzed using an ABI 7900 Sequence Detection System (Applied Biosystems). The primer sequence is listed in Supplementary Table S1. The results were analyzed using the 2−ΔΔCT method.
GCase enzyme activity assay
The tissue samples were ground in 50 mM phosphate buffer (pH 7.0) and sonicated (Bioruptor Plus, Diagenode) in a condition with 5 s “on” and 5 s “off” for 10 cycles at a high level as soon as they were isolated from mice. The GCase enzyme activity assay was performed on the homogenate samples according to the manufacturer's instructions of QuantiChromTM β-Glucosidase Assay Kit (BioAssay Systems). The OD405nm was detected in a Multiscan Spectrum (Tecan, Infinite M200). The concentration of total protein was measured with a Pierce™ BCA Protein Assay Kit (Thermo Fisher), and the relative quantity of GCase per mg protein was calculated respectively.
AAV productions
To produce whole-body and neuron-specific AAV, the Gba gene (CDS) was subcloned into pAOV-human cytomegalovirus immediate early enhancer and promoter (CMV)-Flag or pAOV-SYN-Flag plasmid to generate pAOV-CMV-Gba-Flag or pAOV-synapsin 1 (SYN)-Gba-Flag (Neuron Biotech Co., Ltd, Shanghai, China). pAOV is the AAV packaging plasmid, which carries the inserted gene and produces AAV particles with the helper plasmids. The pAOV-CMV-Flag plasmid contains the CMV promoter to drive the systemic expression of the inserted gene fused with the downstream Flag-tag. The pAOV-SYN-Flag plasmid contains the neuron-specific SYN promoter. AAV-CMV-Gba or AAV-SYN-Gba vectors were produced by transfection of AAV-293 cells with pAOV-CMV-Gba-Flag or pAOV-SYN-Gba-Flag, AAV helper plasmid (pAAV Helper) and AAV Rep/Cap expression plasmids, as well as control AAV vectors (AAV-CMV-NC or AAV-SYN-NC). Viral particles were purified by an iodixanol step-gradient ultracentrifugation method. The genomic titer was 1.5–2.5 × 1013 viral genomes (vg) per mL determined by qPCR. The titers of the AAV9 vectors used in these studies were listed in Supplementary Table S2. The protocol was as reported by Lijuan Zhu et al. 15
AAV injections
Tamoxifen-induced 4-week-old GD mice received tail vein injection of AAV-CMV-Gba or AAV-CMV-NC. A total volume of 30 μL containing 3 × 1011 vg of each AAV vector was injected per mouse. We monitored the survival condition of each mouse for the next month. When these mice were 8 weeks old, tamoxifen was used to induce Gba deletion with i.p. injection. A total of 5 × 1011 vg of AAV-SYN-Gba or AAV-SYN-NC vector with a volume of 25 μL was administered to nGD mice with i.p. injection on postnatal day 5. Injection was performed as described by Ni-Chung Lee et al. 16
Western blot analyses
Protein extracts from freshly frozen tissues (see above) were prepared using a lysis buffer (200 mM Tris-HCl [pH 7.5], 1.5 M NaCl, 10 mM EDTA, 10 mM EGTA, 25 mM sodium pyrophosphate, 10 mM β-glycerophosphate, 1 mM Na3VO4, 50 mM NaF) supplied with a protease inhibitor cocktail (Roche Diagnostics). Protein lysates were quantified with a Pierce™ BCA Protein Assay Kit (Thermo Fisher). Protein samples of 20 μg were separated on a 10% polyacrylamide gel and analyzed by Western blot using anti-Flag (Abmart, 1:2000), anti-GAPDH (Proteintech, 1:2500), anti-RIPK3 (Cell Signaling Technology, 1:300), anti-activated caspase 3 (Cell Signaling Technology, 1:300) and anti-activated PARP (Cell Signaling Technology, 1:300) antibodies. Peroxidase-conjugated mouse or rabbit immunoglobulin G (IgG; Jackson ImmunoResearch, 1:2500) were used as secondary antibodies. Western blots were developed using ImmobilonTM Western Chemiluminescent HRP Substrate (Merck Millipore), and analysis was performed with a Luminescent Image Analyzer (GE, ImageQuant LAS 4000 mini).
Histological analyses
Tissues were immersion fixed with 10% neutral buffered formalin and embedded in paraffin blocks, and 5 μm sections were prepared on Superfrost Plus slides. Brains were cut in sagittal sections. Sections from the different tissues were stained with hematoxylin and eosin (H&E) as reported by Jiajun Xiao et al. 17 Fluoro-Jade C staining proceeded according to the manufacturer's instructions (Merck Millipore). Immunohistochemistry labeling was performed as described by Vitner et al. 2 Sections were incubated with the following primary antibodies: anti-NeuN (Merck Millipore, 1:50), anti-Iba1 (Wako, 1:100), and anti-GFAP (Sigma, 1:500). Alexa Fluor® 488 conjugated goat anti-mouse or rabbit IgG (Jackson ImmunoResearch, 1:200) were used as secondary antibodies, combined with DAPI staining.
Statistical analyses
Statistical significance was assessed using Student's t-test, as reported in the figure legends. Survival curves were compared using the log-rank Mantel-Cox test. The results were significant at p values under 0.05. All statistical tests were performed using Prism software (GraphPad, version 6.0).
Results
Inducible whole-body knockout of Gba in adult mouse led to rapidly progressive disease and death within 7 days
It has been reported that the knockout of Gba caused neonatal lethality in mice. 18 To detect the effects of the AAV9-mediated therapy on the systemic phenotypes that resulted from whole-body Gba deficiency, we first developed the tamoxifen-induced Gba knockout mice, by crossing the UBC-CreERT2 mice with the homozygous Gba(flox/flox) mice, to generate mice with the genotype Gba(flox/flox);UBC-CreERT2 (Fig. 1A and B), in which the CreERT2 recombinase was driven by the UBC promoter in all organs and needed to be further activated by tamoxifen. Previous studies have shown that endogenous estrogens present in females could nonspecifically activate the CreERT2 fusion protein prior to tamoxifen administration, so when the UBC-CreERT2 mouse was used, the males were usually selected to prevent sex effects due to endogenous estrogen present in females. 19 Therefore, for the Gba(flox/flox);UBC-CreERT2 mice, only male mice were used in our research. Without tamoxifen induction, the Gba(flox/flox);UBC-CreERT2 mice did not display any phenotypes, while tamoxifen-induced Gba(flox/flox);UBC-CreERT2 mice gradually showed disease symptoms. Five days after intraperitoneal (i.p.) injection of tamoxifen, the Gba(flox/flox);UBC-CreERT2 mice showed rapid weight loss and motor dysfunction, including abnormal gait, hyperextension of the neck, and seizures, and died (Fig. 1C and D and see also Supplementary Video). The Gba(flox/+);UBC-CreERT2 mice served as the negative controls. All the Gba(flox/flox);UBC-CreERT2 mice died within 7 days after tamoxifen induction (Fig. 1E).

Inducible knock-out of Gba in adult mouse led to rapidly progressive disease and death within 7 days.
Tamoxifen induced GCase deficiency and Gaucher cells in multiple organs of Gba(flox/flox);UBC-CreERT2 mice
Multiple organs were dissected from the Gba(flox/flox);UBC-CreERT2 mice on day 5 after tamoxifen injection. PCR amplification of the genomic DNA showed that tamoxifen treatment efficiently induced the deletion of the Gba gene in the selected organs, including the brain, liver, spleen, lung, and kidney (Fig. 2A). The quantitative PCR assay showed that the expression of the functional Gba mRNA decreased in tissues of tamoxifen-induced Gba(flox/flox);UBC-CreERT2 mice (Fig. 2B). The GCase activity also decreased in the brains and livers of Gba(flox/flox);UBC-CreERT2 mice on day 5 after tamoxifen treatment, compared with the Gba(flox/+);UBC-CreERT2 mice (Fig. 2C). The Gaucher cells were found in the brain, liver, spleen and lung tissues of the tamoxifen-induced Gba(flox/flox);UBC-CreERT2 mice (Fig. 2D). These findings demonstrated that tamoxifen effectively induced whole-body Gba gene deletion in Gba(flox/flox);UBC-CreERT2 mice, which displayed pathologic characteristics of GD. We named this mouse model the tamoxifen-induced GD mouse.
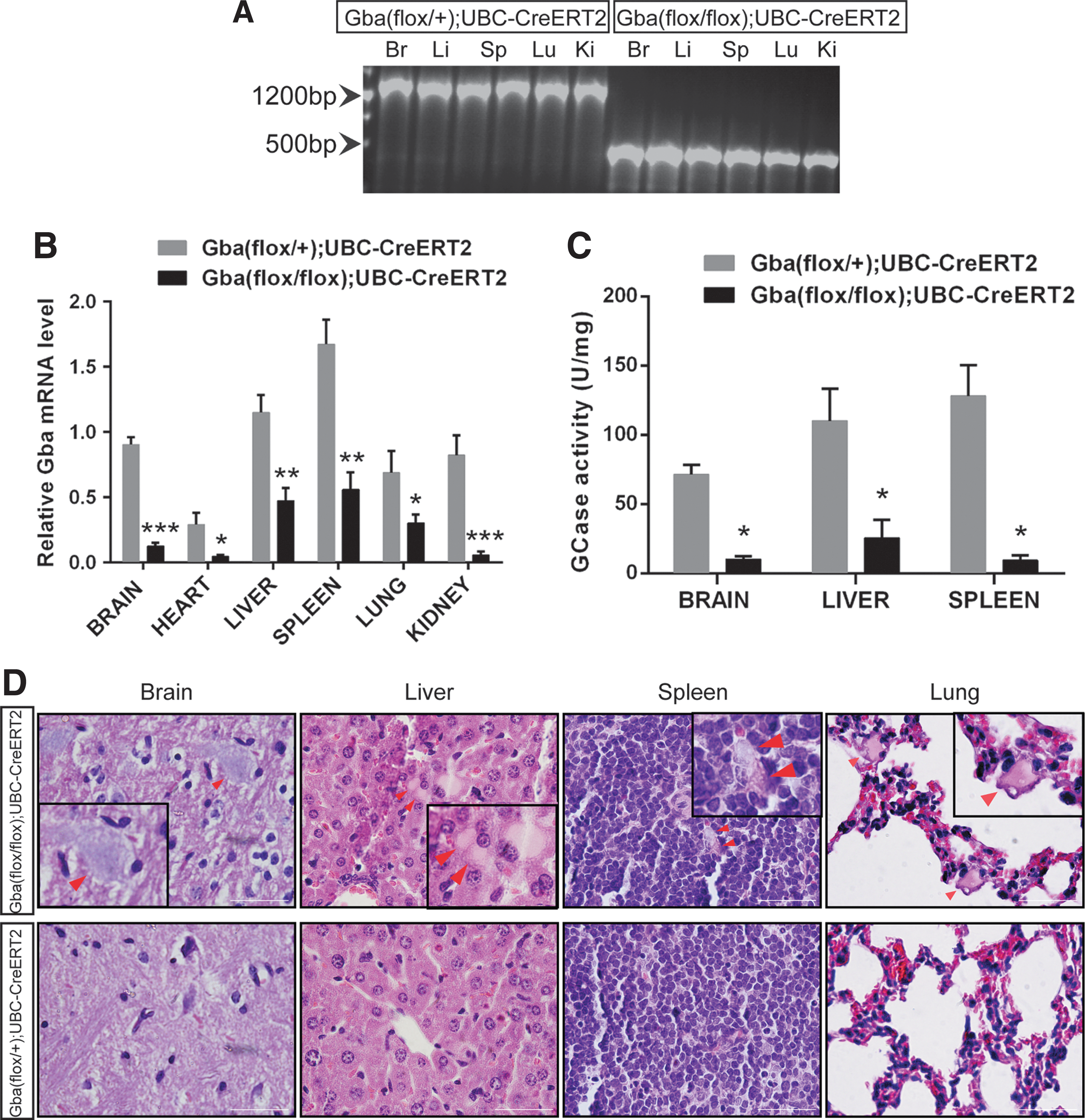
Tamoxifen-induced Gaucher disease (GD) mice developed GCase deficiency and Gaucher cells in multiple organs.
Intravenous injection of AAV9-CMV-Gba prolonged survival in tamoxifen-induced GD mice
It has been reported that the AAV9 vector can mediate systemic gene delivery and efficiently transduce several organs, such as the liver, heart, brain, and skeletal muscle. 20 AAV9 can uniquely traverse the more fortified blood–brain barrier to allow limited transduction of neurons. 21 To detect the effects of AAV-mediated gene therapy in the GD mouse model, AAV9 was selected as the delivery vector to systematically express the wild-type Gba gene. The mouse Gba CDS sequence was cloned into the AAV backbone to generate the AAV9-CMV-Gba expression plasmid, with a downstream Flag-tag sequence driven by the cytomegalovirus (CMV) promoter for systemic expression (Fig. 3A) and the Gba activity assays, in HEK293 transfected with Flag-tagged or untagged Gba expression plasmids, showed that the Flag tag had no effects on the Gba activity (Supplementary Fig. S1). The AAV9-CMV-Gba viral particles were packaged and purified, and the titer was determined by quantitative PCR. Lee et al. reported that systemic injection with 4.6 × 1011 vg of AAV vectors per mouse on postnatal day 7 could rescue multiple disease phenotypes and prolonged the lifespan significantly in aromatic l-amino acid decarboxylase deficiency mouse. 22 Considering our available titer of AAV9-CMV-Gba virus, the dosage of 3 × 1011 vg of AAV9-CMV-Gba vectors per mouse was chose in our research.
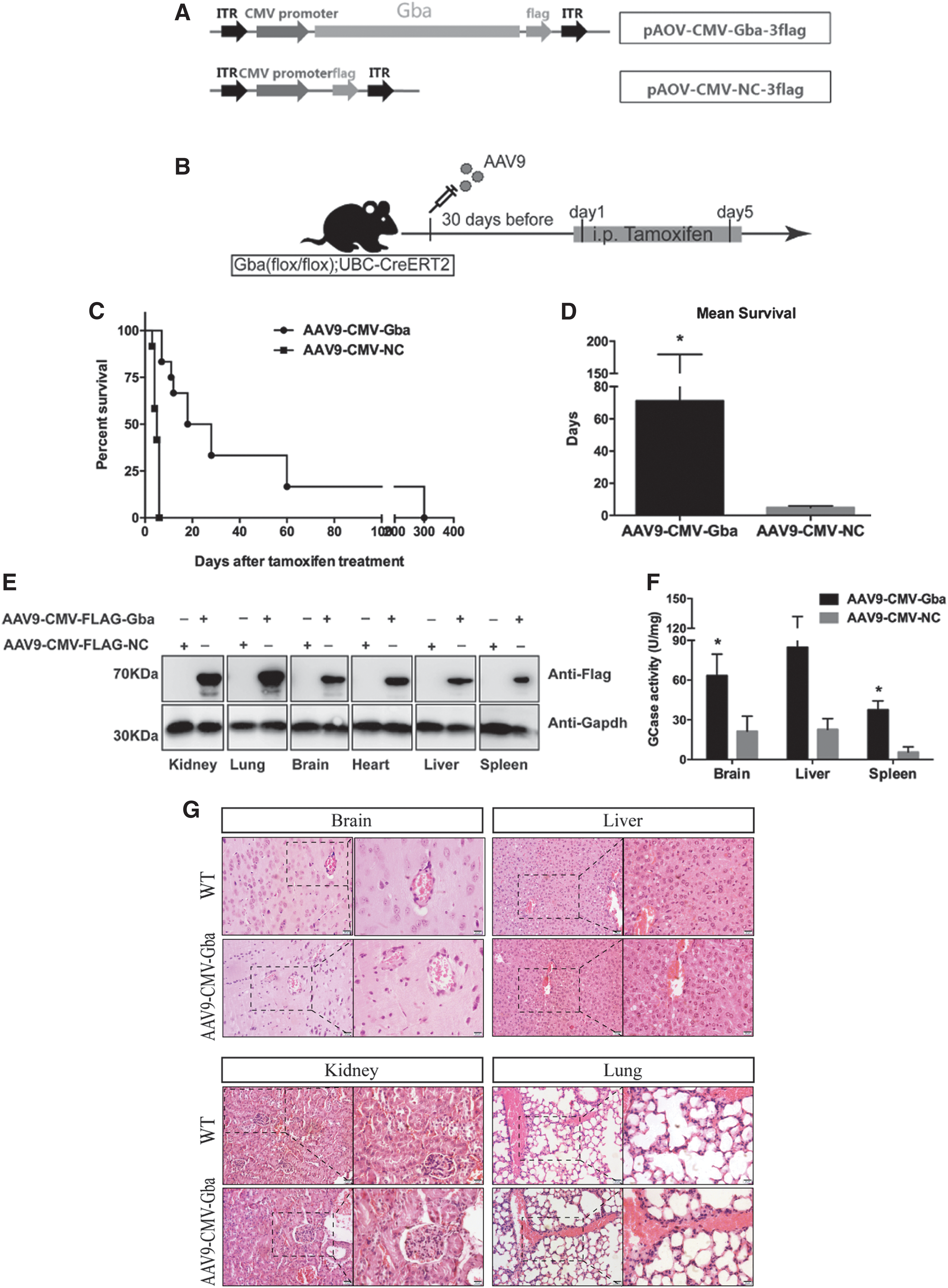
Intravenous injection of AAV9-CMV-Gba prolonged survival in tamoxifen-induced GD mice.
The AAV9 vector used here is single stranded, which is delayed in transgene expression. The AAV-delivered genes usually begin to express 1 week after infection and reach a maximum level in 2 weeks, 8 but the AAV serotype, inserted gene, and the selected promoter may affect the period of maximum transgene expression, and some previous research showed that AAV9-delivered gene expression mediated by CMV promoter peaked at 5 weeks. 23 Considering the rapid disease progression in the tamoxifen-induced GD mice and the delay in AAV-mediated transgene expression, the AAV9 vectors were injected into mice before tamoxifen induction (Fig. 3B). The purpose of the proactive injection was that the delivered Gba gene would be expressed beforehand and exert its function to prevent GCase deficiency after tamoxifen treatment. In our initial preliminary experiments, the AAV9-CMV-Gba virus was intravenously injected 15 and 30 days before tamoxifen induction, and the results showed that the survival of mice injected with AAV9-CMV-Gba at 15 days before tamoxifen induction was extended only by 1–2 days (Supplementary Fig. S2), while the injection 30 days beforehand showed great therapeutic effects, which indicated that AAV9 needed more time to restore the Gba expression or function in adult mice. Compared with the AAV9-CMV-NC-injected group, the mice injected with AAV9-CMV-Gba 30 days in advance exhibited substantially prolonged survival after tamoxifen-induced Gba deletion (Fig. 3C); the longest survival was more than 300 days. The mean survival of AAV9-CMV-Gba-injected mice was almost 14 times longer than that of AAV9-CMV-NC-injected mice (71.25 ± 31.26 vs. 4.917 ± 0.3128, *p < 0.05) (Fig. 3D). To detect the therapeutic effects of lower doses of AAV9, 3 × 1010 vg and 3 × 109 vg of AAV-CMV-Gba vectors per mouse were injected intravenously 30 days before tamoxifen induction, the median survival was prolonged by 3 days and 12 days, respectively (Supplementary Fig. S3A). Further analysis confirmed that the Gba-Flag protein was expressed (Fig. 3E) and the GCase activity increased in the selected tissues of the AAV9-CMV-Gba injected mice (Fig. 3F).
The above results indicated that the restoration of wild-type Gba genes using 3 × 1011 vg of AAV9 vectors 30 days in advance effectively increased the survival and motor performance (seeSSupplementary Video S2) in the tamoxifen-induced GD mice, which suggested that AAV9 mediated Gba delivery was a potential therapeutic method for treating GD. On 15 days after tamoxifen induction, the organs of the Gba(flox/flox);UBC-creET2 mice injected with 3 × 1011 vg of AAV-CMV-Gba were collected to detect the toxic effects. H&E staining of brain, liver, kidney, and lung of AAV-CMV-Gba-treated mice showed that the structure did not change obviously (Fig. 3G).
Intraperitoneal injection of AAV9-SYN-Gba prolonged survival and delayed the onset of disease in nGD mice
At present, there is no available effective method for treating nGD, while AAV9 has the potential to treat nGD mice due to its great central nervous system (CNS) transduction efficiency. Therefore, we further detected the application of AAV9-mediated therapy in Gba brain-specific knockout mice. The Gba(flox/flox);Nestin-Cre mice, whose Gba gene was deleted specifically in the progeny of neural and glial cell precursors, has been reported to develop progressive neurological disease that begins after an initial symptom-free period of 16 days and is fatal within 1 month after birth. 11 This GD mouse model was used as an nGD mouse in this study. To restrict the AAV9-mediated Gba expression to the nervous system, the AAV9-SYN-Gba vectors were produced in which the Gba gene was under the control of the neuron-specific Synapsin 1 (SYN) promoter (Fig. 4A). Based on the Lee's paper, 22 the dose of 5 × 1011 vg per mouse was used and the nGD mice were administered the AAV9-SYN-Gba or AAV9-SYN-NC vector by i.p. injection on postnatal day 5 (Fig. 4B). The AAV9-SYN-Gba-injected nGD mice displayed a normal increase in body weight, while the AAV9-SYN-NC-injected nGD mice had significant weight loss, which appeared on P20 and lasted until death (Fig. 4C). The survival of AAV9-SYN-Gba–treated mice was significantly extended compared with that of the AAV9-SYN-NC-injected controls, with the median survival prolonged 37 days (62 days versus 24 days; p < 0.001) (Fig. 4D) and the mean lifespan doubled (64.45 ± 9.204 versus 27.80 ± 0.7717, p < 0.01) (Fig. 4E), and there was no significantly different therapeutic effect between males and females.

Intraperitoneal injection of AAV9-SYN-Gba prolonged survival and delayed the onset of disease in neuropathic Gaucher disease (nGD) mice.
Similarly, the lower dose of AAV9-SYN-Gba vectors was also tried, and the results showed that 5 × 1010 vg and 5 × 109 vg per mouse almost showed no therapeutic effects (Supplementary Fig. S3B), which indicated that AAV9-SYN-Gba vectors needed high dose to traverse the blood–brain barrier to restore the Gba function. At 15 days after AAV injection, organs of the nGD mice injected with 5 × 1011 vg of AAV-SYN-Gba were also collected to do H&E staining, and the structure of liver, lung, and kidney of nGD that treated with AAV-SYN-Gba vectors showed normal (Fig. 4F).
AAV9-SYN-Gba restored the GCase activity and relieved the neuropathic phenotypes in the brain of nGD mice
We confirmed the efficiency of Gba delivery in AAV9-SYN-Gba-injected nGD mice at day 20, when the body weight began to decrease and obvious symptoms appeared in AAV9-SYN-NC-injected controls. Western blotting showed that the Gba-Flag protein was expressed noticeably in the cortex tissues of AAV9-SYN-Gba-injected nGD mice, while it was not detected in that of AAV9-SYN-NC-injected controls (Fig. 5A). Consistently, the enzyme activity assays also showed that GCase activity was obviously enhanced in the cortex tissues of AAV9-SYN-Gba-injected nGD mice compared with that of the controls, without increases in the liver or lung (Fig. 5B). We further measured the main neuropathological features of GD observed in nGD mice, including neuron loss, astrogliosis and microglia activation. Fluoro-Jade C staining showed that the phenotype of neuronal degeneration in nGD mice was greatly ameliorated by AAV9-SYN-Gba (Fig. 5C). The number of NeuN-marked layer V neurons, which were affected most in nGD mice, 24 increased in AAV9-SYN-Gba-treated mice and the Iba-1-positive microglia cells decreased consistently in cortex layer V (Fig. 5D). In addition, the number of glial fibrillary acidic protein (GFAP)-positive astrocytes decreased (Fig. 5D). The results suggested that neuron loss, astrogliosis and microglial proliferation in nGD mice were weakened by AAV9-SYN-Gba. It has been reported that necroptosis appeared on the symptom stage in nGD mice, and receptor interacting serine/threonine kinase 3 (RIPK3) acted as a marker in the pathway. 25 The Western blotting results also showed that the expression of RIPK3 was decreased by AAV9-SYN-Gba, without any effects on the apoptosis markers PARP and cysteine-aspartic acid protease 3 (Fig. 5E).
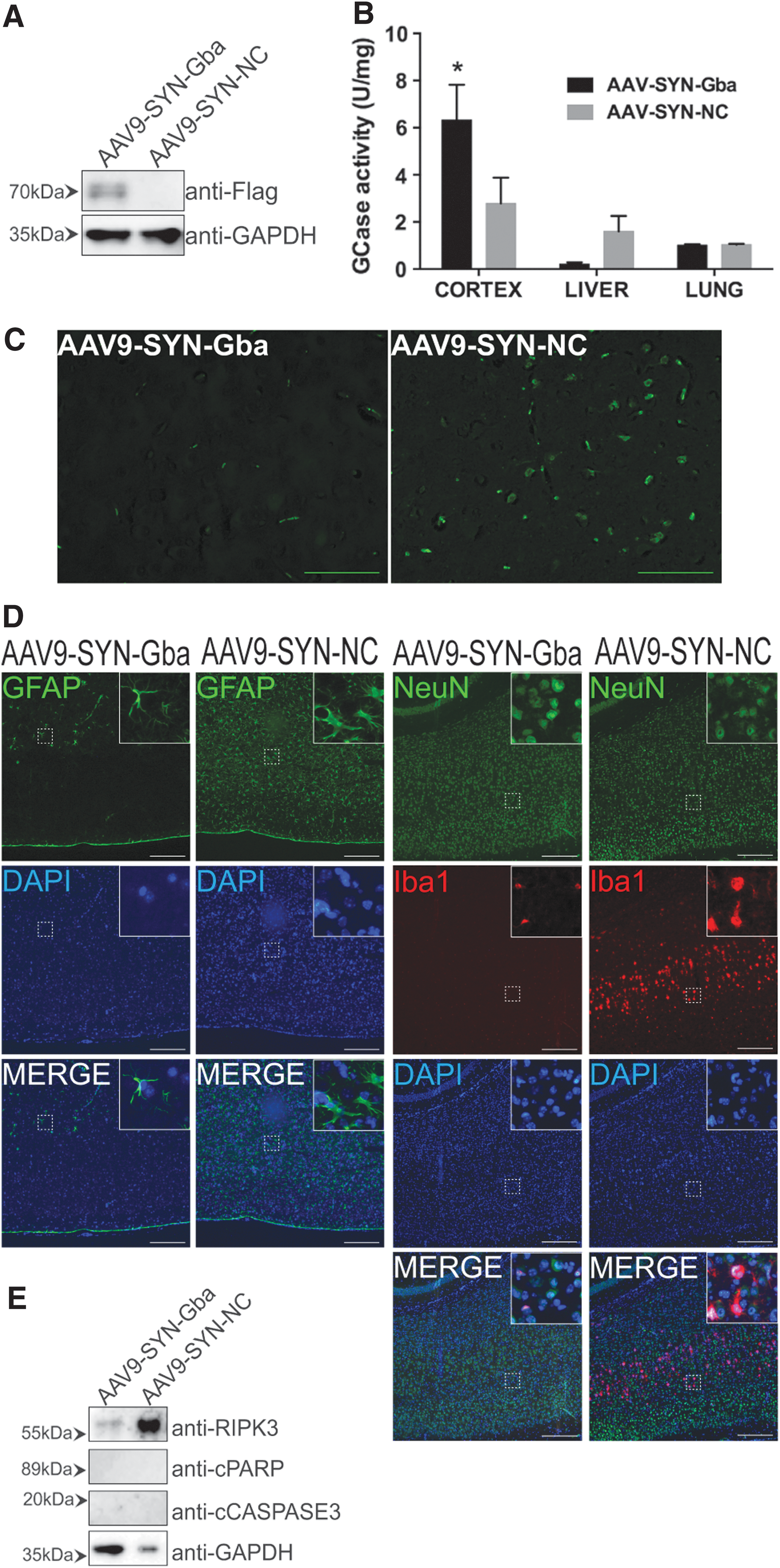
Intraperitoneal injection of AAV9-SYN-Gba restored the GCase activity and relieved the neuropathic phenotypes in the brain of nGD mice.
Discussion
GD is a common lysosomal storage disease in humans. It is an autosomal recessive genetic disease caused by mutations in the gene (GBA) coding for glucocerebrosidase (GCase), which lead to the accumulation of glucosylceramide (GlcCer) in lysosomes. Now, ERT, which supplies GCase to Gaucher cells, has been shown to be the most effective method for treating most type 1 GD patients. SRT which reduces cellular GlcCer by decreasing its production, has also been used to treat mild-to-moderate type 1 GD patients. 3 However, both have many limitations and disadvantages, such as high cost, lifelong dependence, allergic reactions, and particularly the lack of effect on nGD. Therefore, additional therapeutics are under investigation for this disease. In this study, we evaluated the effectiveness of the AAV9 vector expressing the Gba gene delivered systemically in GD mouse models.
AAV has low immunogenicity, low toxicity, and an apparent lack of pathogenicity and can invade both nondividing and dividing cells with persistent long-term transgene expression. Over 100 AAV serotypes have been isolated to date.
26
Currently, AAV vectors are among the most frequently used viral vectors for gene therapy and have shown clinical efficacy in treating several human diseases, such as Leber's congenital amaurosis,
27,28
hemophilia B,
29
aromatic
The existing GD mouse models can be categorized into two types: (1) the chemical-induced GD mouse models, by inhibition of GCase using an irreversible inhibitor, conduritol B-epoxide, 1 and (2) the genetically modified GD mouse. 39 The first GCase-deficient mouse was created by insertion of a neomycin resistance gene between Gba exons 9 and 10, and those mice died shortly after birth. The Gba-modified mouse, in which the exons 9–11 are flanked by loxP sites, was generated later, in 2006. 18 The type 1 GD mouse was created by using the Mx1-Cre-loxP system such that Gba deletion occurred only in hemopoietic stem cells and all progeny cells, and Gaucher cells appeared in the bone marrow, spleen, and liver, with no gross signs of CNS abnormalities. 18 The type 2 or 3 GD mouse model was generated with a Gba point mutation (L444P) that was known to cause human neuropathic GD, and this model was lethal soon after birth. 40 In addition, Nestin-Cre-Gba-loxP mice were later created to delete the Gba gene specifically in neural and glial cells; these mice showed the features of acute neuronal GD (type 2 GD) and died within a month after birth. 11
As described above, GD mice with whole-body ablation or point mutations of Gba developed very short lifespans after birth. To detect the therapeutic effects of the AAV9-Gba vectors in a model with whole-body Gba deficiency, Gba(flox/flox);UBC-CreERT2 mice were generated, in which tamoxifen treatment induced systemic Gba deficiency, and further analysis indicated that tamoxifen effectively induced systemic deletion of Gba gene. As a result, the mice showed abnormal phenotypes and died 7 days after tamoxifen injection with Gaucher-like cells appearing in the brain, liver, lung, and spleen.
Owing to the delayed expression of the AAV9-delivered transgene, the AAV9-CMV-Gba vectors were administered before the tamoxifen-induced Gba deletion, and the results showed that the proactive treatment with 3 × 1011 vg of AAV9-CMV-Gba 30 days before tamoxifen induction effectively improved GD phenotypes and extended the lifespan. The majority of AAV-delivered gene expression are lost during cell proliferation, but their expression is relatively stable in some slowly proliferating or nondividing cells, and some AAV can be integrated into the genome for stable transgene expression. 41 Some previous studies reported that AAV9 mediated gene expression was sustained for more than one year. 42 In this research, the mean survival time of AAV9-CMV-Gba-injected mice was increased by 14 times, with the longest lifespan up to 300 days. We suspected that the long lifespan of very few mice resulted from that the AAV-delivered gene had integrated into the genome, so it remained longer Gba gene expression. Our results also indicated that the fusion protein Gba-Flag was detectable in brain tissue at 120 days after AAV injection (Supplementary Fig. S4). The above results indicated that AAV9-mediated systemic delivery of the Gba gene had potential therapeutic value for the whole-body symptoms in GD.
Moreover, we selected the Gba(flox/flox);Nestin-Cre mice with brain-specific Gba deletion to detect the therapeutic effects of AAV9-mediated gene delivery in the neuropathic form of GD. Further analysis revealed that AAV9-SYN-Gba effectively restored the expression of Gba and the activity of GCase in the brain, which weakened the neurological symptoms and extended the lifespan in nGD mice.
AAV-mediated gene therapy has shown clinical efficacy in treating several human single-gene disorders. Our results indicated that AAV9-mediated Gba gene delivery effectively improved phenotypes and prolonged the lifespan in GD mouse models, and did not show obvious toxic effects, which demonstrates that AAV9-mediated gene transfer is a potentially effective treatment for GD. However, the safety and clinical strategies of AAV9-mediated GD treatment need further study in the future.
Footnotes
Acknowledgments
We sincerely thank Professor Stefan Karlsson (Institute of Laboratory Medicine and the Strategic Research Center for Stem Cell Biology and Cell Therapy, Lund University, Sweden) for providing the Gba (flox/flox) mouse. This work was supported by the National Key Research and Development Program of China (No. 2016YFC1000503), the National Natural Science Foundation of China (No. 81371269 and No. 81741078), and Commission of Science and Technology of Shanghai No. 14140902600 and No. 18411953300.
Author Disclosure
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.