
Abstract
Achromatopsia causes severely reduced visual acuity, photoaversion, and inability to discern colors due to cone photoreceptor dysfunction. In 2010, we reported on day-blindness in sheep caused by a stop-codon mutation of the ovine CNGA3 gene and began gene augmentation therapy trials in this naturally occurring large animal model of CNGA3 achromatopsia. The purpose of this study was to evaluate long-term efficacy and safety results of treatment, findings that hold great relevance for clinical trials that started recently in CNGA3 achromatopsia patients. Nine day-blind sheep were available for long-term follow up. The right eye of each sheep was treated with a single subretinal injection of an Adeno-Associated Virus Type 5 (AAV5) vector carrying either a mouse (n = 4) or a human (n = 5) CNGA3 transgene under control of the 2.1-Kb red/green opsin promoter. The efficacy of treatment was assessed periodically with photopic maze tests and electroretinographic (ERG) recordings for as long as 74 months postoperatively. Safety was assessed by repeated ophthalmic examinations and scotopic ERG recordings. The retinas of three animals that died of unrelated causes >5 years post-treatment were studied histologically and immunohistochemically using anti-hCNGA3 and anti-red/green cone opsin antibodies. Passage time and number of collisions of treated sheep in the photopic maze test were significantly lower at all follow-up examinations as compared with pretreatment values (p = 0.0025 and p < 0.001, respectively). ERG Critical Flicker Fusion Frequency and flicker amplitudes at 30 and 40 Hz showed significant improvement following treatment (p < 0.0001) throughout the study. Ophthalmic examinations and rod ERG recordings showed no abnormalities in the treated eyes. Immunohistochemistry revealed the presence of CNGA3 protein in red/green opsin-positive cells (cones) of the treated eyes. Our results show significant, long-term improvement in cone function, demonstrating a robust rescue effect up to six years following a single treatment with a viral vector that provides episomal delivery of the transgene. This unique follow-up duration confirms the safe and stable nature of AAV5 gene therapy in the ovine achromatopsia model.
Introduction
C
To date, mutations in six genes
In 2010, our group reported on a novel disorder of congenital day-blindness in sheep caused by a C → T substitution leading to the appearance of a stop codon in the ovine CNGA3 gene. 8,9 This new, naturally-occurring large animal model of CNGA3 achromatopsia was thoroughly characterized and investigated by means of visually-guided behavior testing, electroretinography (ERG), and histological, as well as immunohistological (IHC), techniques. 10
Starting in the summer of 2011, affected CNGA3-mutated sheep underwent unilateral gene augmentation therapy using Adeno-Associated Virus Type 5 (AAV5) vectors carrying either the mouse or the human CNGA3 transgene under control of the cone-specific 2.1-Kb red/green opsin promoter. 11 This treatment resulted in restoration of cone-mediated vision, demonstrated by both ERG and behavioral maze tests, that, at the time of our original report, was maintained for three years. 12
The purpose of this study is to provide further follow-up of the treated animals and to investigate the efficacy of gene augmentation therapy up to six years following a single subretinal treatment by means of functional assessment as well as to validate the efficacy of histological and IHC assays. With the advent of clinical trials in CNGA3 achromatopsia patients (NCT02935517 and NCT02610582), this long-term efficacy and safety follow-up holds great relevance for the development of treatments for this debilitating disease. Furthermore, it provides evidence that, at least in this large animal model, episomal delivery of the transgene provided by the AAV5 vectors allows for a long-term, stable effect that appears to resemble that seen in AAV vector-treated eyes of RPE65 mutant dogs that serve as a canine model for Leber's congenital amaurosis (LCA). 13 –18
Materials and Methods
Study design
Starting in the summer of 2011, nine day-blind sheep (five females and four males, aged 4–10 months), homozygous for the CNGA3 mutation, were treated unilaterally in the right eye with a subretinal injection of an AAV5 vector containing either the normal mouse (n = 4) or human (n = 5) CNGA3 transgene under control of a 2.1-Kb red/green opsin promoter. 12 The untreated left eye served as the control. For more than six years since then, efficacy of treatment was evaluated by periodic photopic maze testing and ERG recordings, and safety was assessed by scotopic ERG recordings, ophthalmic examination, and fundus photography. Age at the time of surgery, the transgene delivered, and follow-up examinations for each animal are presented in Table 1. Three animals were sacrificed due to unrelated causes and their retinas used for histology and IHC studies.
Age, treatment, and follow-up duration of nine experimental day-blind sheep treated with Adeno-Associated Virus Type 5 vectors carrying either the mouse or human CNGA3 transgene
ERG, electroretinography; VG, viral genomes; m, mouse; h, human; *, animal sacrificed and studied histologically.
£, animal was tested serologically for paratuberculosis and Visna/maedi virus.
Ethics and animal welfare
Experimental protocols were approved by the Volcani Center Institutional Animal Care and Use Committee and were conducted in accordance with principles outlined in the NIH Guide for the Care and Use of Laboratory Animals. For electrophysiological recordings, animals were premedicated with acepromazine (Vetmarket Ltd, Shoham, Israel) and pethidine (Dolantina; Kern Pharma, Barcelona, Spain), induced with propofol (Diprofol; Taro Pharmaceutical Industries Ltd, Mount Pleasant, NY) and diazepam (Ratiopharm, Ulm, Germany), and anesthetized and ventilated with isoflurane (Piramal Critical Care, Bethlehem, PA).
Visually guided behavior testing
Photopic maze navigation tests were conducted for each animal 0.5–2.0 months prior to gene-augmentation treatment (baseline test), and up to 68.5 months postoperatively (Table S1). Mean (±SE) duration of behavioral follow up was 61.4 ± 1.3 months, with each animal tested 11–15 times. The test protocol has been previously described in detail. 12,19 Briefly, animals were directed to pass through a 9-meter maze with two barrier obstacles. A group of sheep positioned at the end of the maze encouraged the test animal to pass through the maze as quickly as possible. For each animal, a test consisted of two successive trials with the barriers randomly rearranged in their right-left orientation between trials to avoid a learning effect. Passage time and number of collisions with the obstacles were recorded for each trial. A trial lasted up to 30 seconds, and a passage time of 30 seconds was ascribed to animals that failed to transit the maze. The average illumination intensity at the testing area (mean ± SD) was 3,352 ± 277 lux.
In addition, a subset of the maze tests (comprised of the last 5–6 tests for each animal) included additional trials with alternate patching of the treated and control eyes, as described previously. 20
ERG
ERG recordings were conducted 0.5–2.0 months prior to gene augmentation treatment and up to 73.6 months postoperatively. Mean duration of ERG follow-up was 62.0 ± 1.8 months, during which an average of 7.7 ± 1.1 recordings were conducted in each animal (Table 1). The number of ERG follow-ups was smaller than the number of behavioral follow-ups as the former requires general anesthesia, which became increasingly risky as the animals aged and was therefore performed more sparingly. Cone function was evaluated by full-field flash ERG as described previously. 10,12,19 Briefly, recordings were performed using a handheld mini Ganzfeld stimulator (HMsERG, Ocuscience, Henderson, NV) with a bandpass of 0.3–300 Hz. Following 10 minutes of light adaptation (30 cd/m2), responses to a series of flickers of increasing frequency (flashes presented at 10–80 Hz, with an average of 128 responses at each frequency) were recorded. These photopic flicker responses were recorded at four intensities (1, 2.5, 5, and 10 cd·s/m2). Responses of both treated and control eyes were recorded sequentially in random order.
Flicker response amplitudes were measured between peak and trough. Where no flicker response was detected, a value of zero was inserted as the amplitude. The critical flicker fusion frequency (CFFF), or the highest frequency at which the eye could resolve flicker, was determined for each of the four intensities.
Rod function was evaluated postoperatively once in each animal, at least 7 days after the last photopic ERG recording (last column of Table 1), using a previously described protocol. 20 Briefly, under dark-adapted conditions, responses to 10 dim flashes (0.01 cd·s/m2) presented at 0.5 Hz were recorded and averaged to generate a single scotopic flash response. This test was repeated every 4 min for a total of five scotopic flash responses recorded over 20 minutes of dark adaptation. Responses of both eyes were recorded simultaneously.
Histology and IHC
Eyes were enucleated and fixed in Davidson solution. From each eye, three ∼5- mm wide strips were cut along the nasotemporal plane, crossing the center of the optic nerve. The strips were dehydrated and embedded in paraplast (Leica Biosystems, Nussloch, Germany). From each strip, 4-μm sections were cut, and one section was stained with hematoxylin and eosin. For IHC, deparaffinized sections of the eyes were incubated in a decloaking chamber (Biocare Medical, Pacheco, CA) with 10 mM of citrate buffer (pH 6.0) at 125°C, blocked with phosphate-buffered saline solution containing 1% (w/v) bovine serum albumin, 0.1% (w/v) Triton X-100, and 10% (w/v) normal donkey serum, and subsequently incubated overnight at 4°C with anti-hCNGA3 antibody (goat polyclonal, 1:50; Santa Cruz Biotechnology, Inc, Dallas, TX) and anti-red/green cone opsin antibody (rabbit polyclonal, 1:100; Chemicon International, Temecula, CA). After the deparaffinized sections were washed, the corresponding secondary antibodies were applied for 1h at room temperature: rhodamine red-X-conjugated donkey anti-goat IgG and DyLight 488 donkey anti-rabbit IgG (1:250, Jackson ImmunoResearch Laboratories, West Grove, PA). Nuclei were counterstained with 4,6-diamidino-2-phenylindole containing mounting medium (Vectashield, Vector Laboratories, Burlingame, CA). To determine the specificity of the antigen–antibody reaction, corresponding negative controls were exposed to secondary antibody alone.
Data evaluation and statistical analyses
For analysis of visual behavior and ERG, results were grouped into 21 and 12 follow-up groups, respectively, according to time (months) before/after treatment as detailed in Tables S1 and S2. The 68- and 74-month follow-up groups were not included in the ERG analysis since only one animal was recorded at each of these time points.
Visual behavior tests
Results for navigation time and number of collisions of the two trials in each maze navigation test were averaged and were subjected as such, or after log transformation, to analysis of variance. For analysis of the overall data set, the statistical models included the effects of the transgene (mouse or human CNGA3), animal (n = 9), and follow-up time (months) from the treatment. For calculating the least squares mean (LSM) of the maze navigation time and number of collisions, the results were grouped into follow-up groups. For further analysis, the animal effect was replaced in the model with the sex of the animal.
The statistical model for analysis of the data subset collected when the maze test was conducted using alternate eye patching included the effects of transgene, animal nested within transgene, time after treatment (follow-up group), patched eye, and vector x patched eye interaction. A p-value <0.05 was considered significant. Results are expressed as LSM ± SE.
ERG
The following photopic ERG parameters were analyzed for each eye: 30- and 40-Hz flicker amplitudes and CFFF values, at 10 cd·s/m2. For scotopic recordings, b-wave amplitudes of the final (20 min) response were analyzed.
The collected data were analyzed with SPSS version 22.0 (IBM Corporation, Armonk, NY), and with STATA, version 14.0 (StataCorp, College Station, TX). We favored the linear mixed-effect model approach over repeated measures mixed-design since the linear mixed-effect model can tolerate some degree of imbalanced data, which were introduced by animals that were not included in each follow-up group or by noisy and unmeasurable responses in certain recordings. The fixed effects were transgene and follow-up groups while the random effect was the animal. A p-value <0.05 was considered statistically significant.
Results
Visually guided behavior testing
The association between time from treatment and maze navigation time is illustrated in Fig. 1A. Mean maze navigation time was >25 sec in the pretreatment period (follow-up group −1) and, within weeks after treatment, significantly dropped to that of unaffected control animals and remained statistically unchanged (p > 0.05) for the duration of the follow-up period. Analysis of the whole data set showed that, for maze navigation time, time from treatment (follow-up group) had a significant effect (p = 0.001) while the effects of the transgene and animal were not significant (p > 0.05). Sex had no significant effect on navigation time (p > 0.05).
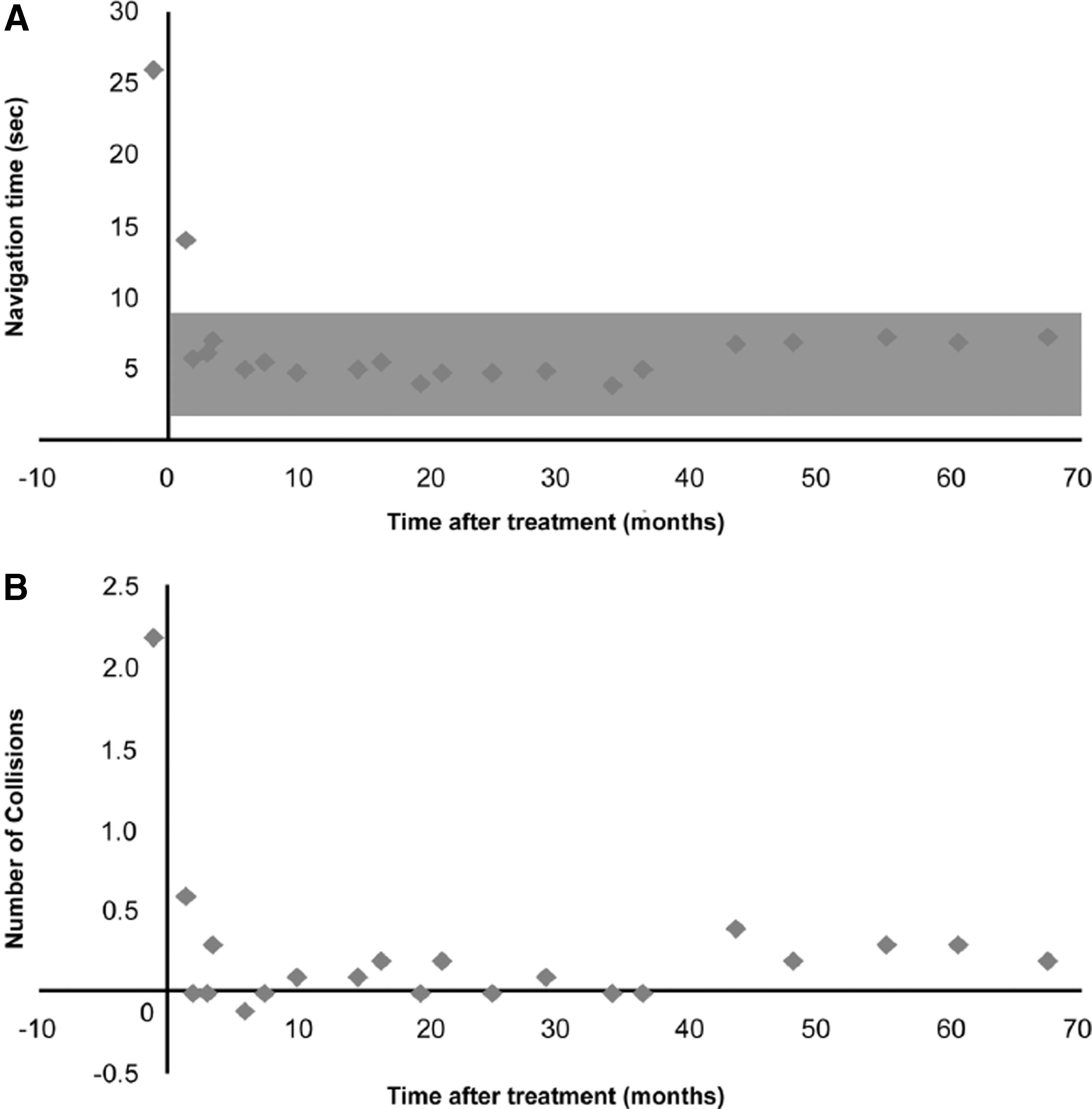
Long-lasting improvement in photopic maze testing of nine day-blind sheep that underwent gene augmentation therapy with the mouse or human CNGA3 transgene. Results are presented as the group's Least Square Mean.
The association between time from treatment and number of collisions with obstacles during maze navigation is illustrated in Fig. 1B. The mean number of collisions was >2 in the pretreatment period (follow-up group −1) but within weeks after treatment significantly dropped (< 0.5 collisions) and remained statistically unchanged for the duration of the follow-up period. Analysis of the number of collisions with obstacles during maze navigation demonstrated that time from treatment had a significant effect (p < 0.001). The transgene treatment also had a significant effect on the number of collisions (p = 0.01) with an average of 0.4 and 0.1 collisions for animals treated with the human CNGA3 and the mouse CNGA3 transgenes, respectively. The specific animal did not have a significant effect on the number of collisions (p > 0.05).
Analysis of the subset of maze navigation tests conducted using alternate patching of eyes demonstrated that patching either the treated (right) or the untreated (left) eye had a significant effect (p < 0.0001) on navigation time. Average navigation time when the treated (right) eye was patched was 26.8 ± 0.7 seconds, significantly (p < 0.05) longer than the navigation time when the untreated (left) eye was patched (8.7 ± 0.7 sec) or when neither eye was patched (6.3 ± 0.7 sec). The transgene had no effect on navigation time in this subset of maze tests (p = 0.18). Eye patching also had a significant effect (p < 0.0001) on the number of collisions with obstacles during the navigation test. Animals collided with obstacles 3.9 ± 0.2 times when the treated eye was patched and 0.3 ± 0.2 times when either the left eye or neither eye was patched. The transgene had a significant effect on the results of this subset of tests. The number of collisions was 4.8 ± 0.3 when the treated right eye was patched, and 3.1 ± 0.3 for animals treated with the human and the mouse transgenes, respectively (p < 0.05). However, no difference (p > 0.05) was found between the two transgene treatment groups when the left eye or neither eye was patched (Supplementary Video S1; Supplementary Data are available online at
ERG
ERG recordings showed significant, long-term improvement in cone function following CNGA3 augmentation therapy (Fig. 2). Representative flicker responses to 30–80 Hz stimuli (at 10 cd·s/m2) are shown in Fig. 2A. Short- and long-term improvement of CFFF at all four light intensities is shown in Fig. 2B. CFFF values for treated eyes in all 11 follow-up groups were significantly higher (p < 0.0001) than preoperative values (follow-up group −1). On the other hand, CFFF values of the untreated control eyes were not significantly different (p > 0.05) from preoperative values (Table S3).
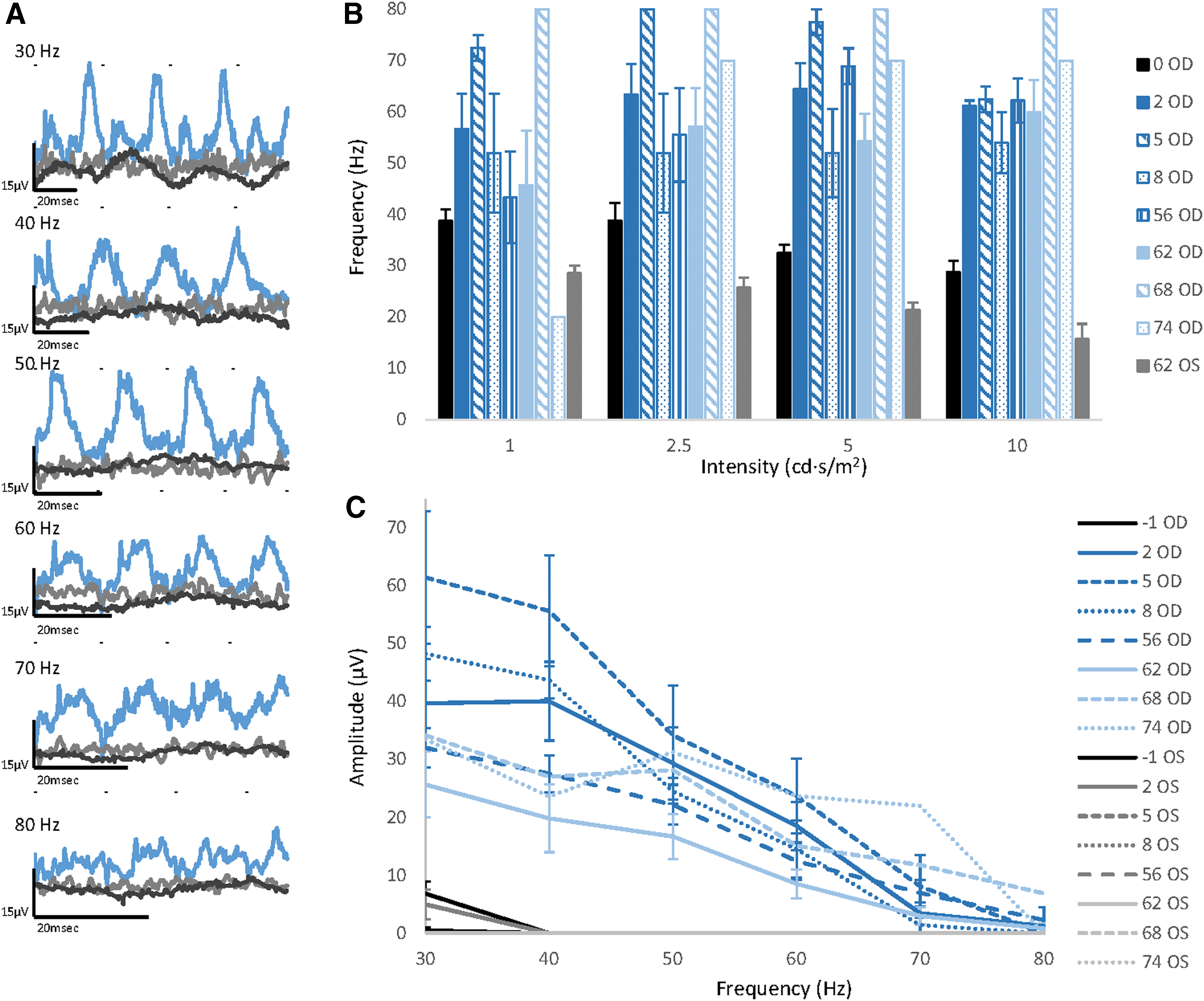
Long-lasting improvement in electroretinography (ERG) recordings of nine day-blind sheep that underwent gene augmentation therapy with the mouse or human CNGA3 transgene.
Responses to the 10 cd·s/m2 30–80 Hz flicker stimuli, plotted as a function of the stimulus frequency, are shown in Fig. 2C. The 30 and 40 Hz flicker amplitudes of treated (right) eyes in all 11 follow-up groups were significantly higher (p < 0.006) than preoperative values (follow-up group −1). On the other hand, the 30 and 40 Hz flicker amplitudes of the control (left) eyes were not significantly different (p > 0.05) than the preoperative values in 8/11 of the follow-up groups (Table S3).
Analysis of rod responses revealed no significant differences between treated and untreated eyes (p > 0.05, data not shown).
Safety, histology, and IHC
Full autopsy of sheep 4954, euthanized 67 months postoperatively due to postparturition complications, showed no external pathologic findings. Excessive amounts of clear serous fluid without fibrin strands were found in the pericardiac sac. No pathological changes were seen in the myocardium, endocardial valves, and papillary muscle of the heart. Diffuse hyperemia was apparent in all lung lobes. Consolidation and color changes were seen in the cranioventral lung lobe, affecting 10–15% of total lung volume. In the abdominal cavity there were chronic focal adhesions of the diaphragmatic liver lobes to the midventral diaphragm. Both kidneys were relatively soft (due to possible autolysis) and diffusely pale. There was focal, patchy, mild hyperemia of the serosa and mucosa of the duodenum and jejunum. No pathological findings were found in the brain and other organs. No abnormal histopathological findings were evident in the cerebrum, cerebellum, and midbrain.
Serological tests for paratuberculosis and Visna/maedi virus in six sheep (Table 1) were negative. Trueperella pyogenes was isolated from the lungs of sheep 4954.
Serial ophthalmic examinations during this follow-up study revealed no significant differences between treated and untreated eyes. Histological H&E sections of the eyes of sacrificed animals (Table 1) were examined by a pathologist. Vascular congestion in the choroid and sclera, as well as evidence of focal areas of retinal atrophy, were seen in the treated eye of sheep 4056 (Fig. 3A–D). An area of focal atrophy, as well as pigment granules, were seen in the treated retina of sheep 5187 (Fig. 3E). In addition, vascular congestion was noticed in the inner retina (Fig. 3F). In both animals, these areas of atrophy and congestion were apparently located within the treated areas because the blebs of sheep 4056 and 5187 were dorsal and ventral to the optic nerve heads, respectively.

Histology, 73 months postoperatively, of the posterior segment of the right eye of sheep 4056 treated with the mouse CNGA3 transgene
IHC showed expression of the CNGA3 protein colocalized with red/green opsin in treated eyes of animals 5187 (treated with the human transgene) and 4056 and 3909 (treated with the mouse transgene) (Fig. 4A & B). No CNGA3 labeling was seen in the control eye of animal 3909 nor in the negative control (Fig. 4D & E). However, CNGA3 labeling was observed in the control eyes of animals 4056 and 5187 (Fig. 4C).

Immunohistochemistry of the retinas of two representative day-blind sheep (5187 and 3909) following gene augmentation therapy with the human and mouse CNGA3 transgene, respectively. Rows
Discussion
We report that, in day-blind sheep, gene augmentation therapy, applied with a single subretinal injection of an AAV5 vector carrying either the mouse or human CNGA3 transgene, results in recovery of cone photoreceptor function as demonstrated by both photopic behavioral maze tests and by ERG examinations (Supplementary Video S1). At the time of this writing, recovery has lasted an average of 62 months in our nine study animals, and 73.6 months in the longest surviving animal, though data collection continues in the five surviving animals (Table 1).
CNGA3 achromatopsia is currently investigated using several animal models including mice and dogs. 21,22 In mice, cone function rescue following gene augmentation therapy using different AAV vectors was demonstrated for a period of nine months. 15 To the best of our knowledge, gene augmentation therapy has not been reported in the canine CNGA3 achromatopsia model, but it has been reported in the canine CNGB3 achromatopsia model, where gene therapy using AAV5 was found to rescue cone function for at least 33 months. 23 Therefore, we believe that our ongoing study is currently the longest gene therapy follow-up report published in any animal model of achromatopsia.
Alongside achromatopsia, gene therapy is being investigated in several other retinal diseases, where large animal models are used for long-term follow up. Therapy using AAV2/5 in a canine model of CNGB1-deficient retinitis pigmentosa was found to rescue rod function for a period of 18 months, 24 and, in a canine model of X-linked retinitis pigmentosa, therapy using AAV2/5 was shown to arrest vision loss for a period of two years. 25 Gene augmentation therapy in an RPE65 canine model of LCA showed restoration of rod and cone function during 4–11 years of follow-up. 13 –18 These long-term studies in canine models of LCA have paved the way for clinical trials in patients, with follow-ups of 3–6 years reported in patients treated with AAV2/2, 13,26,27 and one year in patients treated with AAV2/4. 28 Based on these results, the FDA has recently approved Luxturna™ as a treatment for LCA. Similarly, positive results in our sheep model 12,20 have paved the way for clinical trials in CNGA3 achromatopsia patients (NCT02935517 and NCT02610582). As these clinical trials are ongoing, long-term follow-up of efficacy and safety results reported in the present study is important.
The question whether gene augmentation therapy based on episomal delivery of the transgene (as achieved with AAV vectors) will provide a stable rescue effect or one that may diminish over time is an open one. In contrast to insertion of the transgene into the host genome (as can be achieved with lentiviruses, for example), the DNA delivered by AAV vectors may degrade and become less effective even in nondividing cells like the photoreceptors. In the clinical arena, a number of reports on patients treated for RPE65 LCA suggest some loss of treatment effect and failure to fully arrest disease progression a few years following treatment. 26,27 However, even with the decline in benefit noted at six years post-treatment for RPE65 LCA patients, 27 vision function within the vector-treated area was still above the pretreatment baseline. The stable effect seen to date in the sheep model of CNGA3 achromatopsia may be due to delivery of a higher dose of the vector, the young age of the animals at the time of treatment, and/or the fact that achromatopsia is largely nonprogressive in nature, 1,29,30 such that an underlying degenerative process at the cellular level is not present. Nonetheless, our results reflect the need to study the benefit over a period even longer than six years in both animal models and in human patients.
The improvement in photopic visual behavior and ERG recordings of the treated eyes was significant throughout the study, and no time-dependent attenuation was noted in any of the parameters analyzed. Our IHC analysis also demonstrated the long-lasting effect of the treatment. Expression of the CNGA3 protein, colocalized with red/green opsin, was seen in the treated eyes of the animals sacrificed 58–73 months postoperatively. However, animals 4056 and 5187, that were treated in the right eye with a mouse and a human CNGA3 transgene, respectively, unexpectedly also showed suspected CNGA3 immunolabeling in the untreated left eye (Fig. 4). Nonetheless, results of our ERG recordings and behavioral tests did not reflect any photopic retinal function in the control eyes of these sheep. Thes findings may indicate that only a small number of cones were transfected in the contralateral eyes of animals 4056 and 5187 because our IHC finding is qualitative rather than quantitative. Furthermore, studies in the RPE65 dog model of LCA have shown that, on occasion, there is no correlation between ERG rescue and numbers of surviving photoreceptors. 31 Admittedly, the last ERG recordings and maze tests of these two sheep were conducted 4–9 months prior to death and our IHC findings, but we do not believe that vector distribution and expression in the left eyes occurred in this short interval.
Boye et al studied guanylate cyclase-1 knockout mice treated subretinally in one eye with AAV and found vector genomes in the ipsilateral optic nerve and in the contralateral hemisphere of the brain. 32 Biodistribution of the viral vector was also investigated in nonhuman primates following subretinal delivery of AAV5 carrying the human rhodopsin kinase transgene. 33 Vector genome was recovered from the injected retinas, ipsilateral optic nerves, and select parts of the brain, but no transgene expression was detected outside the retinal bleb. 33 Thus, it is possible that, in our animals, the vector reached the photoreceptors of the contralateral eye either through transneuronal passage from the ipsilateral to the contralateral retinal ganglion cells, possibly at the optic chiasm, or via systemic distribution. The latter route is less likely due to the dilution of the relatively small amount of vector (Table 1) in a 50-kg sheep and its exposure to the systemic immune system. Obviously, biodistribution studies in the optic nerves and chiasm, brain, and other organs are needed to establish the possible infection route.
We did not detect immunolabeling of CNGA3 in untreated control eyes during our previous gene-augmentation sheep studies. 12,20 Therefore, we consider the findings in the control eyes of 4056 and 5187 as preliminary results. Since we have only five surviving treated animals, which represent a unique resource that allows such an ongoing long follow-up, we feel it would be improper to sacrifice them to verify these findings and study the important implications of our IHC findings. Instead, we believe that a prospective study with a new set of experimental animals, as well as additional animal models, and with the use of RNA assays, should be conducted. Additional PCR and IHC studies will be performed in the five surviving sheep when they are sacrificed or die.
Another important issue, in addition to treatment efficacy, is that of safety. Repeated postoperative ophthalmic examinations of the sheep by a board-certified veterinary ophthalmologist showed no significant abnormalities in treated eyes during the study years. In addition, scotopic ERG recordings conducted at the last follow-up examination revealed no impairment of rod function in treated eyes. In two treated eyes, histopathology showed focal areas of retinal atrophy and vascular congestion (Fig. 3). However, despite these lesions, the last antemortem ERG recording and maze test of these eyes showed robust photopic function, not significantly different from that of eyes of the other experimental animals. Interestingly, the two animals in which these lesions were found (in their treated eyes) are the same animals in which IHC staining revealed CNGA3 protein expression in the untreated eyes. However, our surgical records do not document any intravitreal reflux or any other intraoperative protocol deviations that can explain the inflammation or viral distribution in these animals. Nonetheless, it is possible that micro trauma to the RPE, caused when the cannula penetrated the sensory neuroretina (E. Averbukh, unpublished results), may be responsible for some of the pathology in the operated eyes.
Since our last report, 12 four sheep have been euthanized 58–73 months postoperatively. The animals were 5.5–6.3 years old at the time of death and had been anesthetized 7–9 times for surgery and ERG recordings (Table 1). Two animals (3909 and 4954) were euthanized following postparturition complications, and the other two (4056 and 5187) exhibited signs of a wasting disease a few months before death. Though their behavior and appetite were unaffected, they lost weight and were classified as cachectic by a board-certified specialist in small ruminant medicine. Serological testing for the two leading causes of wasting diseases in sheep (paratuberculosis and Visna/maedi) was negative. Autopsy did not reveal a specific cause of death. The hyperemia noted in the lungs and intestines can result from premortem stress and agony. A combination of parturition complications and pneumonia caused by the isolated Trueperella pyogenes, a bacterium that can cause abscesses, mastitis, metritis, and pneumonia in sheep, might explain the animals' extreme exhausted status that finally resulted in recumbency and euthanasia.
In conclusion, we report successful restoration of photopic vision in nine day-blind sheep, following gene augmentation therapy with AAV5 carrying the mouse/human CNGA3 transgene, that lasted more than six years. We intend to continue monitoring the remaining five animals and perform periodic follow-up evaluations using ERG recordings and behavioral tests. We believe that it is essential to preserve these unique and precious animals that currently represent the longest surviving patients of gene augmentation therapy reported in an animal model of achromatopsia.
Footnotes
Acknowledgments
This study was supported (in part) by grants from the U.S.-Israel Binational Science Foundation, the Israel Science Foundation, the Chief Scientist Office of the Israeli Ministry of Health, and the Macula Vision Research Foundation, as well as by unrestricted awards from The Joseph Alexander Foundation and Research to Prevent Blindness and a Yedidut 1 Research Grant. The authors thank Dr. Nir Edery for the pathological study.
Author Disclosure
WWH and the University of Florida have a financial interest in the use of AAV therapies and own equity in a company (AGTC) that might, in the future, commercialize some aspects of this work. All other authors have no competing financial interests.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.