
Abstract
Hemophilia A is a common hereditary bleeding disorder that is characterized by a deficiency of human blood coagulation factor VIII (hFVIII). Previous studies with adeno-associated viral (AAV) vectors identified two liver-specific promoter and enhancer combinations (E03.TTR and E12.A1AT) that drove high level expression of a codon-optimized, B-domain-deleted hFVIII transgene in a mouse model of the disease. This study further evaluated these enhancer/promoter combinations in cynomolgus macaques using two different AAV capsids (AAVrh10 and AAVhu37). Each of the four vector combinations was administered intravenously at a dose of 1.2 × 1013 genome copy/kg into five macaques per group. Delivery of the hFVIII transgene via the AAVhu37 capsid resulted in a substantial increase in hFVIII expression compared to animals administered with AAVrh10 vectors. Two weeks after administration of E03.TTR packaged within the AAVhu37 capsid, average hFVIII expression was 20.2 ± 5.0% of normal, with one animal exhibiting peak expression of 37.1% of normal hFVIII levels. The majority of animals generated an anti-hFVIII antibody response by week 8–10 post vector delivery. However, two of the five macaques administered with AAVhu37.E03.TTR were free of a detectable antibody response for 30 weeks post vector administration. Overall, the study supports the continued development of AAV-based gene therapeutics for hemophilia A using the AAVhu37 capsid.
Introduction
H
In contrast to the small size of the hFIX coding sequence (1.4 kb), the native hFVIII protein is a large multi-domain glycoprotein with complementary DNA (cDNA) that exceeds the packaging capacity for recombinant AAV (>7 kb). 6,7 To overcome this limitation, extensive engineering of both the expression cassette and the hFVIII cDNA is required. Previously, the protein replacement therapy drug, ReFacto® (Pfizer, New York, NY), was successfully designed to mimic the smallest active form of hFVIII by replacing the B domain with a 14 amino acid SQ linker. 8 Subsequent codon optimization of this B-domain-deleted (BDD) hFVIII-SQ (hFVIIIco-SQ) resulted in efficient packaging and increased expression from lentiviral 9,10 and AAV 11 vectors in vivo. Further minimal modifications, such as the addition of a synthetic spacer sequence 11 or deletion of the furin recognition motif, 12,13 have been shown to enhance significantly the biological activity of BDD hFVIII-SQ expressed from AAV.
Efficient transduction of human hepatocytes is critical for the successful application of AAV-mediated gene therapy for the treatment of hemophilia in the clinic, and optimization of transcriptional control elements can improve transgene expression in the liver. The large open reading frame of the BDD hFVIII-SQ transgene precludes the use of full-length liver-specific promoter and enhancer elements for packaging into an AAV vector. Therefore, identifying small functional transcriptional control sequences is essential for advancing AAV-mediated gene therapy for the treatment of hemophilia A. Indeed, the incorporation of small computationally identified liver-specific transcriptional cis-regulatory modules into AAV vectors expressing hFIX were shown to enhance expression in mice and cynomolgus macaques. 14 –16
Using enhancer/promoter combinations that limited the total genome size to <5,250 bp, extensive optimization and in vivo evaluation of 42 BDD hFVIIIco-SQ expression cassette variants was previously performed. 17 These variants were packaged into the AAVrh10 capsid and administered intravenously (i.v.) into FVIII knockout (KO) mice at a dose of 1010 genome copies (GCs) per mouse. Two weeks after injection, mice with five of the variants exhibited significantly higher hFVIII activity levels. Anti-hFVIII antibodies were absent at week 2, although anti-hFVIII antibodies developed later in the majority of groups. Subsequently, the impact of capsid was evaluated by packaging the variant that exhibited the highest immunogenicity into AAV8, AAV9, AAVrh10, AAVhu37, and AAVrh64R1 capsids. The AAVrh10 capsid resulted in the highest levels of hFVIII activity but also the highest levels of anti-hFVIII antibodies, whereas AAVhu37 produced intermediate results in terms of expression and immunogenicity. Based on these results, the AAVrh10 and AAVhu37 capsids and the E03.TTR and E12.A1AT enhancer/promoter combinations were selected for further evaluation in nonhuman primates (NHPs).
Methods
AAV vector production
All AAV vectors were produced by the Penn Vector Core at the University of Pennsylvania, as previously described. 18 Briefly, plasmids expressing hFVIIIco-SQ from either the E03.TTR or E12.A1AT enhancer/promoter combinations were packaged into the AAVrh10 or AAVhu37 capsid. E03.TTR or E12.A1AT enhancer/promoter combinations were selected from extensive in vivo analysis performed previously in mice. 17 E03.TTR is the EnTTR (100 bp enhancer sequence from transthyretin) enhancer in combination with the TTR (transthyretin) promoter, and E12.A1AT is a combination of two copies of ABPS (100 bp distal enhancer from the α1-microglogulin/bikunin precursor shortened to 42 bp) enhancer, plus one copy of EnTTR with the A1AT (modified SERINA1 [α1-antitrypsin]) promoter.
Neutralizing antibody assay
Neutralizing antibodies (NAbs) to AAV8, AAVrh10, AAVhu37, AAV5, and AAV-LK03 were determined in 28 human serum samples from the University of Pennsylvania Hospital blood bank subjects, as previously described. 19,20 The samples were de-identified and a coded number assigned to each one of them.
NHPs
Male cynomolgus macaques were housed at the Children's Hospital of Pennsylvania Nonhuman Primate Facility (Philadelphia, PA) during the studies. All studies were performed according to a study protocol approved by the Institutional Animal Care and Use Committee, the Environmental Health and Radiation Safety Office, and the Institutional Biosafety Committee of the University of Pennsylvania.
All macaques had initial NAb titers of <1:5 to both AAVrh10 and AAVhu37 capsids, determined as previously described. 19,20 Prior to vector administration, macaques were anesthetized with a mixture of ketamine (10–15 mg/kg) and dexmedetomidine (0.05–0.10 mg/kg) injected intramuscularly. Macaques were administered with vectors i.v. via the saphenous vein. Blood samples were taken prior to the initiation of the study and biweekly during the study via venipuncture of the femoral vein. All clinical pathology tests on blood samples were conducted by Antech Diagnostics (Irvine, CA), including complete blood counts and differentials, clinical chemistries, and coagulation panels.
Twenty male cynomolgus macaques were administered i.v. with a dose of 1.2 × 1013 GC/kg of one of four vectors (n = 5 macaques per vector): AAVrh10.E03.TTR.hFVIIIco-SQ.PA75, AAVrh10.E12.A1AT.hFVIIIco-SQ.PA75, AAVhu37.E03.TTR.hFVIIIco-SQ.PA75, or AAVhu37.E12.A1AT.hFVIIIco-SQ.PA75.
Immunosuppression protocol
An immunosuppression regimen was initiated as required in animals where the ability to detect hFVIII expression was lost due to the presence of detectable antibodies to hFVIII (Bethesda unit [BU] >1) following vector administration. The immunosuppression regimen was performed with rituximab (250 mg/m2 i.v. at four-weekly intervals; a total of four infusions) and cyclophosphamide (300 mg/m2 slow i.v. infusion every 15 days; a total of eight doses over 4 months), as previously described. 11
Determination of hFVIII expression in NHP plasma
hFVIII expression was measured by an enzyme-linked immunosorbent assay (ELISA), where all reagents were from Sigma–Aldrich (St. Louis, MO) unless otherwise stated. ELISA plates were coated with anti-hFVIII immunoglobulin G (IgG; Green Mountain Antibodies, Burlington, VT) at a 1:500 dilution in 0.1 M of carbonate buffer (pH 9.6) and incubated overnight at 4°C. Wells were washed four times with 0.1% Tween 20 in phosphate-buffered saline (PBS) and blocked with 5% nonfat milk (Bio-Rad Laboratories, Hercules, CA) in PBS for 1 h at room temperature. Following removal of the blocking buffer, plasma samples diluted in 5% nonfat milk were added to the plates and incubated for 1 h at room temperature. A standard curve was generated for each NHP run on a plate by performing twofold serial dilutions of hFVIII recombinant protein (ReFacto®; Pfizer) in 5% nonfat milk with 4% naïve plasma from the NHP being tested. Plates were then washed four times, and anti-hFVIII IgG (Thermo Fisher Scientific, Waltham, MA) was added at a 1:1,000 dilution in nonfat milk. Following incubation for 1 h at room temperature, plates were washed four times, and horseradish peroxidase (HRP)-conjugated anti-sheep IgG was added at a 1:1,000 dilution in nonfat milk. Following incubation at room temperature for 90 min, plates were washed five times and 3,3′,5,5′-tetramethylbenzidine (TMB) was added for detection. The reaction was stopped after 5 min at room temperature using 2N sulfuric acid, and plates were read at 450 nm using a BioTek μQuant plate reader (Winooski, VT).
Detection of anti-hFVIII IgG in NHP plasma
IgG antibodies against hFVIII in NHP plasma were measured by ELISA. ELISA plates were coated with 1 μg/mL of BDD hFVIII-SQ (Xyntha; Wyeth Pharmaceuticals Inc., Dallas, TX) in 0.1 M of carbonate buffer (pH 9.6) and incubated overnight at 4°C. Wells were washed five times with 0.05% Tween 20 in PBS and blocked with 5% nonfat milk (Bio-Rad Laboratories) in PBS for 1 h at room temperature. Following removal of the blocking buffer, plasma samples diluted starting at a 1:100 dilution in 5% nonfat milk were added to the plates and incubated for 1 h at room temperature. Naïve NHP plasma samples were used as the control. Plates were then washed five times, and HRP-conjugated anti-NHP IgG was added at a 1:2,000 dilution in nonfat milk. Following incubation at room temperature for 90 min, plates were washed eight times and then TMB was added for detection. The reaction was stopped after 5 min at room temperature using 2N sulfuric acid, and plates were read at 450 nm using a BioTek μQuant plate reader. Values fivefold over background levels (naïve NHP samples) were considered positive.
hFVIII inhibitor assay
hFVIII inhibitors were measured by the Bethesda assay with the Nijmegen modification. 21 NHP plasma samples were first incubated at 56°C for 1 h to inactivate NHP FVIII. Twofold dilutions of the test plasma in FVIII-deficient human plasma (George King Bio-Medical, Inc. Overland Park, KS) were then incubated with an equal volume of pooled normal human plasma (George King Bio-Medical, Inc.), together with a control mixture of one volume of imidazole-buffered pooled normal human plasma (pH 7.4) with one volume of FVIII-deficient human plasma. Following incubation at 37°C for 2 h, the residual FVIII activity was determined using one-step activated partial thromboplastin time (PTT). One BU was defined as the reciprocal of the dilution of test plasma at which 50% of human FVIII activity was inhibited. The sensitivity of the assay was 1 BU/mL.
Vector biodistribution
Tissues samples were snap frozen at the time of necropsy, and DNA was extracted using the QIAamp DNA Mini Kit (Qiagen, Valencia, CA). Detection and quantification of vector GCs in extracted DNA were performed by real-time PCR, as previously described. 22 Briefly, genomic DNA was isolated, and vector GCs were quantified using primers and probe designed against the hFVIII sequence of the vector.
RNA isolation and real-time PCR
RNA was isolated from tissue samples, and cDNA was generated, as previously described. 23 Real-time PCR was then performed on cDNA with primers binding to the hFVIII transgene with TaqMan Gene Expression Master Mix for detection or with the primer/probe set for 18S ribosomal RNA (Life Technologies, Carlsbad, CA). Relative transcript expression was determined using the ΔΔCT of each sample normalized to 18S expression, as previously described. 23
Statistical analysis
To identify differences in hFVIII expression in plasma at peak, the data were analyzed using a single fixed factor analysis of variance model with Tukey post hoc tests to identify differences in animal and group mean activity levels. Time-to-event analysis was performed for the generation of antibodies against hFVIII.
Results
Selection of AAV capsids for expression
Two vector constructs were selected to be evaluated in large animals—E03.TTR and E12.A1AT— based on the data generated in a previous FVIII KO mouse study.
17
These constructs were then packaged within AAVrh10, which is a capsid that has been evaluated in the clinic for other indications, and AAVhu37, as it produced intermediate results in terms of expression and immunogenicity. AAVrh10 has a similar NAb prevalence in the human population as AAV8 (55% and 52% are negative for NAbs, respectively), whereas fewer individuals have detectable levels of NAbs to AAVhu37 (66% are negative for NAbs; Supplementary Fig. S1; Supplementary Data are available online at
Evaluation of hFVIII expression in cynomolgus macaques
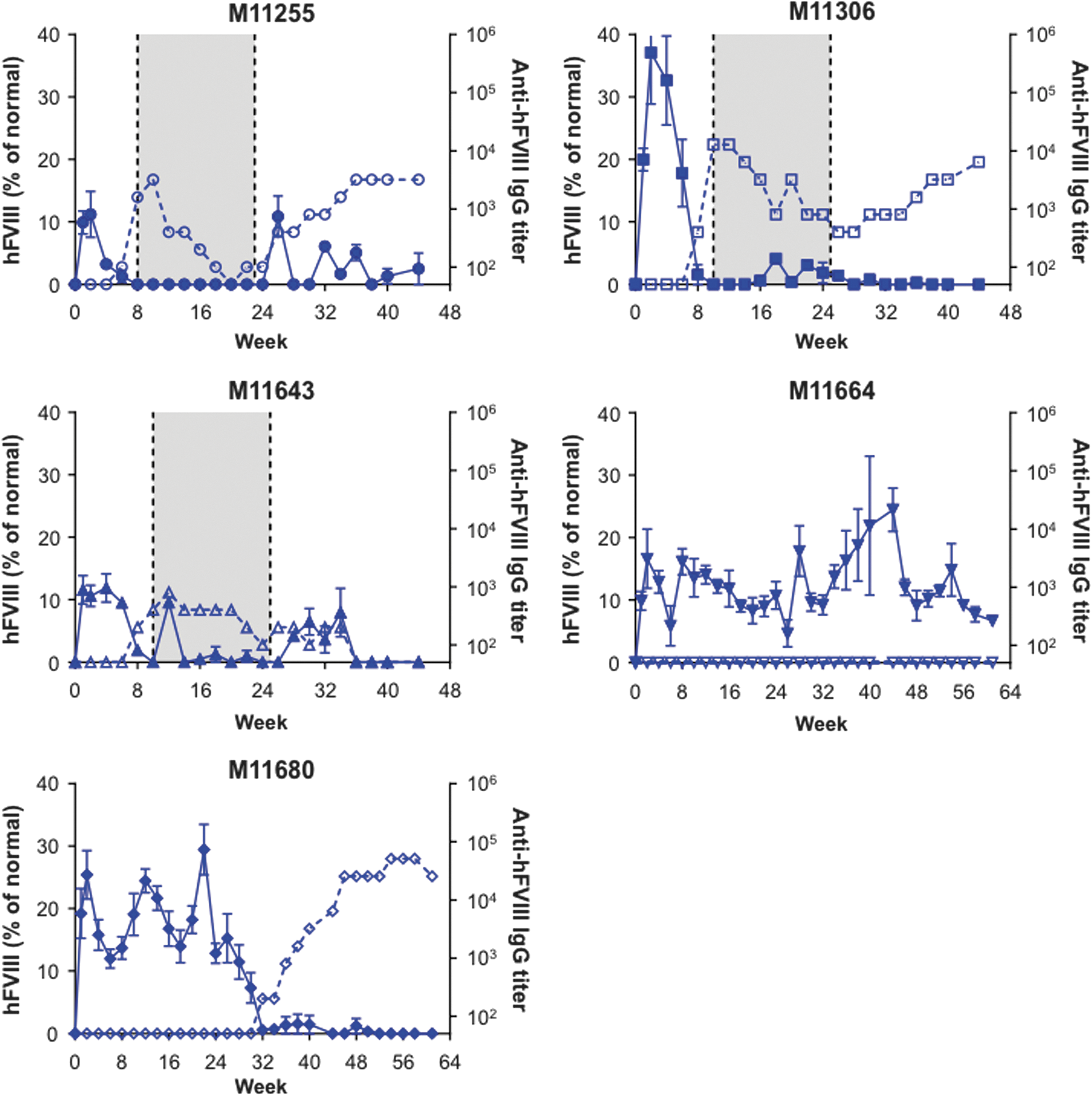
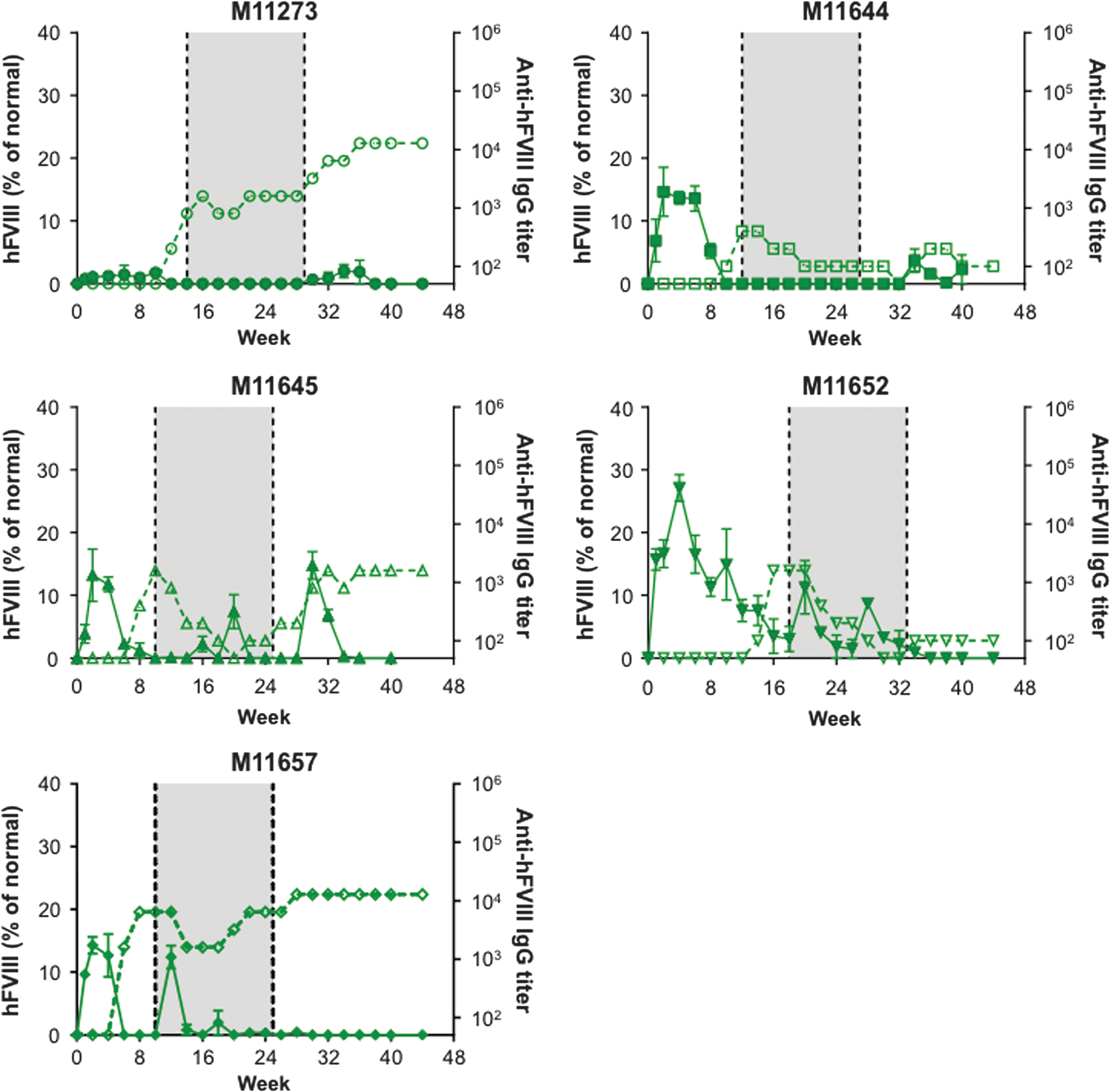
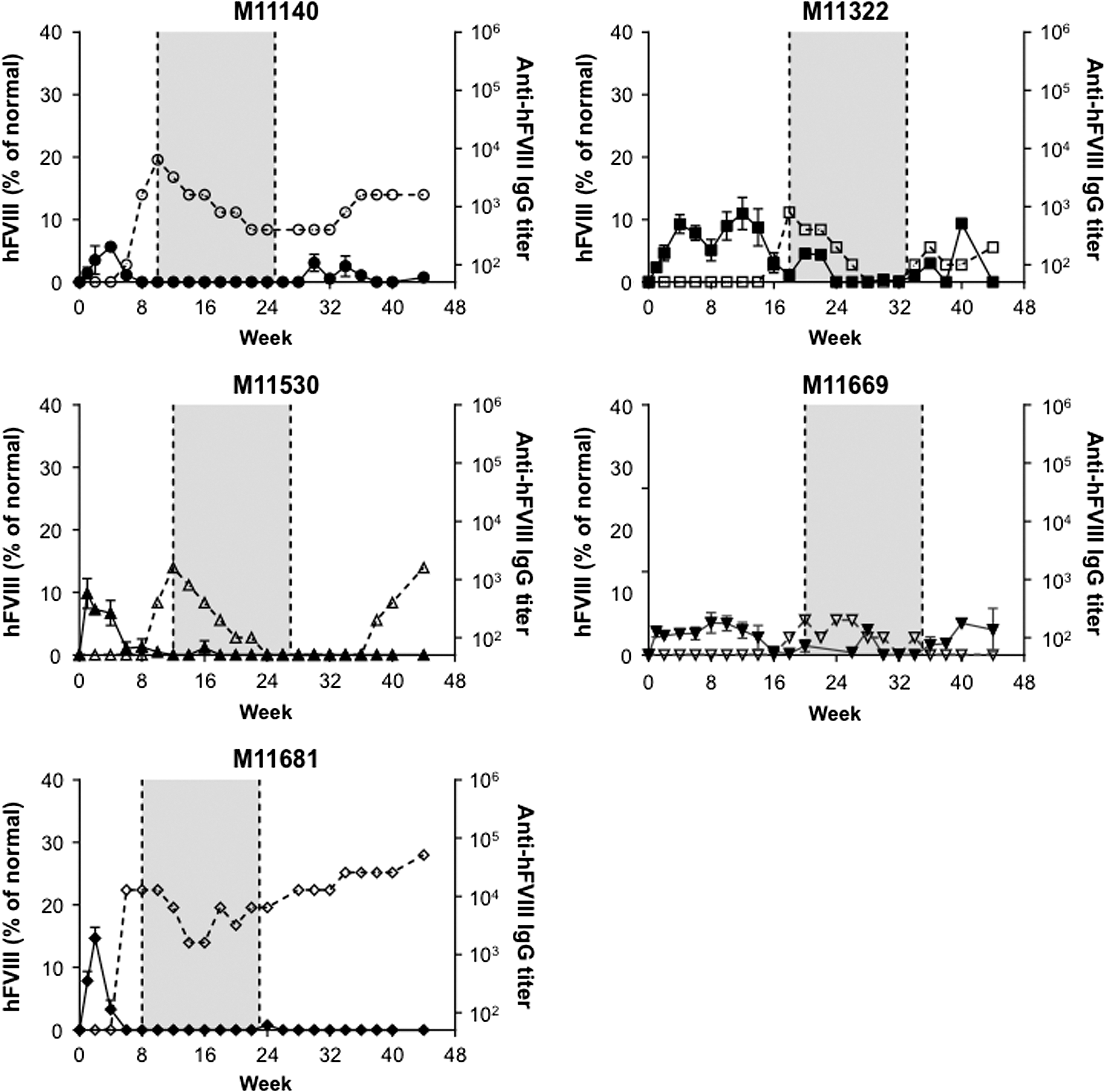
Twenty male cynomolgus macaques were administered i.v. with a dose of 1.2 × 1013 GC/kg of one of the following four vectors: AAVrh10.E03.TTR.hFVIIIco-SQ.PA75 (Fig. 1), AAVhu37.E03.TTR.hFVIIIco-SQ.PA75 (Fig. 2), AAVrh10.E12.A1AT.hFVIIIco-SQ.PA75 (Fig. 3), or AAVhu37.E12.A1AT.hFVIIIco-SQ.PA75 (Fig. 4). The macaques were bled biweekly to evaluate hFVIII expression in plasma and anti-hFVIII antibody titers. Following the loss of detectable hFVIII expression and detection of anti-hFVIII antibodies, an hFVIII inhibitor assay was performed to determine the Bethesda levels. If the BUs were >1, an immunosuppression regimen was initiated (250 mg/m2 rituximab i.v. at four-weekly intervals, for a total of four infusions; 300 mg/m2 cyclophosphamide by slow i.v. infusions every 15 days, for a total of eight doses over 4 months, as previously described 11 ). In Figs. 1 –4, the shaded areas indicate the application of the immunosuppression protocol.
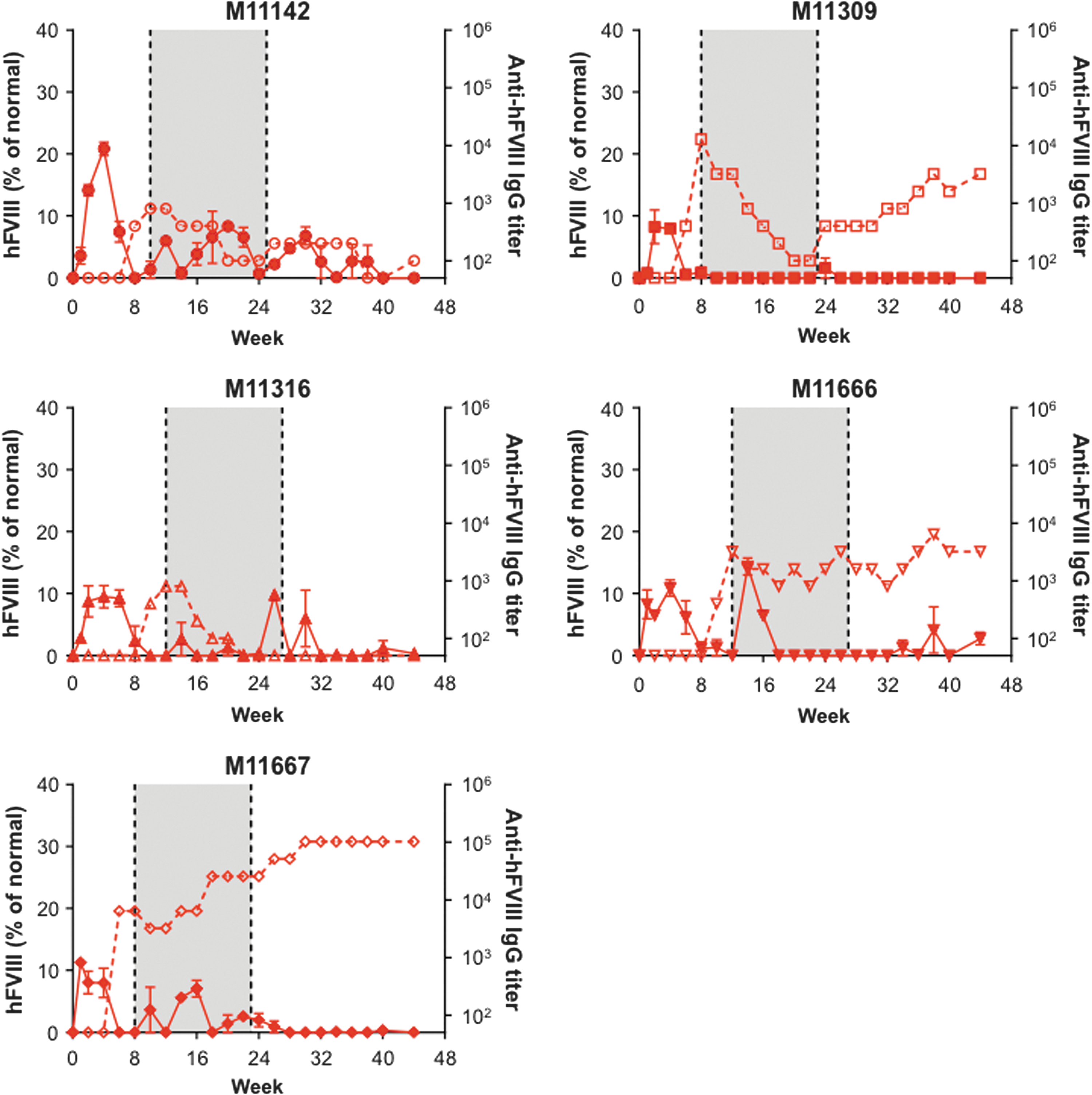
Human blood coagulation factor VIII (hFVIII) expression and anti-hFVIII antibody generation in individual cynomolgus macaques following administration of AAVrh10.E03.TTR.hFVIIIco-SQ.PA75. Five male rhesus macaques were administered intravenously (i.v.) with 1.2 × 1013 genome copies (GC)/kg of AAVrh10.E03.TTR.hFVIIIco-SQ.PA75. Macaques were bled biweekly to evaluate hFVIII expression in plasma by enzyme-linked immunosorbent assay (ELISA). Values are expressed as mean ± standard error of the mean (SEM; solid line). Anti-hFVIII immunoglobulin G (IgG) titers were also evaluated biweekly in plasma by ELISA (1/dilution; dashed line). The shaded area indicates the application of the immunosuppression protocol as required.
hFVIII expression and anti-hFVIII antibody generation in individual cynomolgus macaques following administration of AAVhu37.E03.TTR.hFVIIIco-SQ.PA75. Five male rhesus macaques were administered i.v. with 1.2 × 1013 GC/kg of AAVhu37.E03.TTR.hFVIIIco-SQ.PA75. Macaques were bled biweekly to evaluate hFVIII expression in plasma by ELISA. Values are expressed as mean ± SEM (solid line). Anti-hFVIII IgG titers were also evaluated biweekly in plasma by ELISA (1/dilution; dashed line). The shaded area indicates the application of the immunosuppression protocol as required.
hFVIII expression and anti-hFVIII antibody generation in individual cynomolgus macaques following administration of AAVrh10.E12.A1AT.hFVIIIco-SQ.PA75. Five male rhesus macaques were administered i.v. with 1.2 × 1013 GC/kg of AAVrh10.E12.A1AT.hFVIIIco-SQ.PA75. Macaques were bled biweekly to evaluate hFVIII expression in plasma by ELISA. Values are expressed as mean ± SEM (solid line). Anti-hFVIII IgG titers were also evaluated biweekly in plasma by ELISA (1/dilution; dashed line). The shaded area indicates the application of the immunosuppression protocol as required.
hFVIII expression and anti-hFVIII antibody generation in individual cynomolgus macaques following administration of AAVhu37.E12.A1AT.hFVIIIco-SQ.PA75. Five male rhesus macaques were administered i.v. with 1.2 × 1013 GC/kg of AAVhu37.E12.A1AT.hFVIIIco-SQ.PA75. Macaques were bled biweekly to evaluate hFVIII expression in plasma by ELISA. Values are expressed as mean ± SEM (solid line). Anti-hFVIII IgG titers were also evaluated biweekly in plasma by ELISA (1/dilution; dashed line). The shaded area indicates the application of the immunosuppression protocol as required.
All macaques expressed hFVIII, with the exception of M11273 administered with AAVrh10.E12.A1AT, where the expression was barely above detectable levels throughout the course of the study (Fig. 3). This lack of expression in macaque M11273 may be due to NAbs to capsid that were either not detected at the time of screening (1 month prior to vector administration) or due to seroconversion between the time of screening and vector administration. Although peak hFVIII expression levels were remarkably consistent across the five macaques administered with AAVhu37.E12.A1AT (Fig. 5D), the groups administered with AAVrh10.E03.TTR, AAVhu37.E03.TTR, and AAVrh10.E12.A1AT showed significant variations in hFVIII levels (Fig. 5A–C). In each of these groups, one macaque expressed significantly more hFVIII than the other animals (M11142, M11306, and M11652, respectively). At peak, the average hFVIII expression was 12.2 ± 2.2%, 23.4 ± 5.1%, 14.2 ± 4.0%, and 9.8 ± 1.5% of normal for AAVrh10.E03.TTR, AAVhu37.E03.TTR, AAVrh10.E12.A1AT, and AAVhu37.E12.A1AT vectors, respectively (mean ± standard error of the mean; Fig. 5E).
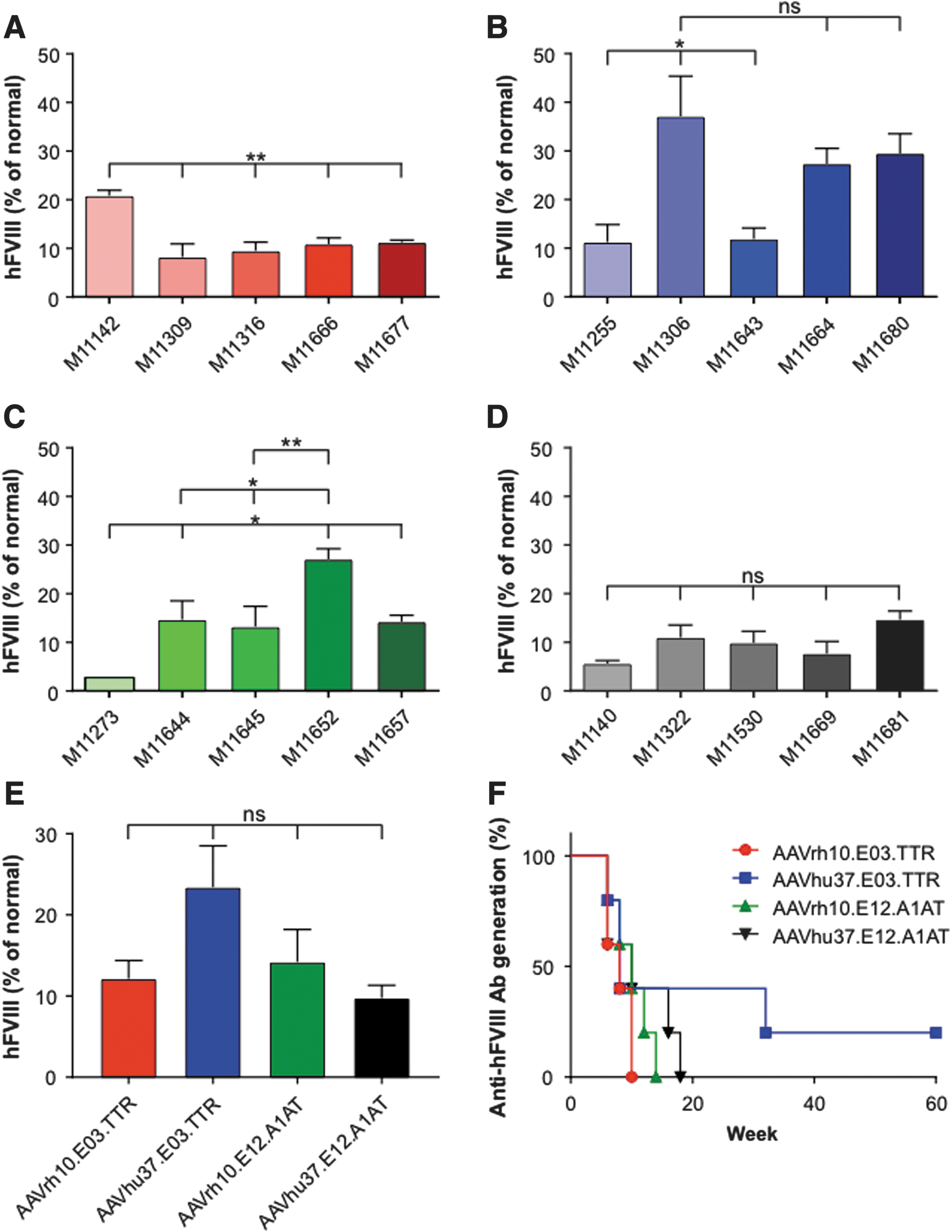
Comparison of peak hFVIII expression in cynomolgus macaques. Five male cynomolgus macaques were administered i.v. with 1.2 × 1013 GC/kg of
Generation of anti-hFVIII antibodies in cynomolgus macaques
All macaques administered with AAVrh10.E03.TTR (Fig. 1), AAVrh10.E12.A1AT (Fig. 3), and AAVhu37.E12.A1AT (Fig. 4), and three of the five macaques administered with AAVhu37.E03.TTR (Fig. 2), developed anti-hFVIII antibodies during the first 30 weeks post vector administration. Development of anti-hFVIII antibodies in nearly all animals was perhaps unsurprising, as the human FVIII protein is known to be immunogenic in NHPs. Time-to-event analysis was performed for the generation of antibodies against hFVIII (Fig. 5F). All macaques administered with AAVrh10.E03.TTR, AAVrh10.E12.A1AT, and AAVhu37.E12.A1AT developed antibodies by weeks 10, 14, and 18, respectively.
These 18 animals that developed detectable anti-hFVIII antibodies were evaluated for the presence of inhibitors by the Bethesda assay. All animals with detectable anti-hFVIII antibodies also had Bethesda titers of >1 BU (range 1–795.7 BU; median 5 BU) at the same time point and were subjected to the immunosuppression protocol when anti-hFVIII antibodies developed, which varied between 8 and 20 weeks following gene transfer. The effect of immunosuppression in these cynomolgus macaques fell into three categories: (1) a decrease in anti-hFVIII antibody titer during the immunosuppression regimen that then remained undetectable following withdrawal of immunosuppression, (2) a decrease in anti-hFVIII antibody titer during the immunosuppression regimen that then reverted following completion of immunosuppression, and (3) no effect of immunosuppression on anti-hFVIII antibody titer. Expression of hFVIII declined to undetectable levels with the appearance of anti-hFVIII antibody in all animals and remained low, even in animals in which the antibodies subsequently declined as a result of immunosuppression.
Two of the five macaques administered AAVhu37.E03.TTR did not develop anti-hFVIII antibodies during the first 30 weeks post vector administration (M11664 and M11680; Fig. 2). M11664 continued to express hFVIII at around 12% of normal throughout the duration of the study. However, at week 32 post vector administration, M11680 generated anti-hFVIII antibodies that coincided with a decline in hFVIII to barely detectable levels (Fig. 2). All animals had vector GCs and hFVIII mRNA detectable in the liver at the time of necropsy; the presence or absence of anti-hFVIII antibodies did not affect transcript level (Supplementary Fig. S3A and B).
Influence of anti-hFVIII antibodies on the coagulation pathway
Due to the concern over a potential interaction between the expressed transgene (hFVIII) and the coagulation pathway, prothrombin time (PT) and PTT values were monitored throughout the study as part of the coagulation panel testing performed by Antech Diagnostics (Fig. 6A–D and Supplementary Figs. S4–S7).
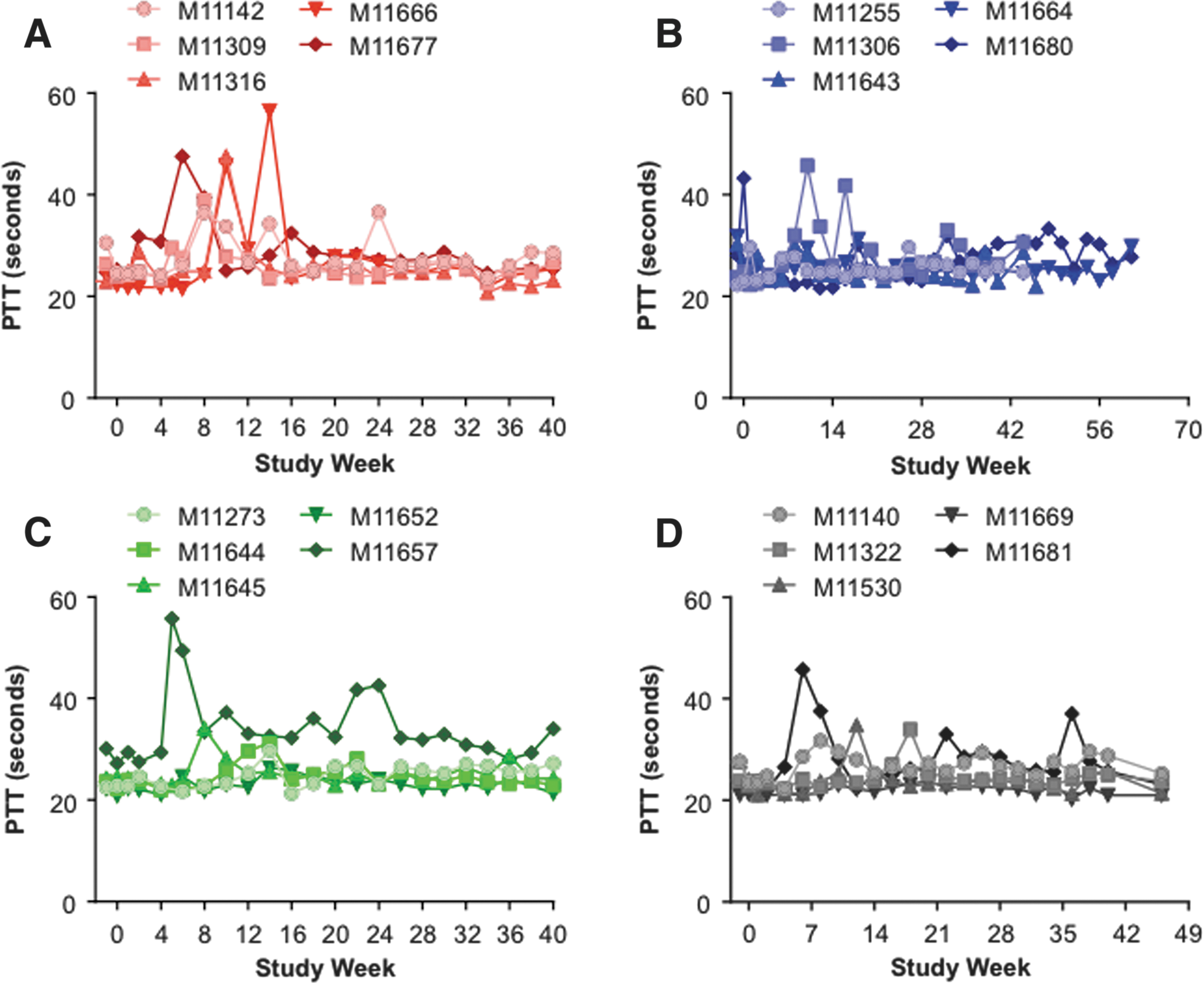
Partial thromboplastin time (PTT) values in vector-administered cynomolgus macaques. Five male cynomolgus macaques were administered i.v. with 1.2 × 1013 GC/kg of
Average PT values ranged from 10 to 14 s, and post vector administration PT values in the macaques only exceeded 14 s on two occasions. The PT was 14.8 s at week 18 for M11680, and 16.7 s at week 36 for M11681 (data not shown). All PT values returned to within normal limits by the following time point. However, more variability was observed in PTT values post vector administration (Fig. 6). Clotting, as determined by PTT, should occur within 25–35 s. On six occasions during the course of the study, PTT values >45 s were recorded but did not coincide with any observed coagulopathies. The exception to this was in macaque M11657 administered with AAVrh10.E12.A1AT, which had a severe bleeding event during the initial phase of the study due to an injury to the base of the tail (Fig. 6C). The PTT values in this animal remained >28 s for the duration of the study. In the majority of animals, the initial elevation in PTT values correlated with the first detection of anti-hFVIII IgG antibodies (Supplementary Figs. S4–S7). However, even in animals where the immunosuppression regimen was not able to decrease or control anti-hFVIII antibody titers (e.g., animals M11666, M11667, and M11306), the PTT returned to within normal limits in the majority of animals 2–4 weeks after the first peak levels. Sporadic elongation of PTT values were observed in some animals following the completion of the immunosuppression regimen when anti-hFVIII titers reverted toward the end of the study.
Discussion
AAV gene therapy has reached the clinic for the treatment of hemophilia A and B. 1 –5 However, vector development has been more challenging for the treatment of hemophilia A compared to hemophilia B due to the relatively large size of the hFVIII transgene sequence. Customization of the expression cassette for successful AAV packaging and expression of hFVIII requires modification and codon optimization of the transgene, as well as the use of small enhancer and promoter sequences. 8 –11 Previously, an extensive study was performed in a murine model of hemophilia A to identify the best promoter/enhancer combinations for liver-directed expression of BDD hFVIIIco-SQ. 17 No substantial differences in yield, vector protein composition, or genome integrity were found between the 42 variants evaluated. Thus, selection criteria for further evaluation in NHPs included analysis of hFVIII activity and immunogenicity. Of the top five expressing vectors, two candidates were selected for evaluation in NHPs: E03.TTR and E12.A1AT. In the present study, NHPs were administered with AAVrh10 or AAVhu37 vectors in combination with the E03.TTR and E12.A1AT enhancer/promoters.
Following systemic administration of AAVrh10.E03.TTR, AAVhu37.E03.TTR, AAVrh10.E12.A1AT, and AAVhu37.E12.A1AT vectors into cynomolgus macaques, peak hFVIII expression levels were observed (average of 12.2%, 23.4%, 14.2%, and 9.8% of normal, respectively). To remove the interference of endogenous macaque FVIII in the ELISA performed for determination of hFVIII transgene expression, NHP plasma samples obtained pre-dosing were added to a final concentration of 4% of the dilution buffer. This buffer containing 4% NHP plasma was used for dilution of all samples and to generate the standard curve. By performing the assay in this manner, the amount of endogenous macaque FVIII remained the same in all samples being tested and in the standards. Once inhibitors have developed, the ELISA is no longer able to detect the hFVIII levels accurately, even though the transduced hepatocytes may continuously produce hFVIII. Therefore, evaluation of peak hFVIII levels is more informative for the determination of efficacy.
Although the translation of results from mice to larger animal models can be difficult, the same systemic dose of vector generally translates to an approximate 10-fold decrease in expression from mice to NHPs. 25 This trend was consistent with the results, where peak hFVIII expression from similar doses of vector based on body mass decreased 14-fold in AAVrh10.E03.TTR-administered NHPs (12.2% of normal) compared to mice (166% of normal 17 ), and ninefold in AAVrh10.E12.A1AT-administered NHPs (14.2%) compared to mice (129% of normal 17 ). Conversely, others have seen larger differences when moving from mice to NHPs. McIntosh et al. observed a 49-fold change when translating expression of the rAAV8-HLP-codop-hFVIII-V3 vector from mice to rhesus macaques. 11 Studies with the AAV5 capsid for delivery of the hFVIII transgene have reported less than twofold reductions in expression from mice to primates. 24 However, this is likely due to the lower level of hFVIII expression seen in mice following AAV5 administration compared to other vector capsids. A dose of 2 × 1012 vector genomes (vg)/kg (approximately 4 × 1010 vg/mouse) of an AAV5-based vector produced 5% of normal hFVIII levels 24 compared to >100% of normal levels following injection of 1010 GC/mouse of AAVrh10 or AAVhu37 vectors. 17
As two vectors have been evaluated in clinical trials, it is now possible to translate dose and hFVIII expression from mice to NHPs and humans. 4,5 Administration i.v. of 6 × 1013 vg/kg of an AAV5-based vector in mice and humans resulted in steady-state hFVIII expression of 380% and 104% of normal, respectively, which is a 3.7-fold reduction. 4,24 Interestingly, a slightly lower dose in macaques (3.6 × 1013 vg/kg) resulted in 43% of normal hFVIII levels, which is similar to the recently reported median and mean values for subjects administered with 4 × 1013 vg/kg in an expanded clinical trial. In studies using the AAV-LK03 vector capsid for expression, similar doses in macaques and humans (2 × 1012 vg/kg and 1012 vg/kg, respectively) resulted in 30% of normal hFVIII levels in NHPs and an average of 11% of normal levels in two human subjects. 5 These relatively modest reductions in efficacy from macaques to humans following i.v. administration of similar vector doses highlight the importance of large-animal models in the translation of gene therapy applications into the clinic.
In addition to the technical challenges of vector development, immune responses that lead to the development of inhibitory antibodies to therapeutic FVIII in >30% of severe (<1% of normal FVIII activity) hemophilia A patients are problematic. By comparison, only 3% of patients with severe hemophilia B develop inhibitors. 26 –30 In the present study, the majority of macaques (18/20) developed anti-hFVIII antibodies within the first 30 weeks after vector administration, with median times to antibody generation of 8–10 weeks across groups. This is unsurprising, as the study was evaluating the expression of a foreign human protein in macaques. However, two macaques administered with AAVhu37.E03.TTR did not develop anti-hFVIII antibodies during the initial phase of the study (up to week 30 post vector administration).
In a previous study, the immunogenicity of the hFVIII transgene was also examined as a function of the AAV capsid used for delivery in FVIII KO mice. 17 These studies focused primarily on clade E AAVs, including AAV8, AAVrh10, AAVhu37, and AAVrh64R1. Over the course of the study, the percentage of mice that generated anti-hFVIII antibodies ranged from 20% (AAV8 and AAV9) to 63% (AAVrh10), with antibodies occurring in 40% of mice administered with AAVhu37. Although the promoter used in the capsid comparison study in mice (E06.TTR) was different from the two used in this study, the relative propensity of mice to generate anti-hFVIII antibodies when administered with AAVrh10 or AAVhu37 capsids was surprisingly modeled in NHPs. Although it is generally believed that mice are far less immune responsive than NHPs, this was not the case in this study. In cynomolgus macaques, 100% of animals administered with AAVrh10 developed anti-hFVIII antibodies by week 8–10 compared to 60% of macaques administered with AAVhu37. Time-to-event analysis also showed that anti-hFVIII antibody generation occurred earlier in the AAVrh10-administered groups, regardless of the vector expression cassette. Although there was not a statistically significant difference in time-to-antibody generation between the capsids, the trend toward reduced inhibitors following AAVhu37 administration could be a function of increased liver transduction with AAVhu37, leading to enhanced tolerance of the hFVIII transgene, a process that has been previously described for other transgenes. 31 –39 Interestingly, AAVhu37 also has a lower prevalence of naturally occurring NAbs in the human population compared to other capsids evaluated for treatment of hemophilia A (AAVrh10, AAV5, and AAV-LK03) or AAV8. Therefore, use of the AAVhu37 capsid would be preferable for this application.
When anti-hFVIII antibodies developed in the macaques, a peak in PTT levels was also observed for 11 animals. Only six occasions were observed where PTT values were >45 s (outside the range of normal variation), and these did not correlate with any observed coagulopathies. The exception was macaque M11657, which had a severe bleeding event during the initial phase of the study due to an injury to the base of the tail. In this animal, the PTT values remained >28 s for the duration of the study. Changes in PTT could indicate cross-reaction of the anti-human FVIII antibodies to endogenous macaque FVIII. However, this is unlikely to be the case in the majority of the animals, as PTT values returned to within normal limits 2–4 weeks after the first peak levels, indicating a transient effect and not a prolonged immune response to ongoing hFVIII production from the gene therapy vectors.
In conclusion, AAVhu37 has emerged as the least immunogenic capsid in both mice and NHPs, suggesting that capsid choice can play a substantial role in directing the development of transgene-specific B-cell responses in these animals. AAVhu37 was isolated from human samples and is a member of the clade E family, along with AAV8 and AAVrh10, which were isolated from rhesus macaques. This study expanded on the initial analyses of AAVhu37 for transgene expression and immunogenicity in both mice and rhesus macaques. 40,41 Additionally, AAVhu37 was evaluated for manufacturability, and it was shown that the binding affinity of AAVhu37 to AVB resin is suitable for the scalable purification of this capsid. 42 In the present study, yields for vector lots of AAVhu37 were significantly higher than AAVrh10 (data not shown; p < 0.05, Student's t-test). Together, these promising data suggest that the AAVhu37-based gene therapy approach for the treatment of hemophilia A has the potential to move into the clinic, with levels of expression following i.v. administration of 1.2 × 1013 GC/kg being sufficient to allow modification of a severe hemophilia A phenotype, potentially removing the requirement for recombinant FVIII infusions.
Footnotes
Acknowledgments
We would like to thank Mingyao Li and Yan Che for statistical analysis, and Leah Makaron and Mason E. Garnett for invaluable technical assistance. This research was supported by Ultragenyx/Bayer.
Author Disclosure
J.M.W. is an advisor to, holds equity in, and has a sponsored research agreement with REGENXBIO and Scout Bio. He has a sponsored research agreement with Ultragenyx, Biogen, and Janssen, which are licensees of Penn technology. He also holds equity in Solid Bio, and he is an inventor on patents that have been licensed to various biopharmaceutical companies. J.A.G, L.W., and J.M.W. are inventors on patents licensed to various biopharmaceutical companies. S.W. is a shareholder in Ultragenyx. No competing financial interests exist for the remaining authors.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.