
Abstract
Oncolytic virotherapy is a promising strategy for the treatment of cancer. Influenza A virus has shown potential as an oncolytic agent. In this study, a recombinant PR8 influenza viral vector, called delNS1-GM-CSF, was generated with a partial deletion in NS and the granulocyte-macrophage colony-stimulating factor (GM-CSF) coding sequence inserted into the influenza nonstructural protein 1 gene. The morphological characteristics of delNS1-GM-CSF were examined. The delNS1-GM-CSF virus replicated well in various cell lines, including MDCK, A549, SMCC7721, and HepG2 cells. Moreover, selective cytotoxicity of the virus was observed in various hepatocellular carcinoma (HCC) cell lines, while no effect was demonstrated in the normal liver cell line LO2, as indicated by 3-(4,5-dimethylthiazole-2-yl)-2,5-diphenyltetrazolium bromide and crystal violet assays. Importantly, using a model based on the growth of HepG2 cells as a xenograft in nude mice, it was found that a reassortant delNS1-GM-CSF virus inhibited tumor growth significantly following intratumoral injection in a dose-dependent manner. Ex vivo results showed that the tumor inhibition efficacy of delNS1-GM-CSF was observed in HCC clinical samples. Taken together, these results are the first to demonstrate that influenza A viruses may have potential as oncolytic virotherapeutic agents against HCC.
Introduction
Hepatocellular carcinoma (HCC) is the sixth most common malignant tumor and second leading cause of cancer-related deaths. 1,2 Although great progress has been made in HCC therapy, including surgical resection, liver transplantation, radiotherapy, hyperthermia, and molecular target therapy, the 5-year survival rate for HCC patients remains poor. 2,3 Therefore, innovative therapies for efficient HCC treatment are needed.
Oncolytic viruses (OVs) are promising agents for the treatment of cancer. 4 –6 OVs that can be genetically engineered or natural viruses can replicate in and kill malignant cells without harming normal tissues. 7 In most cancer cells, the mechanisms that prevent a viral infection are defective. 8 Thus, most viruses can easily replicate in cancer cells but not in normal cells. The massive replication of viruses in tumor cells may directly lead to cell lysis and induce systemic antitumor immunity. 9 OVs can kill infected cancer cells through conventional means of cell death—apoptosis, autophagy, necrosis, and pyroptosis. In addition to killing infected cells, OVs can destroy tumor blood vessels and amplify specific antitumor immune responses. 10 Several OVs have been investigated in clinical trials, including adenovirus, herpes simplex virus, measles virus, vesicular stomatitis virus, and vaccinia virus. 11 –16 Strikingly, in October 2015, a modified herpes simplex virus type 1, termed talimogene laherparepvec (T-VEC; Amgen, Thousand Oaks, CA), was the first OV therapy to be approved by the Food and Drug Administration for the treatment of unresectable melanomas. 17 Indeed, several OVs have been tested in HCC patients in Phase I or II clinical trials. 18 –20 However, not all cancer patients with HCC had a favorable response. Thus, novel OVs that show good performance in vitro and in vivo must be studied.
Influenza A virus (IAV) is a serious public-health threat that has long been studied as an OV. As a member of the family Orthomyxoviridae, IAV is an RNA virus that is incapable of integrating into the host genome. NS1 is responsible for interferon antagonism and is critical for viral replication in interferon-competent cells. Recently, Peter Palese's group reported that a recombinant influenza virus expressing a single-chain antibody antagonizing the immune checkpoint CTLA4 (IAV-CTLA4) could delay tumor growth in a mouse melanoma model. 21 In addition, Egorov et al. 22 found that the introduction of the attenuation marker “elastase cleavage site” in flu viral hemagglutinin allowed for safe, effective OV therapy. Previously, the recombinant influenza viruses rFLU/RSV/F + G, rFLU/HAdV, and rgFLU-HCVCE1E2 were successfully produced, and it was found that they elicited robust protective immune responses against respiratory syncytial virus, human adenovirus, and hepatitis C virus, respectively. 23 –25 Granulocyte-macrophage colony-stimulating factor (GM-CSF) is a monomeric glycoprotein secreted by macrophages, T cells, mast cells, natural killer cells, endothelial cells, and fibroblasts that functions as a cytokine. 26 GM-CSF stimulates stem cells to produce granulocytes (neutrophils, eosinophils, and basophils) and monocytes. In the last two decades, GM-CSF has been an effective therapy to treat tumor patients. 27 The present study generated a replication-incompetent IAV, termed delNS1-GM-CSF, which utilizes truncated NS viral segments to express GM-CSF. The reassortant virus replicated well in various cell lines. The oncolytic efficacy of delNS1-GM-CSF was investigated in vitro and in a mouse model of HCC.
Methods
Cell lines and viruses
The human hepatoma cell lines HepG2, SMMC7721, MHCC97L, Huh7.5, as well as the normal human liver cell line L02, were purchased from the American Type Culture Collection (Manassas, VA), cultured in Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum, and incubated at 37°C with 5% CO2. The influenza virus A/PR/8/34 (abbreviated PR8) used in this study was grown in the allantoic cavities of 10-day-old specific pathogen-free (SPF) chicken embryos (Beijing Laboratory Animal Center, Beijing, China).
Plasmid-based system for the rescue of the recombinant influenza virus
Reverse genetics was used to generate the reassortant influenza virus delNS1-GM-CSF. The delNS1-GM-CSF virus has a deletion in the NS gene segment, resulting in the expression of a truncated, non-functional NS1 protein. Meanwhile, the foreign GM-CSF gene was integrated into the NS segment. The pNS1/GM-CSF plasmid and remaining seven plasmids (pHW-PB2, pHW-PB1, pHW-PA, pHW-HA, pHW-NP, pHW-NA, and pHW-M) derived from influenza PR8 were co-transfected into MDCK and COS-1 cells (at a ratio of 1:2) using Effectene Transfection Reagent (Qiagen, Hilden, Germany) according to the manufacturercturering to the manufacturerufacturered, non-functional NS1 protein. Meanwhile, thent delNS1-GM-CSF virus supernatant and cell lysates were collected, and the 50% tissue culture infectious dose (TCID50) in MDCK cells was used to determine the virus titer.
Electron microscopy
The recombinant delNS1-GM-CSF virus was characterized by electron microscopy. For morphology and size determinations, negative staining of concentrated, purified delNS1-GM-CSF was conducted, followed by visualization by transmission electron microscopy.
Viral growth curves
To examine the viral growth kinetics, A549, MDCK, SMCC7721, and HepG2 cells were infected with the delNS1-GM-CSF virus at a multiplicity of infection (MOI) of 0.01, and the plates were incubated at 37°C with 5% CO2. Cell culture supernatants were harvested at 12, 24, 48, 72, and 96 h post infection, and the viral titers were assessed as the logTCID50/mL.
Cell viability assays
HepG2, SMMC7721, MHCC97L, Huh7.5, and L02 cells were seeded on 96-well plates at 1.5 × 104 cells/well, and the delNS1-GM-CSF virus at various MOIs (0.1, 1, and 3) was added. On days 2, 3, and 4 after infection, cell viability was determined by the 3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide (MTT) assay. MTT was added to the culture medium for 3 h at 37°C. The medium in each well was then aspirated, and the precipitates were dissolved in dimethyl sulfoxide. The absorbance was read at 570 nm using a microplate reader.
Cytopathic effect assay
HepG2, SMMC7721, MHCC97L, Huh7.5, and L02 cells were grown to subconfluence and were infected with the delNS1-GM-CSF virus at various MOIs (0.1, 1, and 3). Three days after infection, 2% crystal violet in 20% methanol was added to the cells for 15 min. Next, the cells were washed with distilled water and photographed under a microscope.
Animal experiments
Six-week-old female BALB/c nude mice were subcutaneously injected with 2 × 106 HepG2 cells in a volume of 100 μL into the right flank. When the tumors reached a volume of 80–150 mm 3 , after 10–14 days, the mice were randomly divided into three groups (n = 8 in each group). One group received a high-dose intratumoral inoculation of 2 × 109 plaque forming units (PFU) of delNS1-GM-CSF in a volume of 100 μL, one group received a low-dose intratumoral inoculation of 2 × 106 PFU of delNS1-GM-CSF in a volume of 100 μL, and one group received 100 μL of phosphate-buffered saline (PBS) over a successive 5-day period. Single-site injection was used in this experiment. The overall physical condition and behavior of the mice after the first injection were monitored every 2 days, and measurements of tumor size were taken 14 days following the initial injection until 42 days. Caliper measurements of tumor sizes were taken at regular intervals throughout the experiment, and the length (l) and width (w) were recorded to calculate the tumor volume (V) using the formula (l × w 2 )/2. At 0, 5, 10, 15, and 20 days after the first delNS1-GM-CSF administration, the mice were sacrificed, and the tumors were snap-frozen for RNA extraction and IAV-specific real-time quantitative reverse transcription polymerase chain reaction (qRT-PCR). All experimental protocols were approved by the Institutional Animal Care and Use Committee and Ethics Committee of Beijing 302 Hospital. All facilities were accredited by the Animal Care and Ethics Committee of Beijing 302 Hospital.
qRT-PCR
Total RNA was obtained from the tumor tissues of mice using TRIzol Reagent (Invitrogen, Carlsbad, CA). cDNA was generated by reverse transcription using PrimeScript RT Master Mix (Takara, Shiga, Japan). The primer pairs for IAV M (forward: GCAAATGGTGCAGGCAATGA; reverse: ATCTGCACCCCCATTCGTTT) and GAPDH (forward: GGTGGTCTCCTCTGACTTCAACA; reverse: GTTGCTGTAGCCAAATTCGTTGT) were designed and synthesized by Invitrogen. qRT-PCR was performed in triplicate wells of a 96-well reaction plate using an ABI 7500 PCR System (Thermo Fisher Scientific, Waltham, MA).
Enzyme-linked immunosorbent assay
GM-CSF levels were measured using an enzyme-linked immunosorbent assay kit (Dakewe, Beijing, China).
Ex vivo model
Sixty-three primary fresh cancer tissue specimens from HCC patients were dissected into small volumes and cultured. Thirty-six hours later, delNS1-GM-CSF viruses were added to the culture system. The MTT assay was used to detect cell viability with a tumor histoculture endpoint staining computer image analysis (TECIA) system 96 h later. This study was approved by the Ethics Committee of Beijing 302 Hospital. Informed consent was obtained from all human participants.
Statistics
GraphPad Prism v5 was used for all data analyses (GraphPad Software, Inc., San Diego, CA). Groups were compared using analysis of variance. A p-value of <0.05 was considered to indicate statistical significance.
Results
Construction and characteristics of the recombinant delNS1-GM-CSF virus
The GM-CSF gene was inserted into the coding region of the PR8 NS segment, as illustrated in Fig. 1A. The rescued recombinant virus was sequenced completely and was the same as that of the plasmid used, as expected. Using reverse genetics technology, the recombinant influenza virus delNS1-GM-CSF, which expressed the GM-CSF gene, was recovered, along with the remaining gene segments from the PR8 virus. The hemagglutinin titers of delNS1-GM-CSF were 1:256–512 for the first passage in the allantoic fluid of the eggs. After concentration and purification, the recombinant delNS1-GM-CSF viral particles examined by electron microscopy were >80% viral spherical structures, 60–120 nm in size, with a characteristic lipid membrane bilayer on their outer surfaces (Fig. 1B). The delNS1-GM-CSF viruses were similar to wild-type influenza virus in both morphology and size. Moreover, the antigenic characteristics of delNS1-GM-CSF were stable for at least 1 year at −80°C (data not shown).
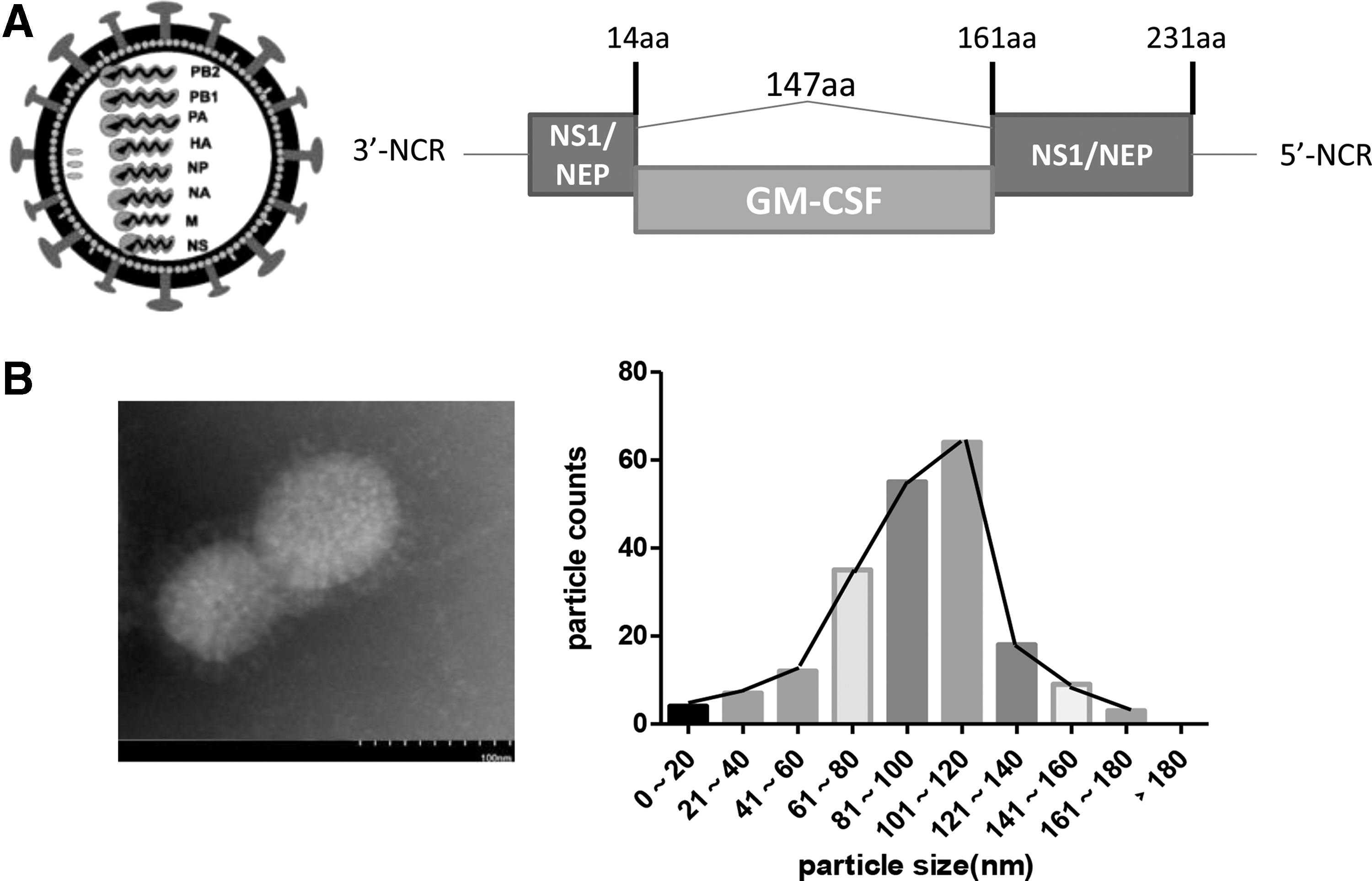
Construction and characterization of delNS1-GM-CSF.
To assess the viral growth kinetics further, MDCK, A549, SMCC7721, and Huh7.5 cells were infected with delNS1-GM-CSF viruses, and titers were determined. The culture medium was harvested at the indicated times post infection. As shown in Fig. 2, delNS1-GM-CSF viruses were replicated to titers of approximately 104 TCID50/mL in MDCK, A549, SMCC7721, and Huh7.5 cells after 12 h. The viral titers increased over time until the peak of 106 TCID50/mL was reached at 72 h. Next, a decreased tendency was observed at 96 h. Similarly, wild-type PR8 virus exhibited the same virus growth kinetics as the delNS1-GM-CSF viruses in these cell lines (Fig. 2).
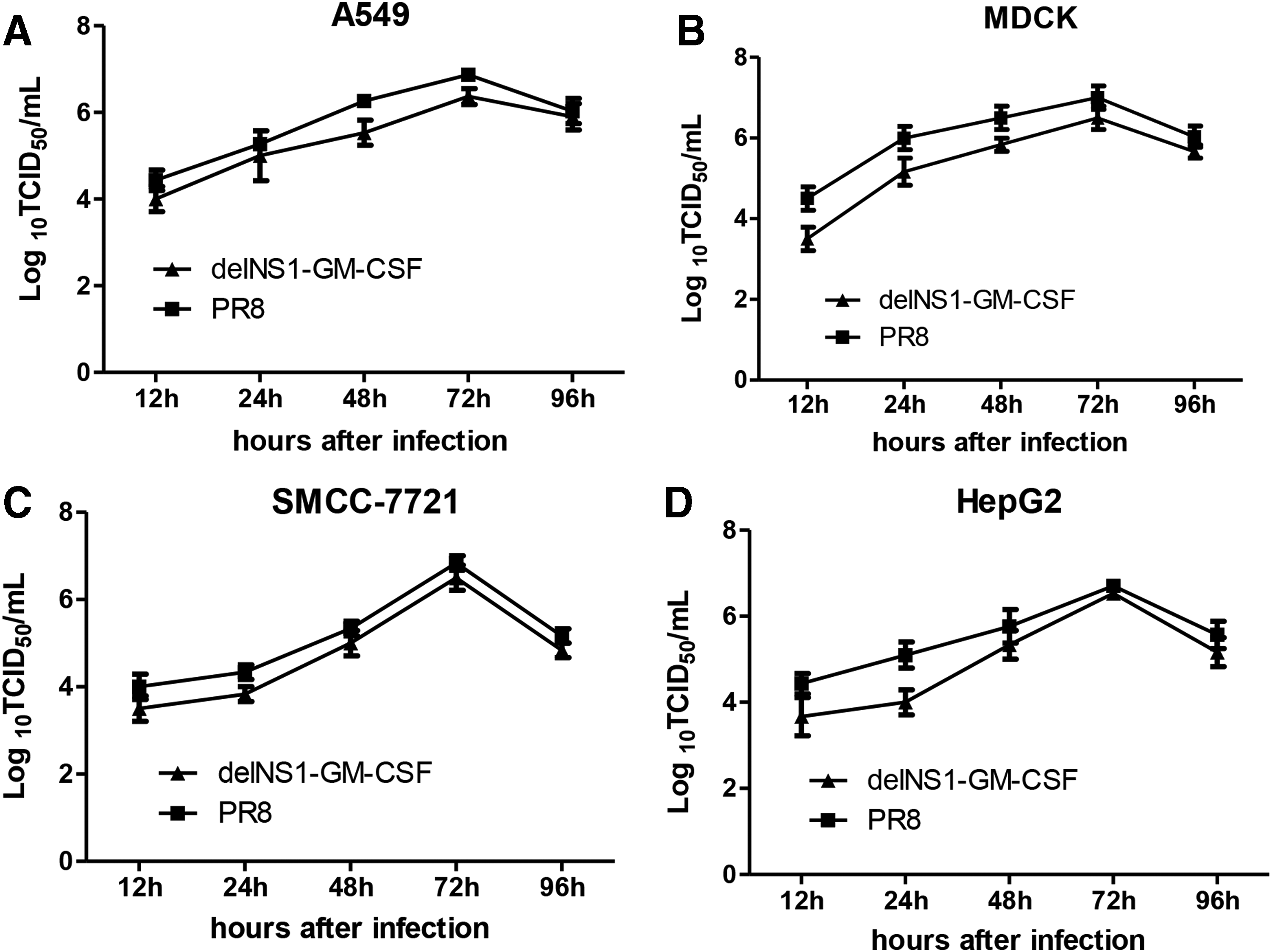
Virus growth kinetics of delNS1-GM-CSF in various cell lines. A549 (
Selective cytotoxicity of delNS1-GM-CSF in various HCC cell lines
To determine the selective cytotoxic effects of delNS1-GM-CSF, four HCC cell lines (SMMC-7721, HepG2, MHCC97L, and Huh7.5) and normal liver L02 cells were treated with different MOIs ranging from 0.1 to 3, and the cell viability was measured 48, 72, or 96 h post infection by the MTT assay. As shown in Fig. 3, delNS1-GM-CSF at a MOI of 1 or 3 significantly decreased HCC cell viability compared to that at a MOI of 0.1 at 48 h post infection (Fig. 3A). However, no severe side effect was induced by delNS1-GM-CSF infection at a MOI of 0.1, 1, or 3 for 96 h in the normal liver cell line LO2 assayed by the same methods. To identify the oncolytic ability of delNS1-GM-CSF further, crystal violet staining was performed. Similar to the MTT results, delNS1-GM-CSF infection at a MOI of 1 or 3 induced significant cytopathic effects in SMMC-7721, HepG2, MHCC97L, and Huh7.5 cells at 48 h post infection compared to uninfected control cells, while little cytotoxicity was observed in LO2 cells (Fig. 3F). These results suggest that delNS1-GM-CSF selectively killed various HCC cell lines in a time- and dose-dependent manner.
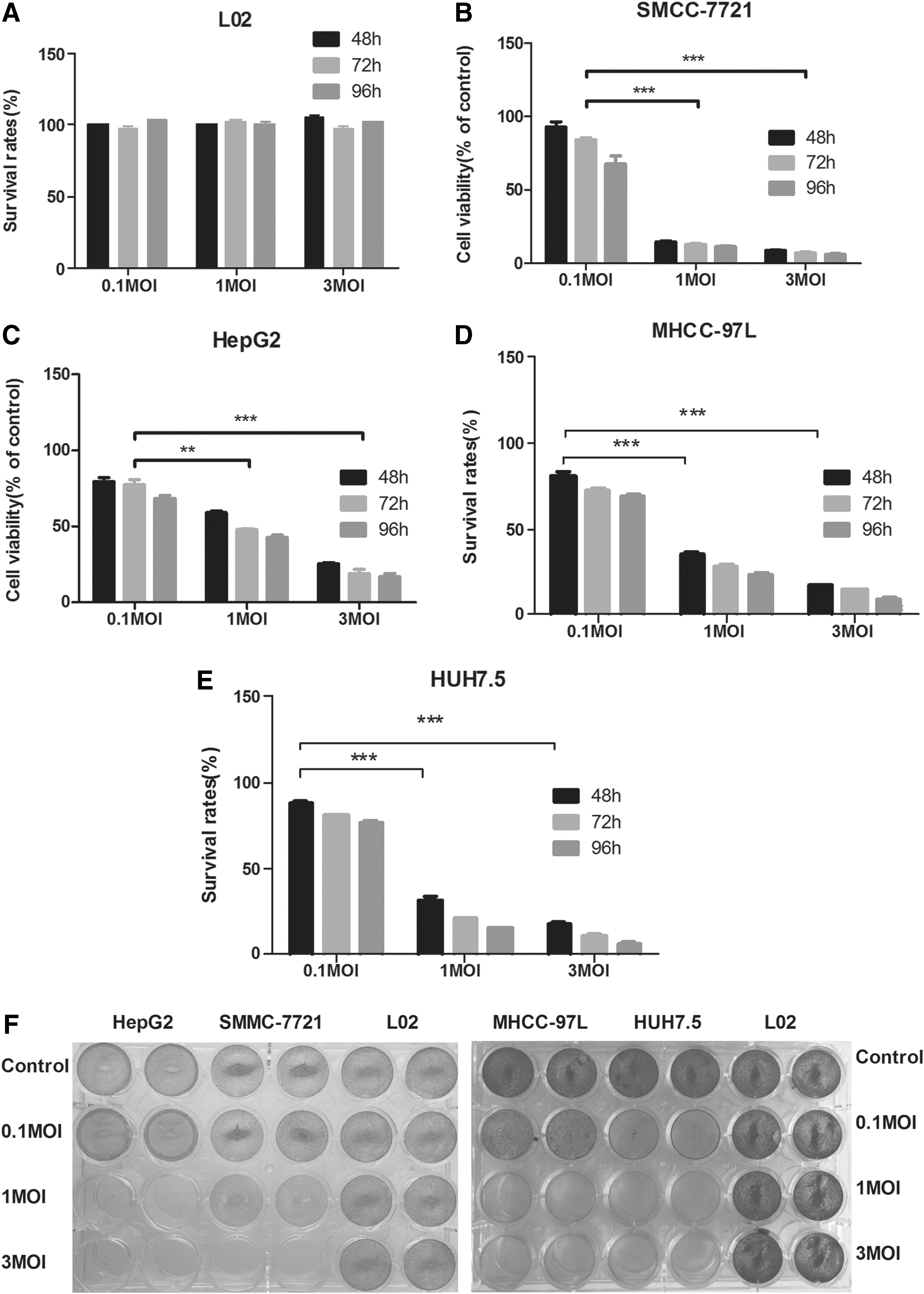
DelNS1-GM-CSF showed stronger cytotoxicity in hepatoma cells without severe side effects in normal liver cells. LO2 (
Oncolytic effects of delNS1-GM-CSF in vivo and ex vivo
To determine the in vivo oncolytic efficacy of delNS1-GM-CSF, a subcutaneous HepG2 xenograft model was established in BALB/c nude mice. When the tumor volume reached 80–150 mm 3 , mice were randomized to receive successively five high or low doses of delNS1-GM-CSF by intratumoral injection, spaced 24 h apart, and tumor growth was measured every 2 days for 42 days after the first injection (Fig. 4A). The tumors grew rapidly and exceeded a mean of 2,800 mm 3 within a month in the PBS group. Tumor growth in the delNS1-GM-CSF groups was significantly inhibited. It is worth noting that nearly 74% of the tumor burden was inhibited in the delNS1-GM-CSF group compared to that in the PBS group at the end of the fourth week, with a mean volume of 528 mm 3 (Fig. 4B). At the end of the experiments, it was observed that the tumor volumes in the delNS1-GM-CSF group were substantially smaller than those in the control group. The mean tumor weights of the high-dose group were significantly reduced by 75% and those of the low-dose group were reduced by 38% compared to those of the PBS group (Fig. 4C). Furthermore, the viral titers of delNS1-GM-CSF in the tumor were measured by qRT-PCR at the indicated time point after initial injection. Viral replication of delNS1-GM-CSF peaked at 5 days after the first delNS1-GM-CSF administration, with a mean of 4.5 × 103 copies/ng of total RNA. Next, delNS1-GM-CSF viral titers were decreased gradually at 10 and 15 days and were undetectable at 20 days (Fig. 4D). It was also found that no viral titers of delNS1-GM-CSF were detected in other organs of inoculated mice, including the heart, liver, spleen, lung, and kidney (data not shown). In addition, serum HI antibodies against IAV viruses were examined at 1 and 3 weeks post first injection. Results displayed that HI titers were increased rapidly at 3 weeks, with a mean of 1:300 (Fig. 4E). Next, the GM-CSF concentration was tested in HepG2 cell culture supernatant at different time points, and it was found that GM-CSF concentrations increased gradually from 24 to 96 h, with a mean of 650 pg/mL at 96 h (Fig. 4F). Meanwhile, GM-CSF concentrations in tumor tissue homogenate of nude mice were investigated. Results revealed that GM-CSF concentrations achieved about 2,600 pg/mL at 3 weeks post first injection (Fig. 4G). Finally, the tumor inhibition efficacy of delNS1-GM-CSF was detected in HCC clinical samples. Ex vivo results showed that the delNS1-GM-CSF virus significantly promoted liver tumor cell death (inhibition rate 61%; Fig. 4E). These data show that delNS1-GM-CSF treatment reduced the growth of HCC tumors in vivo in a dose-dependent manner and in primary HCC patient tissues.

Oncolytic efficacy of the delNS1-GM-CSF virus in vivo and ex vivo.
Discussion
Over the past few decades, increasing evidence has shown that OVs can cure cancer. 28,29 An IAV with deletion of the NS gene segment exhibited oncolytic activity in cells and animal models of prostate cancer, colon carcinoma, and melanoma. Replication of the delNS1 virus can be inhibited through PKR activation, which is always blocked in tumor cells. Thus, the delNS1 virus can replicate in those tumor cells. 30 –32 Herein, a recombinant influenza virus, delNS1-GM-CSF, encoding GM-CSF in the NS gene segment was produced using reverse genetics, and the antitumor activity of delNS1-GM-CSF was investigated in vitro and in vivo. The delNS1-GM-CSF virus replicated effectively in MDCK, A549, 7721, and HepG2 cells, similar to wild-type PR8 virus. Selective cytotoxicity of delNS1-GM-CSF was observed in a panel of human HCC cell lines. Next, the therapeutic effects of the delNS1-GM-CSF virus were investigated in a subcutaneous HepG2 xenograft model, and it was found that delNS1-GM-CSF administered intratumorally caused significant inhibition of tumor growth in HCC models compared to a mock-inoculated tumor group. In addition, delNS1-GM-CSF administered intravenously reduced tumor growth in this animal model (data not shown). In an ex vivo experiment, the infiltrating T cells (CD3+) were also analyzed in HCC tumors by immunohistological staining, and it was found that there is no obvious correlation between CD3+ T cells and the inhibition rate (data not shown). Among them, many cases (inhibition rate 61%) did not respond to delNS1-GM-CSF treatment. The likely reason is the high cutoff value of the inhibition rate (10%) and the individual differences of each HCC patient. Of course, whether the antitumor activity of delNS1-GM-CSF is universal to other tumors warrants further investigations in other mouse xenograft models of subcutaneous, orthotopic, and peritoneal tumor metastasis in mice as well as in humans.
The GM-CSF gene was introduced into the oncolytic flu vector to form a dual-regulated gene-viral system. The production of progeny infectious virus particles was observed in A549, MDCK, 7721, and HepG2 cells. DelNS1-GM-CSF could effectively replicate in these cells. Effective delivery to tumor tissues and rapid growth within tumor locations are critical factors for oncolytic virotherapy. 33 It was demonstrated that intratumoral administration of delNS1-GM-CSF is an effective strategy to deliver a virus to tumors. DelNS1-GM-CSF thrives mostly within tumor tissues, indicating that delNS1-GM-CSF has high tumor tropism. Moreover, all treated mice remained asymptomatic. In addition, BALB/c nu/nu mice were inoculated intrahepatically with delNS1-GM-CSF (2 × 109 PFU) and were monitored daily for clinical manifestations for 7 days. All mock-inoculated and delNS1-GM-CSF-inoculated mice survived without any detectable abnormalities (data not shown). Thus, it is speculated that delNS1-GM-CSF is safe for animals. However, additional experiments performed in syngeneic HCC models are needed to clarify the tumor specificity and mechanism of action of cancer cell oncolysis mediated by delNS1-GM-CSF.
In summary, a reassortant influenza virus expressing GM-CSF was used as an antitumor agent. The present study describes the generation of a GM-CSF-armed oncolytic delNS1-GM-CSF virus and shows that delNS1-GM-CSF exerted oncolytic effects in human HCC cells, mouse xenograft models, and primary HCC tumor tissues. These results warrant further investigation of delNS1-GM-CSF as an oncolytic IAV that combines oncolytic with enhanced immunostimulatory properties alone and in combination with cancer antibody drugs, including PD1/PDL1 and CTLA4. The findings demonstrate that oncolytic delNS1-GM-CSF virus may serve as a novel treatment for patients with HCC.
Footnotes
Acknowledgments
This work was supported in part by the funding from the National High Technology Research and Development Program of China (2015AA020924), Natural Science Foundation of Beijing Municipality (7162185).
Author Disclosure
No competing financial interests exist.