
Abstract
The long non-coding RNA HOXA-AS2 has been found to be an oncogene in several types of human malignant tumors. However, its role in regulating the occurrence and development of papillary thyroid cancer (PTC) is still unclear. The present study investigated the function and mechanism(s) of HOXA-AS2 in PTC progression. Using quantitative real-time polymerase chain reaction, HOXA-AS2 was found to be differentially expressed in PTC tissues and cell lines. Kaplan–Meier analysis indicated that the overall survival rate of patients with higher levels of HOXA-AS2 was lower than those with relatively lower levels. Loss-of-function assays revealed that HOXA-AS2 knockdown inhibited PTC progression by inhibiting cellular proliferation, migration, and invasion and accelerating apoptosis. Mechanistically, loss-of-function assays showed a positive correlation between HOXA3 and HOXA-AS2 expression. Subcellular fractionation assay results revealed abundant HOXA-AS2 expression in the cytoplasm of PTC cells. Additionally, FOXD2-AS1 was found to upregulate HOXA3 expression by binding to miR-15a-5p. Finally, rescue assays demonstrated the overall function of the HOXA-AS2/miR-15a-5p/HOXA3 axis in PTC progression. These findings will significantly contribute to further research and the development of more efficient treatments for thyroid cancer in the future.
Introduction
Thyroid cancer is a highly prevalent cancer resulting from complex endocrine dyscrasia that has a high occurrence and mortality. 1 –3 The increased incidence of thyroid cancer over the years has threatened human health. 4,5 Although some popular treatments, such as radiation, chemotherapy, and surgery, have been widely applied to treat thyroid cancer, the prognosis remains unfavorable. 6,7 Therefore, development of novel therapeutic methods for this cancer is urgently needed. Thyroid cancer is commonly divided into four subtypes: papillary thyroid carcinoma (PTC; most common), anaplastic thyroid carcinoma, follicular carcinoma, and medullary thyroid carcinoma. 8 The present study focused on molecular mechanisms contributing to the progression of PTC.
Long non-coding RNAs (lncRNAs) are >200 nt in length and do not encode proteins. 9 LncRNAs are crucial modulators in multiple biological processes, such as cellular apoptosis, invasion, and migration, 10 –12 and their ectopic expression can promote carcinogenesis. 13 –17 The lncRNA HOXA-AS2 has been reported to be an oncogene in several types of human cancer. 18 –21 However, it is unclear whether it functions as a regulator in PTC. LncRNAs can also exert their function in human tumors by competitively sharing microRNAs (miRNAs or miRs) with mRNAs. MiRNAs are short non-coding RNAs between 19 and 22 nt in length 22 that can participate in competitive endogenous RNA (ceRNA) pathways. 23 –26 According to previous reports, some lncRNAs function in human malignant tumors by acting as ceRNAs. 27 –30 Therefore, the present study investigated the function and mechanism(s) of HOXA-AS2 in the progression of PTC. In addition, the study also examined whether HOXA-AS2 acts as a ceRNA to regulate other nearby genes (e.g., HOXA3) by sponging miRNA, thereby modulating downstream signaling pathways (e.g., Wnt/β-catenin).
Methods
Collection of tissue samples
Approval from the Ethics Committee of Navy General Hospital (Beijing, P.R. China), as well as written informed consent from all included patients, was obtained prior to study onset. Matched PTC (n = 68) and normal (n = 68) tissues were obtained from PTC patients following surgical resection at Navy General Hospital. All tissue samples were directly frozen and stored in liquid nitrogen until use. PTC was confirmed in tissue samples via pathological examination. The mean expression level of HOXA-AS2 in all PTC tissues was determined and used to further divide these samples into two subgroups based on high and low HOXA-AS2 expression.
Cell culture
One normal human thyroid epithelial (Nthy-ori3-1) and three human papillary thyroid cancer cell lines (K-1, TPC and BHP2-7) were obtained from American Type Culture Collection (Manassas, VA). All cell lines were cultured in RPMI-1640 medium supplemented with 10% fetal bovine serum (FBS, Gibco, Thermo Fisher Scientific, Inc., Waltham, MA, USA), 0.1 mM amino acids (L-Glutamine, Gibco), 1 mM sodium pyruvate and 1% penicillin-streptomycin (Sigma-Aldrich, St. Louis, MO). Cell culture was conducted in a humidified environment with 5% CO2 at 37°C.
Cell transfection
MiR-15a-5p mimics/inhibitors and respective controls were purchased from GenePharma (Shanghai, P.R. China). Small interfering RNAs (siRNAs) targeted to HOXA-AS2 (si-HOXA-AS2#1 and si-HOXA-AS2#2) or HOXA3 (si-HOXA3#1 and si-HOXA3#2) and control siRNA (si-NC) were also purchased from GenePharma. A pcDNA-HOXA3 plasmid was obtained from Genechem Biotech (Shanghai, P.R. China), and Lipofectamine 2000 (Invitrogen, Carlsbad, CA) was used for cellular transfections in accordance with the manufacturer's guidelines.
Quantitative real-time polymerase chain reaction
TRIzol reagent (Thermo Fisher Scientific, Waltham, MA) was used to extract total RNA from cells according to the manufacturer's instructions. The isolated RNA was used to synthesize cDNA using an All-in-One™ miRNA quantitative real-time polymerase chain reaction (qRT-PCR) detection kit (GeneCopoeia, FulenGen, P.R. China). A Prime Script™ RT reagent kit (Takara, Shiga, Japan) was used for reverse transcription, and qRT-PCR was repeated three times on a 96-well PCR plate. Each well (20 μl) contained 1 μL of cDNA 10 mM dNTPs and 1 × 20 μL of Platinum SYBR Green qPCR SuperMix UDG (Invitrogen). The primers sequences for HOXA-AS2 were listed as follows: GTCTGCGAAGGCCTAAAGGT (forward), GACTCCCTGATGTTCGCGTC (reverse). The primers sequences for miR-15a-5p were listed as follows: TAGCAGCACATAATGGTTTGTG (forward), CTCTACAGCTATATTGCCAGCCA (reverse). The primers sequences for HOXA3 were listed as follows: CATCCCTTGGGTCGTCTCTC (forward), TTCTCGGGGCAAATCTGGTC (reverse). The primers sequences for U6 were listed as follows: AGGAGACGGGAACGACAAAC (forward), GAAGAGGATTCGCTGACGGT (reverse). The primers sequences for GAPDH were listed as follows: GCTTTCTTTCCTTTCGCGCT (forward), TTTGCGGTGGAAATGTCCTT (reverse). MiRNA and mRNA expression levels were normalized to U6-small nuclear RNA and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) by the 2–ΔΔCt method. 31
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay
Cells were seeded onto 96-well culture plates (Corning Costar, Corning, NY) at a density of 2 × 103 cells per well and cultured for 24 h prior to transfection. Next, 20 μL of MTT solution (5 mg/mL; Sigma–Aldrich) was added and incubated in each well for 4 h at 37°C. After, the media was removed from each well, and intracellular formazan crystals were dissolved by adding 200 μL of dimethyl sulfoxide (Sigma–Aldrich). Cell viability was detected by measuring the absorbance at 490 nm on a Multiskan Ascent 354 microplate reader (Thermo Labsystems, Waltham, MA).
Flow cytometry
Following transfection, cells were collected and washed with phosphate-buffered saline before re-suspending 1 × 106 cells from each sample in binding buffer. Binding buffer contained phosphate buffer saline (PBS) and 1% FBS. Then, an Annexin V-fluorescein isothiocyanate/propidium iodide kit (BD Pharmingen, San Diego, CA) was used to measure cell viability via flow cytometry (BD FACS Aria; BD Biosciences, Franklin Lakes, NJ).
Transwell® assay
After transfection, PTC cell lines (1 × 104) were re-suspended in 200 μL of serum-free medium. For both invasion and migration assays, PTC cell lines were then seeded into the upper chambers of Transwell® plates (8 μm pore size; Corning Costar), which were coated with or without Matrigel (BD Biosciences). Medium containing 10% FBS was used as a chemoattractant and added to the bottom chamber of each well, and cells were incubated at 37°C with 5% CO2 for 24 or 48 h for migration and invasion assays, respectively. Afterward, cells present in the upper chamber were wiped out with cotton swabs, whereas cells that had adhered to the bottom chamber were fixed with methanol and stained with 0.1% crystal violet. Stained cells were imaged at 100 × magnification under a microscope (Olympus, Tokyo, Japan).
Luciferase reporter assay
The sequences for wild-type HOXA-AS2 (HOXA-AS2-WT) and HOXA3 (HOXA-WT), as well as mutant sequences (HOXA-AS2-MUT and HOXA3-MUT), containing the wild-type or mutant putative binding sites of miR-15a-5p were augmented and then cloned into pmIRGLO-control luciferase reporter vectors (Promega, Madison, WI). The reporter vectors with miR-15a-5p mimics and controls (miR-NC) were co-transfected into K-1 and BHP2-7 cells with Lipofectamine 2000 (Invitrogen) for 36 h. Because HOXA-AS2 were highly expressed in K-1 and BHP2-7 cells, we selected the two cells for subsequent analysis. Afterward, cells were collected and lysed, and luciferase activity was tested with a Dual-Luciferase® Reporter Assay System (Promega).
Subcellular fractionation assay
Cytoplasmic and nuclear fractions were extracted from PTC cell lines using NE-PER Nuclear and Cytoplasmic Extraction Reagents (Thermo Fisher Scientific). 1 × 107 K-1 and BHP2-7 cells were first washed twice in pre-chilled PBS and lysed in 500 μl of cell fractionations buffer containing NP40 (10 mM HEPES, 60 mM KCl, 1 mM EDTA, 0.075% (v/v) NP40, 1 mM DTT and 1 mM PMSF, pH 7.6). After incubated on ice for 3 minutes, lysates were centrifuged at 1500 g for 4 minutes. The nuclear and cytoplasmic cell fractions were separated. The cytoplasm supernatant was transferred into a fresh RNase-free tube. Nuclear fractions were cultured in 100 μl of nuclear lysis buffer containing 20 mM tris-HCl, 420 mM NaCl, 1.5 mM MgCl2, 0.2 mM EDTA, 1 mM PMSF and 25% (v/v) glycerol, pH 8.0. After incubation at 4°C for an hour, nuclear extracts were reaped through centrifugation at 12,000 g for 10 minutes. RNAs derived from the cytoplasmic or nuclear extracts were obtained using TRIzol (Life Technologies, Carlsbad, CA, USA). Followed by elution, isolated RNAs were analyzed by qRT-PCR. The levels of nucleus control (U6), cytoplasm control (GAPDH) and HOXA-AS2 were separately measured.
RNA-binding protein immunoprecipitation assay
For RNA-binding protein immunoprecipitation (RIP) assays, PTC cell lines were co-transfected with pcDNA-MS2, pcDNA-HOXA-AS2-MS2, and pBobi-MS2-GFP. The RIP assay was conducted utilizing an anti-green fluorescent protein antibody (Abcam, Cambridge, United Kingdom) and MagnaRIP RIP kit (Millipore, Bedford, MA) according to the manufacturer's instructions. Immunoglobulin G (Millipore) was used as a negative control (input group).
Western blot
Radioimmunoprecipitation assay buffer was used to extract all proteins from cells, and a NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific) was used to determine the concentration of proteins in each sample. To prepare protein samples, we treated cells thrice with 3 ml of pre-cooled 0.01M PBS. After removing PBS, cells were incubated with 1 ml RIPA lysis buffer on ice for half an hour. Thereafter, cells were centrifuged at 12000 rpm at 4°C for 5 minutes. At length, the supernatant was transferred into 0.5 min centrifugal tube, followed by preservation at −20°C. Proteins (50 μg/lane) were separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis and then transferred onto polyvinylidene fluoride membranes (Millipore). Membranes were blocked with 5% bovine serum albumin in Tris-buffered saline with 0.1% Tween 20 (TBST) for 2 h at room temperature prior to incubation with primary antibodies at 4°C overnight. Primary antibodies used were anti-HOXA3 (1:1,000; Santa Cruz Biotechnology, Santa Cruz, CA), anti-β-catenin (1:1,000; Cell Signaling Technology, Danvers, MA), anti-cyclin D1 (1:1,000; Cell Signaling Technology), anti-c-Myc (1:1,000; Cell Signaling Technology), and anti-GAPDH (1:5,000; Sigma–Aldrich). Then, membranes were washed in TBST and incubated with a horseradish peroxidase-tagged secondary antibody at 37°C for 1 h. Afterward, membranes were again washed with TBST, and protein bands of interest were detected by an enhanced chemiluminescence system (Pierce, Rockford, IL). Bands were quantified by ImageJ software (National Institutes of Health, Bethesda, MD).
In vivo assays
6-week-old female nude mice were bought from National Laboratory Animal Center (Beijing, China). Mice were first maintained in the SPF-grade pathogen-free animal laboratory equipped with laminar flow equipment and 12 h light-dark cycles. Nude mice were free to autoclaved food and water. All experiments with mice were conducted strictly in accordance with the Guide for the Care and Use of Laboratory Animals. In vivo assay was approved by the review board of the State key laboratory of environmental microorganism utilization and safety control at Shaoxing Hospital of China Medical University (Shaoxing, P.R. China). 5 × 104 K-1 cells were transfected with various plasmids or combinations. Cells were reaped and subcutaneously injected into the left flank of the nude mice using sterile 28-gauge needles. One week after injection, mice were randomly divided into two groups (n = 4 per group) when PTC tumors had reached approximately 3 mm 3 . Tumor volume was measured every fourth day. Four weeks after implantation, all mice were sacrificed. Tumors were excised and weighed for subsequent analysis. The tumor sizes were determined by measuring their length (l) and width (w) via a digital caliper. The tumor volumes (V) were calculated with the following equation: V = lw2/2. Finally, we obtained approximately 8 × 103 mm 3 PTC tumors.
Bioinformatics analysis
The University of California-Santa Cruz genome database (
Statistical analysis
All results are shown as means ± standard deviations from three independent experiments. Student's t-test was applied to measure the differences between two groups. One-way analysis of variance was performed to compare differences among more than two groups. Statistical analyses were completed using IBM SPSS Statistics for Windows v19.0 (IBM Corp., Armonk, NY). Statistically significant correlations between HOXA-AS2 and miR-15a-5p, as well as miR-15a-5p and HOXA3, were analyzed using Spearman's correlation analysis. Differences were considered statistically significant with a p-value of <0.05.
Results
Heightened expression of HOXA-AS2 is associated with a poor prognosis in PTC patients
qRT-PCR results showed HOXA-AS2 expression levels in PTC tissues and cell lines were obviously higher than that in matched normal tissues and control cells (Fig. 1A–C). Using the mean HOXA-AS2 expression value as a threshold, PTC samples were classified into high and low HOXA-AS2 expression groups. Next, the correlation between HOXA-AS2 expression and clinicopathologic features of PTC patients was analyzed. As presented in Table 1, dysregulation of HOXA-AS2 indicated advanced tumor-node-metastasis staging and lymph node metastasis. According to Kaplan–Meier analysis, patients with higher HOXA-AS2 expression had a lower survival rate than those with lower expression (p = 0.007; Fig. 1C). These findings indicate a potential oncogenic role for HOXA-AS2 in PTC.
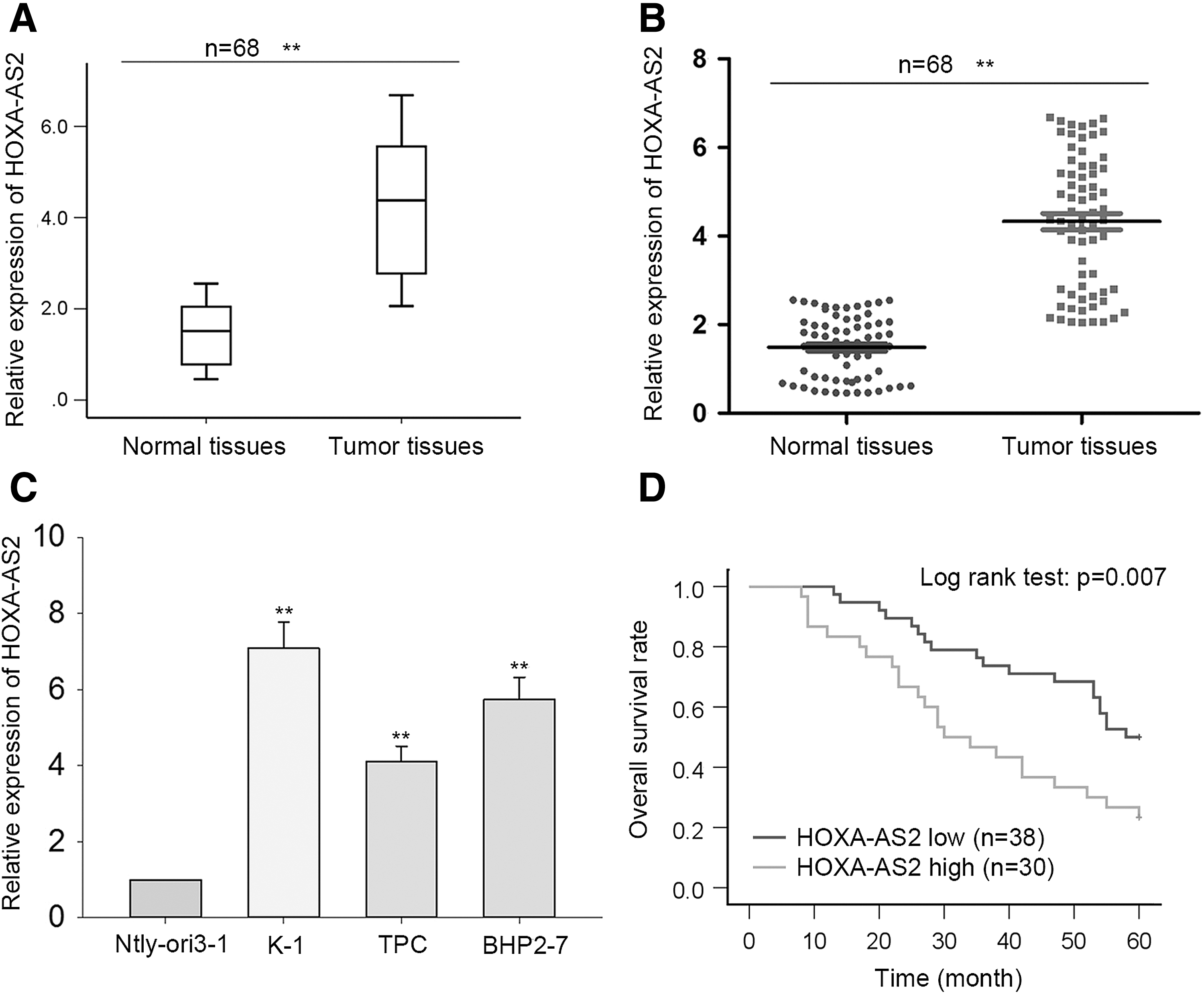
Higher expression of HOXA-AS2 is associated with a poor prognosis in papillary thyroid cancer (PTC) patients.
Correlation between HOXA-AS2 expression and clinical features of PTC patients (n = 68)
Low/high by the sample median. Pearson's chi-square test.
p < 0.05; ** p < 0.01; *** p < 0.001.
PTC, papillary thyroid cancer; TNM, tumor-node-metastasis.
HOXA-AS2 knockdown suppressed cell proliferation and accelerated apoptosis
In order to assess HOXA-AS2 function in PTC, K-1 and BHP2-7 cell lines were transfected with si-HOXA-AS2#1 and si-HOXA-AS2#2 or si-NC (Fig. 2A). MTT assay results revealed that the proliferation of cells transfected with either si-HOXA-AS2#1 or si-HOXA-AS2#2 was greatly suppressed (Fig. 2B). Moreover, flow cytometry results showed that knockdown of HOXA-AS2 remarkably enhanced the apoptosis rate of PTC cells (Fig. 2C).

Knockdown of HOXA-AS2 suppressed cell proliferation and accelerated apoptosis.
Knockdown of HOXA-AS2 inhibited migration and invasion of PTC cells
To probe into the effect HOXA-AS2 depletion on the migratory and invasive abilities of K-1 and BHP2-7 cells, we carried out Transwell® assays. Results revealed that HOXA-AS2 knockdown significantly reduced the number of both migratory and invasive cells, indicating the negative impact of HOXA-AS2 knockdown on PTC cell migration and invasion (Fig. 3A and B).

Silencing of HOXA-AS2 inhibited migration and invasion of PTC cells.
Knockdown of HOXA3 suppressed PTC cell proliferation, migration, and invasion but induced apoptosis
According to an online search of the University of California-Santa Cruz genome database (

Knockdown of HOXA3 suppressed cell proliferation, migration, and invasion but induced apoptosis.
HOXA-AS2 is a molecular sponge of miR-15a-5p
Previous studies have shown that lncRNAs can act as ceRNAs to upregulate mRNAs by sponging miRNAs.
32
–34
Based on the above findings, HOXA-AS2 was hypothesized to act as a ceRNA to upregulate HOXA3 in PTC. Subcellular fractionation assay results showed that the level of cytoplasmic HOXA-AS2 in PTC cells was greater than that in the nucleus (Fig. 5A), suggesting that HOXA-AS2 lncRNA regulates gene expression in PTC cells at the post-transcriptional level. Subsequently, 14 miRNAs that can bind HOXA3 were found using bioinformatics analysis software (miRanda, picTar, TargetScan, and PITA; Fig. 5B). A RIP assay was then used to determine whether these 14 miRNAs can also bind HOXA-AS2 in PTC cells. The results showed co-immunoprecipitation of HOXA-AS2 and miR-15a-5p from BHP2-7 and K-1 cell lines (Fig. 5C). Next, HOXA-AS2 and miR-15a-5p binding sites were predicted using DIANA tools (

HOXA-AS2 is a molecular sponge of miR-15a-5p.
HOXA-AS2 contributed to HOXA3 upregulation by competitively binding miR-15a-5p
The putative binding sequence between miR-15a-5p and HOXA3 was obtained from Target-Scan† (Fig. 6A) and confirmed by Dual-Luciferase® reporter assay. Co-transfection of HOXA3–WT with miR-15a-5p mimics reduced luciferase activity in PTC cells. However, activity was recovered by upregulation of HOXA-AS2 (Fig. 6B). Spearman's correlation analysis revealed a negative association between miR-15a-5p and HOXA3 expression (Fig. 6C). Moreover, qRT-PCR and Western blot analyses of PTC cells co-transfected with sh-HOXA-AS2 and miR-15a-5p inhibitors showed that decreases in HOXA3 mRNA and protein levels caused by sh-HOXA-AS2 could be recovered by miR-15a-5p inhibitors (Fig. 6D and E). These findings indicate that HOXA-AS2 positively modulates HOXA3 expression in PTC cells by competitively binding to miR-15a-5p.
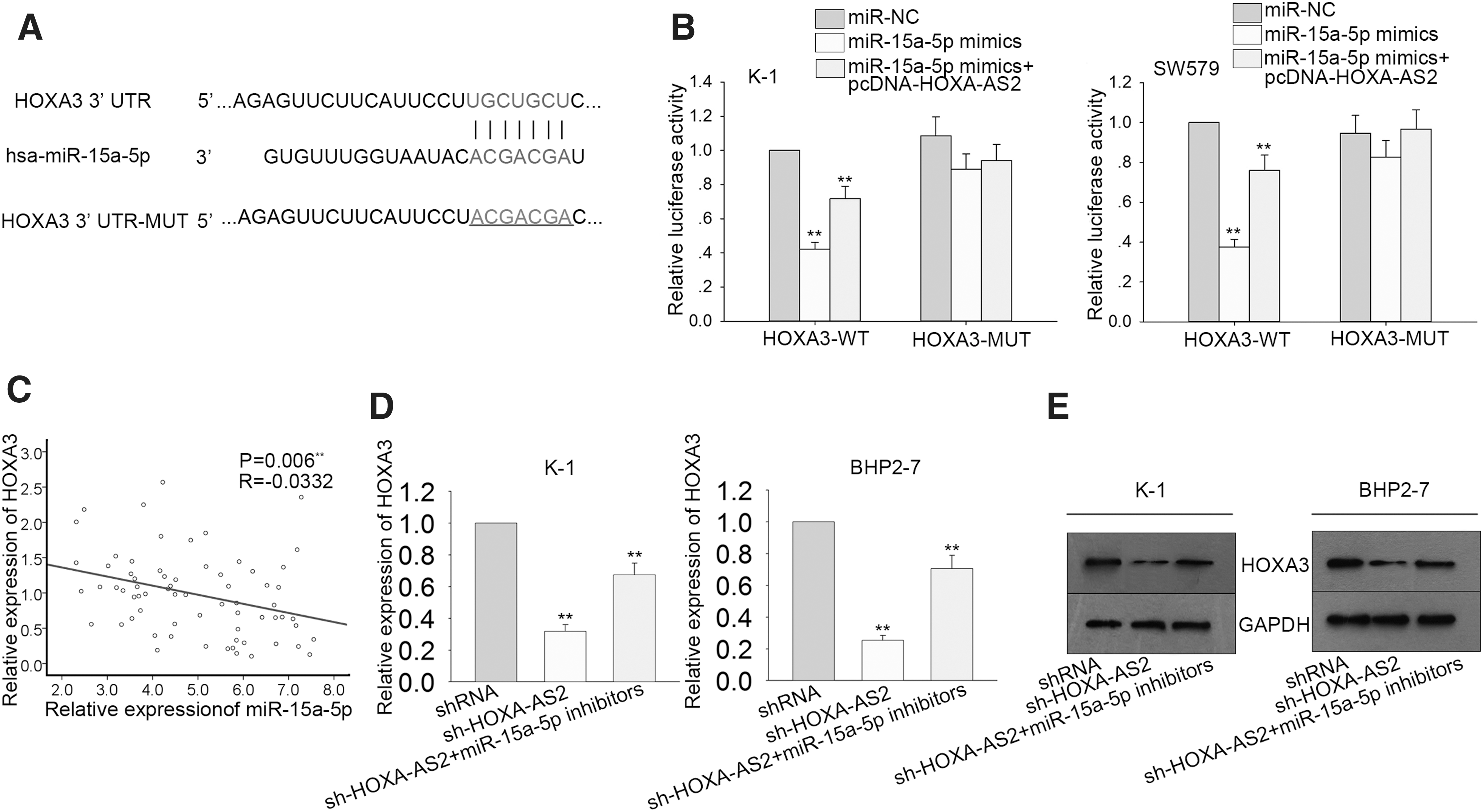
HOXA-AS2 contributed to the upregulation of HOXA3 by competitively binding miR-15a-5p.
HOXA-AS2/miR-15a-5p/HOXA3 axis regulates PTC progression
Rescue assays were used to demonstrate the role of HOXA-AS2/miR-15a-5p/HOXA3 axis in PTC progression. Results of MTT assays showed that HOXA3 and miR-15a-5p inhibitors partially reversed sh-HOXA-AS2-mediated cell proliferation in K-1 cell line (Fig. 7A). Moreover, the increased rate of apoptosis caused by HOXA-AS2 knockdown could be decreased by upregulation of HOXA3 or downregulation of miR-15a-5p (Fig. 7B). Furthermore, Transwell® assays revealed that migration and invasion activity suppressed by sh-HOXA-AS2 could be recovered by HOXA3 overexpression or miR-15a-5p inhibitors (Fig. 7C and D).
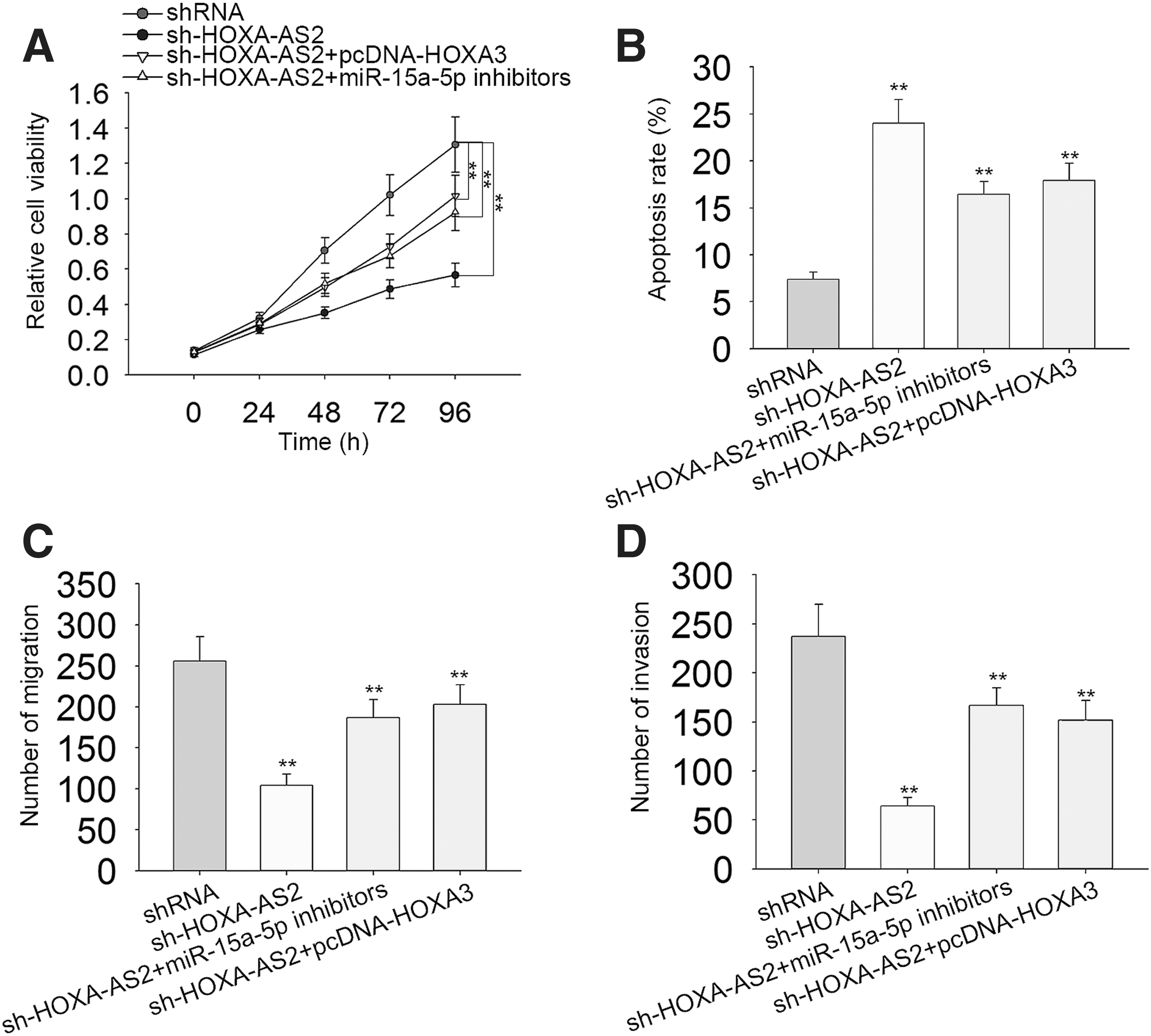
The HOXA-AS2/miR-15a-5p/HOXA3 axis regulates PTC progression.
HOXA-AS2/miR-15a-5p/HOXA3 axis regulates PTC tumor growth in vivo
In vivo experiments using mice were conducted to demonstrate the function of the HOXA-AS2/miR-15a-5p/HOXA3 axis further in PTC tumor growth. As illustrated in Fig. 8A, tumor growth was inhibited by HOXA-AS2 knockdown but promoted by miR-15a-5p knockdown or HOXA3 overexpression. Additionally, the growth inhibition caused by HOXA-AS2 knockdown was partly recovered by miR-15a-5p silencing or HOXA3 overexpression. Moreover, the decreased miR-15a-5p expression or increased HOXA3 expression significantly rescued the sh-HOXA-AS2-mediated reduced tumor volume and weight. These results illustrate regulation of PTC tumor growth in vivo by the HOXA-AS2/miR-15a-5p/HOXA3 axis.

The HOXA-AS2/miR-15a-5p/HOXA3 axis regulates PTC cell growth in vivo.
HOXA-AS2/miR-15a-5p/HOXA3 axis synergistically regulates Wnt/β-catenin signaling
According to previous studies, Wnt/β-catenin signaling plays a crucial role in lncRNA-mediated tumor progression. Based on the above findings, the HOXA-AS2/miR-15a-5p/HOXA3 axis was hypothesized to regulate Wnt/β-catenin signaling in PTC cells. Using Western blot, the levels of three core proteins in the Wnt/β-catenin signaling pathway (β-catenin, c-Myc, and cyclin D1) were examined. The results showed that knockdown of HOXA-AS2 decreased the levels of all three of these proteins (Fig. 9). However, their levels could be recovered by knockdown of miR-15a-5p or overexpression of HOXA3 (Fig. 9). These results suggest the HOXA-AS2/miR-15a-5p/HOXA3 axis modulates PTC progression possibly by regulating Wnt/β-catenin signaling proteins.

The HOXA-AS2/miR-15a-5p/HOXA3 axis synergistically regulates the Wnt/β-catenin signaling pathway. The protein levels of β-catenin, c-Myc, and cyclin D1 were examined in PTC cells co-transfected with sh-NC, sh-HOXA-AS2, miR-15a-5p inhibitors, and pcDNA-HOXA3. Error bars represent the mean ± standard deviation of at least three independent experiments.
Discussion
Thyroid cancer is a common type of human cancer. 35 Although surgical resection is typically an efficient treatment for thyroid tumors, the 5-year survival rate is still bleak, 36 and many patients die of this disease. 37 Therefore, it is critical to find more novel therapeutic targets for thyroid cancer. As the most common subtype, PTC was the focus of the present study. Recently, lncRNAs attract increasing attention of researchers, due to their contribution to initiation and development of PTC. With that, we aimed to probe into the novel lncRNA-mediated molecular mechanism in PTC. Currently, we focused on the functional and mechanistic implication of HOXA-AS2 in PTC.
LncRNAs are known to regulate various genes that are crucial for the initiation of tumors 38 and have oncogenic functions in various malignant tumors. 39 –43 In particular, lncRNA HOXA-AS2 has been shown to act as a tumor promoter in hepatocellular carcinoma, 44 pancreatic cancer, 18 and colorectal carcinoma. 21 However, it is unclear whether HOXA-AS2 is involved in PTC progression. In the present study, we first found the significant upregulation of HOXA-AS2 in PTC tissues and cell lines. Kaplan-Meier method revealed the negative relevance between HOXA-AS2 upregulation and overall survival of PTC patients, implying the potential involvement of HOXA-AS2 in PTC progression. Results from loss-of-function assays indicated that HOXA-AS2 positively regulated PTC cell proliferation, apoptosis, migration and invasion, acting as an oncogene. In mechanism, we observed that HOXA-AS2 could promote HOXA3 expression in PTC cells. Further functional assays demonstrated that HOXA3 exerted oncogenic role in PTC, consistently with HOXA-AS2. Many previous studies have reported that lncRNAs can upregulate mRNAs by acting as ceRNAs. 45 –47 Influenced by this notion, we speculated that HOXA-AS2 might act as a ceRNA in PTC to upregulate HOXA3. Using bioinformatics analysis and mechanistic experiments, it was observed that HOXA-AS2 functioned as a ceRNA via sponging miR-15a-5p to release HOXA3 expression in PTC. Besides, the positive impact of HOXA-AS2-miR-15a-5p-HOXA3 axis on PTC cellular process and tumor growth was validated by rescue assays and in vivo experiments.
The Wnt/β-catenin signaling pathway has been shown to be a critical element in the tumorigenesis of human tumors. 48 Specifically, Wang et al. reported that lncRNA PTCSC3/miR-574-5p modulates cell proliferation and migration in PTC via Wnt/β-catenin signaling. 49 Therefore, the possibility of an interaction between the Wnt/β-catenin signaling pathway and HOXA-AS2 in PTC was investigated in the present study. Here, the effect of the HOXA-AS2/miR-15a-5p/HOXA3 axis on the core Wnt/β-catenin signaling factors β-catenin, c-Myc, and cyclin D1 50,51 was measured. Importantly, HOXA-AS2, miR-15a-5p, and HOXA3 significantly regulated the levels of all three proteins, suggesting HOXA-AS2/miR-15a-5p/HOXA3 axis modulation of PTC progression via the Wnt/β-catenin signaling pathway.
In conclusion, the present study revealed that HOXA-AS2 functions as an oncogene in PTC and promotes progression of this disease by acting as a ceRNA, sponging miR-15a-5p, and regulating HOXA3. In turn, the HOXA-AS2/miR-15a-5p/HOXA3 axis modulates core proteins involved in Wnt/β-catenin signaling, supporting tumor growth. These findings will significantly contribute to further research and the development of more efficient treatments for thyroid cancer in the future.
Footnotes
Acknowledgments
The authors thank all subjects who participated in the study.
Author Disclosure
There are no conflicts of interest to disclose.