
Abstract
Corneal epithelial dystrophies are typically characterized by symptoms such as pain, light sensitivity, and corneal opacification leading to impaired vision. The development of gene therapy for such conditions has been hindered by an inability to achieve sustained and extensive gene transfer, as the epithelium is highly replicative and has evolved to exclude foreign material. We undertook a comprehensive study in mice aiming to overcome these impediments. Direct injection of lentiviral vector within the stem cell niche resulted in centripetal streaks of epithelial transgene expression sustained for >1 year, indicating limbal epithelial stem cell transduction in situ. The extent of transgene expression varied markedly but at maximum covered 26% of the corneal surface. After intrastromal injection, adeno-associated viral (AAV) vectors were found to penetrate Bowman's membrane and mediate widespread, but transient (12–16 days), epithelial transgene expression. This was sufficient, when applied within a Cre/lox system, to result in recombined epithelium covering up to approximately 80% of the corneal surface. Lastly, systemic delivery of AAV2/9 in neonatal mice resulted in extensive corneal transduction, despite the relative avascularity of the tissue. These findings provide the foundations of a gene therapy toolkit for the corneal epithelium, which might be applied to correction of inherited epithelial dystrophies.
Introduction
T
Gene therapy is conventionally applied to the correction of inherited genetic defects, and a number of such conditions have been described affecting the epithelium. Individually these are rare; however, many of them display an increased prevalence within the Japanese population. 12 –14 Gelatinous droplike corneal dystrophy (GDLD) is inherited in an autosomal recessive pattern, caused by a loss of function in the gene TACSTD2, 15,16 and is both among the most prevalent (1 in 33,000 in Japanese populations) and most severe of the epithelial dystrophies. Its primary phenotype is the accumulation of subepithelial amyloid deposits that render the cornea opaque, 17 –19 and until recently the only treatment commonly applied to manage GDLD was anterior lamellar keratoplasty (ALK). However, this procedure does not resolve the disease permanently as ALK only replaces the central portion of the cornea, allowing the recipient's limbal stem cells to remain in situ. Healthy graft epithelium is therefore replaced over time by the recipient's diseased epithelial cells via the process of normal corneal renewal. Recurrence of the condition thus often occurs within 1–2 years. 20 An alternative that has shown some initial promise for the treatment of inherited epithelial dystrophies is the supplementation of ALK surgery with allogeneic LESC transplantation. 21,22 LESC transplantation has become established as a successful treatment for limbal stem cell deficiency caused, for instance, by alkali burns. Although limbal epithelial transplants can be effective in this context, 23 their long-term efficacy and mechanism of action are unclear, especially in the case of allogeneic transplantation. It is often assumed or implied that reconstitution of the epithelium results from successful engraftment of the transplanted stem cells, but this may only be partially true. Studies that have genetically characterized restored corneal epithelial tissue subsequent to allogenic LESC grafts have found that in many cases the restored healthy tissue is derived from the recipient rather than the donor. 24 –26 This suggests the engrafted cells may largely function to support or restore the recipient's remaining endogenous limbal stem cells. Thus, this approach may not be optimal for the long-term treatment of inherited epithelial dystrophies. Moreover, many allogeneic LESC grafts eventually fail in the long term (5+ years), possibly due to an incomplete/failed engraftment or insufficient immunosuppression. 27,28
To date, the development of corneal epithelial-directed gene therapy has been limited. To be effective, corneal epithelial gene therapy must overcome the challenge of mediating stable gene delivery to a dynamic tissue that has evolved to exclude infectious agents and, to date, the most promising results have been achieved with lentiviral vectors. In the adult human corneal rim these have been shown to mediate efficient gene transfer to the corneal epithelium ex vivo, with epithelial transgene expression being shown at 3 days posttransduction. 29 The most sustained expression demonstrated thus far lasted 6 weeks. This was achieved in vivo by topical delivery of a lentiviral vector to the rat cornea after epithelial scarification 30 ; however, the efficiency of transduction appeared limited. The same study also provided the only example of application of adeno-associated virus (AAV) to target the corneal epithelium that we are aware of at present. Transgene expression resulting from topical application of an AAV2/2 vector was found to be more limited than that obtained with the lentiviral vector, both in terms of extent and duration, which was around 2 weeks. In terms of application, a promising small interfering RNA (siRNA)-based gene therapy has been developed for Meesmann epithelial dystrophy. The siRNA was shown to mediate specific and substantial (63%) knockdown of the mutant allele within patient epithelium ex vivo and was able to phenotypically correct a transfected epithelial cell line. 31 However, the authors note that the therapeutic siRNA lacks a suitable delivery vehicle for efficacious application in vivo.
There remains therefore significant scope for both improvement and application of gene transfer to the corneal epithelium, and here we present a comprehensive study examining multiple approaches aiming to mediate extensive and sustained gene delivery to this tissue via gene delivery to LESCs in situ.
Materials and Methods
Animals
The mouse strains used were wild-type C57BL/6J and Ai9 (007909; Jackson Laboratory, Bar Harbor, ME), which contain a loxP-flanked tdTomato reporter gene at the Gt(ROSA)26Sor locus. All mice were maintained under a 12-h light–dark cycle. Unless otherwise stated all animals used were adult (10+ weeks). Wild-type animals were all females, whereas Ai9 mice were mixed sex. All experiments were approved by the local institutional animal care and use committees (UCL, London, UK) and conformed to the guidelines on the care and use of animals adopted by the Society for Neuroscience (Washington, D.C.) and the Association for Research in Vision and Ophthalmology (Rockville, MD).
Recombinant AAV vector preparation and titer
Recombinant AAV (rAAV) vectors were produced via triple transient transfection. The plasmid construct, AAV serotype-specific packaging plasmid, and helper plasmid were mixed at a ratio of 1:1:3 at 20 μg of total DNA per milliliter of Dulbecco's modified Eagle's medium; mixed with polyethylenimine (Polysciences Inc., Warrington, PA) to a final concentration of 50 mg/mL; and incubated for 10 min at room temperature to form transfection complexes. These were added to HEK293T cells at 50 μg of DNA per 175-cm2 flask and left for 72 h. The cells were collected, concentrated, and lysed by freeze–thawing (4 × ) in TD buffer to release the vector. All AAV serotypes except AAV2/9 were purified by affinity with an AVB Sepharose column (GE Healthcare, Piscataway, NJ) and eluted with 50 mM glycine (pH 2.7) into 1 M Tris (pH 8.8). AAV2/9 was purified by a combination of size-exclusion and anion-exchange chromatography, using Sephacryl S300 and Poros HQ50, respectively (GE Healthcare). Vectors were washed in 1 × PBS-MK and concentrated to a volume of 100–150 μL, using Vivaspin 4 (10-kDa) concentrators (Vivaproducts, Littleton, MA). Vector genome (VG)-based titers were determined by quantitative real-time PCR (qPCR), using an inverted terminal repeat (ITR) binding assay as previously described. 32
Lentiviral vector preparation and titer
HIV-1-based lentiviral vectors were also produced by triple transient transfection. The plasmid construct, second-generation packaging plasmid, and envelope plasmid, mixed at a ratio of 4:1:3 at 32 μg/mL of Opti-MEM, were mixed with polyethylenimine (Polysciences Inc.) to a final concentration of 50 mg/mL and incubated for 10 min at room temperature to form transfection complexes. These were added to HEK293T cells at 80 μg of DNA per 175-cm2 flask and left for 72 h. Vector was purified and concentrated from supernatant medium by filtration (0.45 μm) and ultracentrifugation (90,000 relative centrifugal force) at 48 and 72 h. Viral particle (VP)-based titers were determined by infection of HEK293T cells with a serial dilution of concentrated vector followed by enumeration of infectious events.
Intraocular injections
Before all surgeries mice were anesthetized with a single intraperitoneal injection of a 0.01-mL/g mixture of Domitor (medetomidine hydrochloride, 1 mg/mL), ketamine (100 mg/mL), and water at a ratio of 5:3:42. After all surgeries anesthesia was reversed and a 1% chloramphenicol ointment (FDC International, Hampshire, UK) applied topically to the cornea.
Intrastromal injections were performed under observation thorough an operating microscope. Vector or control solution was injected with a 34-gauge hypodermic needle mounted on a 5-μL Hamilton syringe (Hamilton, Reno, NV). The needle tip was positioned (bevel up) about one-half to one-quarter of the corneal radius in from limbus and penetrated the cornea in this position. The needle was then pushed into the cornea, taking care to remain within the stroma. Once the needle tip was positioned centrally a 2-μL volume of vector or control solution was injected. On injection the cornea became opaque in the area of the injected bleb (usually extending over most of the cornea); this was completely resolved 24 h postinjection.
Limbal subepithelial injections were also performed under observation through an operating microscope. The eye was mildly prolapsed and secured in position with an elasticated Vinyl drape. Vector or control solutions were injected with a needle pulled from borosilicate glass capillaries (1.0 mm O.D. × 0.78 mm I.D.; Harvard Apparatus, Holliston, MA) and an injection holder set (IM-H1; Narishige, Tokyo, Japan). The pulled capillaries had a tip diameter <10 μm and a long taper morphology. Flow of solution to be injected was continuous, driven by approximately 1 MPa of air pressure. While solution was flowing multiple injections were performed (∼30–50) around the entire limbal region of the cornea. Because of the continuous flow neither the total volume injected nor the volume per injection could be accurately measured. Precise vector dose is therefore unknown.
Systemic delivery of AAV2/9 via intraperitoneal injection
These injections were all performed on postnatal day 0 (P0) mice within 12 h of birth. Injections were performed under observation thorough an operating microscope and using a 34-gauge hypodermic needle mounted on a 10-μL Hamilton syringe. The animal was physically restrained and the needle guided into the peritoneal cavity through the abdominal wall. The injection volume was 10 μL.
In vivo imaging
Before all in vivo imaging mice were anesthetized to minimize any motion blurring. Green fluorescent protein (GFP) fluorescence was imaged by scanning laser ophthalmoscopy. The instrument used was a Spectralis HRA2 with a 55° lens (Heidelberg Engineering, Heidelberg, Germany) modified for use with animals and with increased 488-nm excitation laser power. All images were taken at the 25° angle setting. Infrared (790-nm) images were taken at a power setting of 75%, and 30 images were averaged and normalized. GFP (488-nm) images were taken at a power setting of 95%, and 30 images were averaged and normalized. A single infrared image was taken to visualize the positioning of the eye. GFP expression was visualized by compositing multiple 488-nm channel images from multiple focal planes. Red fluorescent protein (RFP) fluorescence was imaged by fundoscopy. The instrument used was a slit lamp (SC-16; Keeler Instruments, Malvern, PA) modified for use with animals and with a filter set to detected RFP (excitation, 550 ± 15 nm; emission, 580 nm longpass). Images were taken with an attached DC500 digital camera and associated software (Leica, Wetzlar, Germany). Before imaging, corneal refractive power was neutralized by placing a 5-mm coverslip on the cornea covered with a coupling medium solution (Viscotears; Novartis Pharmaceutical, Basel, Switzerland). All images were taken at × 25 magnification and the following exposure times were used: bright field, 0.2 s; RFP, 30 s.
Histology
For corneal flat mounts eyes were enucleated postmortem and the cornea isolated by an orbital cut just posterior to the limbus. Tissue was fixed in 4% paraformaldehyde for 1 h before four evenly spaced radial cuts were made with a scalpel. Tissue was then counterstained with 4′,6-diamidino-2-phenylindole (DAPI) and mounted with DAKO fluorescence mounting medium (S3023; DAKO, Glostrup, Denmark). For corneal sections, eyes were enucleated postmortem, and the globe was punctured with the tip of a 30-gauge needle and then fixed in 4% paraformaldehyde for 1 h before embedding in optimal cutting temperature medium. Twenty-micrometer cryosections were cut in sagittal orientation, counterstained with DAPI, and mounted with DAKO fluorescence mounting medium. All images were acquired by confocal microscopy (DM5500Q; Leica). CD31 staining was achieved with rat anti-mouse CD31 antibody (MEC13.3; BD Biosciences, San Jose, CA) according to the protocol outlined by Cao and colleagues. 33
Quantification of transgene expression within the epithelium by confocal imaging of flat mounts
An area measuring 480 × 960 μm covering an area starting in the central cornea and extended toward a randomly determined quadrant was typically used for analysis. Images were taken at a z-resolution of 1 μm through the whole depth of the epithelium into the apical stroma. The peak of DAPI staining was found to correlate well with the dense basal epithelial cell layer, and an automated process could thus be used to align multiple samples to this reference point in the z-axis. In cases where this failed (usually due to poorly flattened tissue) the sample was aligned manually. The z-planes that constituted the area of interest were selected and a pixel intensity threshold set manually that satisfactorily separated fluorescent signal from background. Data to be compared all used the same threshold value, and all other aspects of the imaging were kept consistent. An automated process then counted all pixels above threshold intensity in the z-slices of interest.
Results
Intraepithelial injection of integrating lentiviral vector transduces LESCs and results in sustained transgene expression within their lineages
The optimal means by which to mediate pan-epithelial transgene delivery is to target LESCs, which are responsible for the renewal and maintenance of the tissue. We therefore tested the efficacy of injecting an integrating HIV-1-based lentiviral vector (LNT-SFFV-eGFP; titer, 1 × 10 7 VP/mL) directly into the murine LESC niche around the entire circumference of the corneoscleral junction. We used an ultrafine glass needle to make about 30 injections per eye.
In total, 94 eyes were injected and of these 88 were suitable to undergo complete analysis; the remainder were either culled for histology or died before completion of the experiment. For the first 10–12 weeks postinjection (WPI) eyes were assessed weekly or fortnightly by scanning laser ophthalmoscopy for the development of the streaklike patterns of transgene expression indicative of early transient amplifying cell (eTAC) or LESC transduction. After this period, follow-up became monthly and was continued until at least 52 WPI. If, after 12 WPI, an eye did not display any streaks for more than two consecutive time points, follow-up was discontinued. Figure 1A shows these data as a Kaplan-Meier survival curve. Such a long period of follow-up was selected to definitively differentiate between LESC and eTAC transduction. At 52 WPI, 32% of the eyes had sustained streaks for the whole period, strongly indicating LESC transduction. The extent of resulting epithelial transgene expression varied substantially. The best example is shown in Fig. 1B, in which approximately 25% of the corneal surface is covered by epithelium expressing GFP. A more typical example is shown in Fig. 1C, in which coverage was approximately 7%. Because of the lack of a specific pan-epithelial promotor, the ubiquitous spleen focus-forming virus (SFFV) promotor was used. Transgene expression might not therefore be expected to be restricted to the epithelium, and indeed stromal expression was also seen. Although stromal and epithelial expression could be reliably distinguished on the basis of spatial and temporal patterning, confocal microscopy of corneal flat mounts postmortem confirmed the premise that the streaks were indeed epithelial (Fig. 1D).

Intraepithelial injection of lentiviral vector mediates sustained epithelial transgene expression via limbal epithelial stem cell (LESC) transduction.
Intrastromal injection of AAV (but not lentiviral) vector mediates transient epithelial transduction across Bowman's membrane
Although the transgene expression achieved by injecting a lentiviral vector directly into the limbus is superior to any other transduction achieved to date, the injections proved technically demanding. Thus, the success rate was relatively low (∼32% at 52 WPI) and the best transduction achieved remained limited in terms of spatial extent (∼25% of the corneal surface). A potentially more reproducible approach is to target the epithelium from the basal direction through Bowman's membrane via an intrastromal injection. Lentiviral and AAV vectors were both assessed for this ability to transduce the corneal epithelium via this route of administration. Intrastromal injections of either LNT-SFFV-eGFP (1 × 10 6 VP/eye) or AAV2/8[Y733F]-CMV-eGFP (1 × 10 11 VG/eye) were carried out in adult mice (n = 4 eyes per vector). Animals were culled at 4 days postinjection (DPI) and transgene expression assessed on cryosections. In all cases lentiviral vector-mediated transgene expression was restricted to the stroma (Fig. 2A), but AAV vector-mediated expression was seen in both the stroma and the epithelium (Fig. 2B). In contrast to lentiviral vectors, rAAV vectors do not efficiently integrate their genomes into that of the host. 34 In the highly replicative epithelium, transgene expression would therefore be expected to decline over time due to loss of transduced cells from the corneal surface and to the dilution of vector episomes on cell replication. We did indeed observe such an effect; we noted that at 8 DPI AAV-mediated epithelial transgene expression appeared maximal (Fig. 2C), and by 18 DPI (Fig. 2D) had been almost entirely lost.

Intrastromal injection of adeno-associated virus (AAV) mediates transient trans-Bowman's epithelial transduction.
Having shown that AAV is able to penetrate Bowman's membrane and mediate transient transduction of the epithelium, we examined which AAV serotype was best suited to this purpose. Six AAV serotypes were assessed: AAV2/5, AAV2/8, AAV2/9, AAV2/8[Y733F], AAV[ShH10], and AAV[Anc80] titer matched to a dose of 1.4 × 10 11 VG/eye (n = 6 eyes per serotype). Animals were culled at 8 DPI and corneas were flat mounted. A region of each cornea was selected for analysis by confocal microscopy; a representative orthogonal projection of such a region is shown in Fig. 2E. Transgene expression (eGFP signal above threshold) in the epithelium of this region was then quantified (Fig. 2F). All the AAV serotypes tested mediated corneal epithelial expression. However, AAV2/8[Y733F] and AAV[ShH10] mediated substantially more transgene expression than the other serotypes tested.
Intrastromal injection of AAV-Cre mediates genomic recombination in LESCs that is propagated to their lineages
Any transduction of LESCs achieved via intrastromal injection of AAV would not be visualized with an eGFP transgene because of the lack of vector integration and the temporal limitations outlined above. The Cre/lox recombination system, however, allows permanent expression of a marker gene to result from transient expression of Cre recombinase. This system was therefore used to assess the ability of intrastromal AAV to transduce LESCs. Intrastromal injections (1 × 10 11 VG/eye) of AAV2/8[Y733F]-CMV-Cre were performed in Ai9 mice whose cells express the fluorescent tdTomato transgene on exposure to Cre. Sagittal sections of injected corneas at 21 DPI show extensive RFP expression both in the central cornea (Fig. 3A) and at the basal corneoscleral junction where the LESCs are located (Fig. 3B). A small cohort (n = 4) was assessed by fundoscopy fortnightly for 17 weeks, a period that was sufficient to be highly indicative of LESC transduction based on our lentiviral data. All four eyes developed centripetal streaks of recombined cells and sustained them over the entire period of follow-up. Even within this small cohort it was clear that both the extent and reliability of LESC transduction improved on that achieved via intraepithelial injection of a lentiviral vector. Figure 3C shows the greatest spatial extent of recombined epithelium observed (∼82% of corneal surface) and Fig. 3D the least (∼22%). The latter figure is equivalent to maximum extent achieved with a lentiviral vector. Across the whole group, the mean (±1 SD) extent was 53 ± 25.0%. It was also noted that before 10 WPI, epithelial streaks became increasingly more defined over time. This perhaps represents the extinction of lineages derived from late, central TACs, leaving solely those derived from LESCs and early, more limbal TACs. Again, because a ubiquitous promoter was used, both stromal and epithelial recombination was expected and observed, and thus histology was used to confirm localization of tdTomato within the epithelium (Fig. 3E).
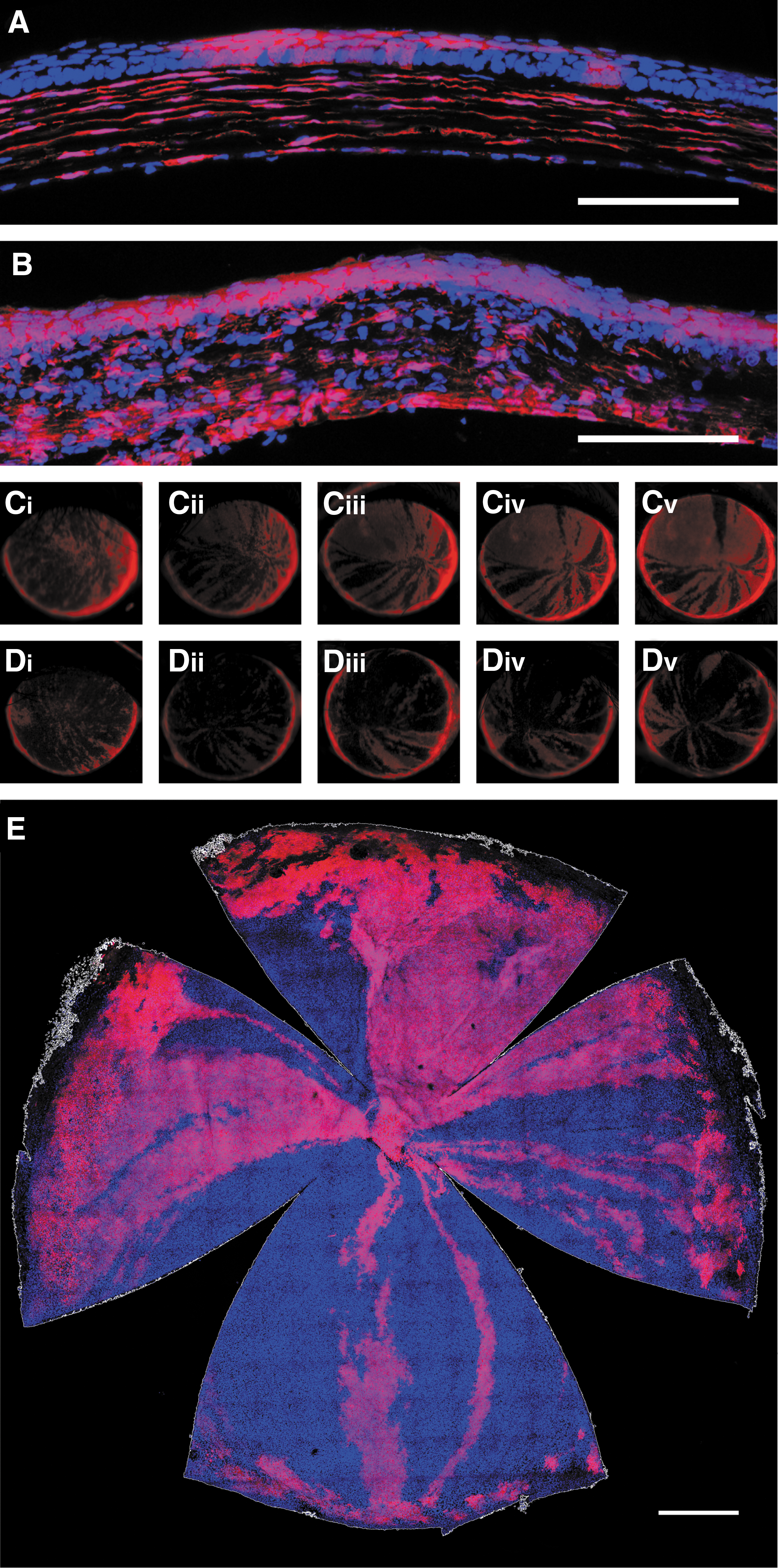
Intrastromal injection of AAV can mediate genomic modification of LESCs within Ai9 mice.
Intraperitoneal injection of AAV2/9 on P0 mediates gene delivery to all layers of the cornea, including the epithelium
AAV2/9 has been shown to result in widespread somatic gene delivery if delivered systemically at early postnatal time points. 35 We therefore assessed its ability to transduce the cornea after systemic injection in neonatal mice. Intraperitoneal injections (7.2 × 10 11 VG per animal) of AAV2/9-CMV-Cre were performed in P0 Ai9 mice.
At 28 DPI, extensive tdTomato expression was seen in flat-mounted corneas (Fig. 4A). To better visualize the distribution of expression with regard to corneal layers, a pixel intensity threshold (127–255) was applied to each z-plane of the confocal image, and each plane was then false colored by depth (Fig. 4B) with the ImageJ 36 ICA look-up table. Layer-specific z-stacks were also taken at a higher magnification. The majority of tdTomato expression is localized within the stroma (Fig. 4B and D), but multiple islands of epithelial expression are also seen that appear to result from clonal expansion (Fig. 4B and E) and some expression was also seen in the endothelium (Fig. 4C). Over time these islands of epithelial expression developed into centripetal streaks (Fig. 4F–H), which persisted until at least 21 weeks postinjection and many of which originated in the limbus.
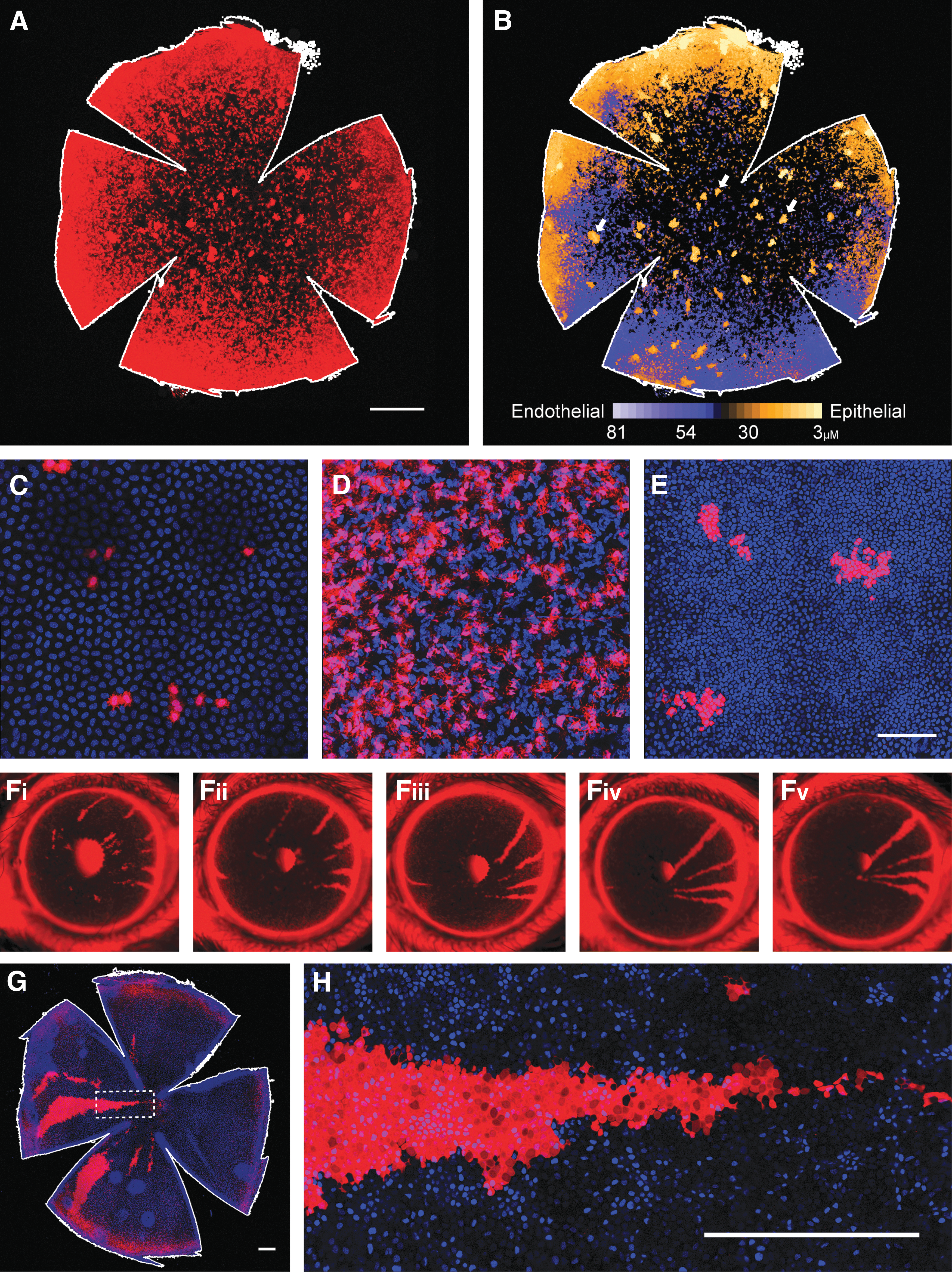
Intraperitoneal injection of AAV2/9 on postnatal day 0 (P0) mediates gene delivery to all layers of the cornea including the epithelium.
Discussion
We have presented a comprehensive study of gene transfer to corneal epithelial cells, utilizing several different routes of vector administration and both lentiviral and AAV-based vector systems. We achieved substantial improvements over the results achieved to date both in terms of extent and duration of transgene expression. Intraepithelial injection of lentiviral vector at the LESC niche is the most intuitive and direct of these approaches and thus is likely the most suitable for further development and clinical application. This approach gave rise to epithelial transgene expression that was sustained for more than 1 year and thus likely resulted from LESC transduction. Technically, however, these injections were demanding and the extent of transduction achieved, although superior to any documented to date, remained somewhat limited. In the human eye, which is larger and possesses visible landmarks 37 to indicate the LESC niche, it is likely that the procedure may prove less challenging. However, the human LESC niche is also substantially more complex than the murine, being associated with specialized structures within the limbus, which might complicate the procedure. 38,39 Therefore, optimization within a larger animal model, such as the pig, with a limbal anatomy more closely resembling that of the human may be required to develop the procedure further.
Gene therapy in which an integrating vector is used to correct a genetic disorder via transduction of an adult stem cell resulting in therapeutic gene expression throughout the entire resulting cell lineage is a well-established approach. Indeed, some of the first successful clinical applications of gene therapy used this methodology, transducing hematopoietic stem cells ex vivo for the treatment of severe combined immune deficiency. 40,41 More recently, the technique has been applied to epithelial stem cells of the skin in the correction of junctional epidermal bullosa. 42 In such cases, however, the stem cells are extracted from the patient and transduced ex vivo, followed by reengraftment. This approach presents the potential advantages of allowing selection and amplification of transduced clones before engraftment 42 and avoiding direct exposure of the patient to the gene therapy vector. However, the effectiveness of this strategy is entirely dependent on the feasibility of achieving functional engraftment. In the case of LESCs the clinical data to date cast some doubt on the efficiency of donor cell engraftment, 24 –26 and thus gene transfer to the adult stem cell in situ may constitute a superior approach for the correction of epithelial dystrophies.
Our second approach was to apply the technically simpler technique of intrastromal injection to transduce the epithelium across the physiological barrier of Bowman's membrane. Whereas lentiviral vectors were found to be unable to penetrate this barrier, AAV vectors mediated substantial transduction, possibly because their much smaller size allowed dissemination across the membrane. To date there has been little evidence of AAV having any substantial efficacy in the corneal epithelium. 30 Nonetheless, we found that not only was AAV able to mediate substantial epithelial transduction, but that all AAV serotypes tested were able to do so, albeit with some variation in efficacy. This would suggest that this tropism is relatively robust and not dependent on any single mechanism of viral entry, adding to the likelihood that some serotypes of AAV will maintain this tropism across species. The murine Bowman's membrane is known to be relatively thin in comparison with that of humans, 43 –45 and thus it is possible that AAV dissemination across this more substantial barrier would be more limited than that observed here. However, developing serotypes able to overcome the physical barriers present within a tissue has been a major focus of AAV vector development and engineering. Specifically, within the eye, achieving substantial transduction of photoreceptors via intravitreal injection of vector has been a longstanding objective. 46,47 This requires the AAV virion to penetrate the inner limiting membrane and ∼200 μm of neural tissue, which several engineered serotypes have now been shown capable of doing. Experiments in the anterior segment revealed that AAVAnc80 can penetrate Descemet's membrane and mediate transduction of the corneal stroma after an intracameral injection. 48 These data suggest that some AAV serotypes may be able to mediate epithelial transduction even across the more substantial barrier posed by a human Bowman's membrane, although this still needs to be established. The intrastromal injection technique applied here (complete stromal hydration) would likely be undesirable within the clinical context. A more applicable technique would be to make multiple small-volume intrastromal injections around the limbal circumference in order to maximize vector delivery to LESCs and minimize the risk of stromal scarring within the visual axis.
The major limitation of rAAV within a replicative tissue is its inability to replicate its genome on host cell division, and indeed in the corneal epithelium we observed this resulting in loss of transgene expression over time. We observed that near complete loss of epithelial transgene expression took around 18 days, in agreement with previous estimates of approximately 3 weeks for complete epithelial turnover. 9 Loss of transgene expression therefore proceeded too rapidly to determine the ability of AAV vectors to transduce LESCs. We therefore combined intrastromal injection of AAV with a Cre recombinase system and showed that not only was AAV-mediated recombination sustained for long enough to be highly indicative of LESC transduction, but that the extent of recombined lineages also began to approach pan-epithelial coverage. The Cre recombinase system is a laboratory tool unsuitable for clinical application; however, the key attribute of the system utilized here is the induction of a permanent genomic change via transient transgene expression. This property is shared by the much more clinically applicable CRISPR gene-editing system and, moreover, sustained expression of Cas9 may not be desirable because of an increased risk of off-target cleavage. 49 These results suggest that combining CRISPR technology with the ability of AAV to mediate extensive but transient LESC transduction in situ, as demonstrated here, might be a promising future avenue for the correction of epithelial dystrophies.
Intrastromal and limbal subepithelial injections attempted to deliver vector either directly within the target LESC niche or close to it. Although such approaches provide an obvious benefit in terms of cellular targeting and transduction efficiency, they are also invasive and technically demanding procedures. It has previously been demonstrated that AAV2/9 has the ability to mediate widespread transgene delivery seemingly throughout the entire murine soma (including some evidence of corneal transduction) when delivered systemically at an early postnatal time point.
50
This facet of AAV2/9 has been exploited in the treatment of young infants with spinal muscular atrophy 1 with some promising results.
51
We therefore examined the potential corneal application of this route of administration in more detail, specifically with regard to the corneal epithelium and LESCs. Systemic propagation of vector likely occurs via dissemination in blood, and within the retina it has been shown that the extent of transduction resulting from systemic AAV2/9 delivery is closely correlated with the extent of retinal angiogenesis at the point of injection,
52
a finding mirrored in our study (Supplementary Fig. S1B; supplementary data are available online at
To conclude, here we have presented an extensive comparison of vectors and administration routes in order to begin to develop a gene delivery toolkit for the corneal epithelium, with potential applications in the correction of corneal epithelial dystrophies by gene therapy.
Footnotes
Acknowledgments
Ms. Ryea Maswood, Ms. Olha Semenyuk, and Dr. Scott Robbie provided technical support. Dr. Ulrich Luhmann provided advice and guidance. This study was supported by grants from Fight for Sight (grant 1783), the BRC for Ophthalmology at Moorfields Eye Hospital and UCL Institute of Ophthalmology (IS-BRC-1215-20002), the European Research Council (grant ERC-2012-ADG_20120314), and the Medical Research Council (grant MR/M007871/1).
Author Disclosure
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.