
Abstract
Disparities at the major histocompatibility complex (MHC) antigens and associated minor antigens trigger harmful immune responses, leading to graft rejection after transplantation. We showed that MHC-silenced cells and tissues are efficiently protected against rejection. In complex vascularized organs, the endothelium is the major interface between donor and recipient. This study therefore aimed to reduce the immunogenicity of the lung by silencing MHC expression on the endothelium. In porcine lungs, short-hairpin RNAs targeting beta-2-microglobulin and class II-transactivator transcripts were delivered by lentiviral vectors during normothermic ex vivo perfusion to silence swine leukocyte antigen (SLA) I and II expression permanently. The results demonstrated the feasibility of genetically engineering all lung regions, achieving a targeted silencing effect for SLA I and II of 67% and 52%, respectively, without affecting cell viability or tissue integrity. This decrease in immunogenicity carries the potential to generate immunologically invisible organs to counteract the burden of rejection and immunosuppression.
Introduction
D
Regarding the possibility for genetic modification and clinical perspectives, the lung is exceptional among transplanted organs. In contrast to end-stage kidney disease where individuals may live with the support of hemodialysis, no bridging or replacement therapy exists for end-stage lung disease. In addition, despite the widespread success of lung transplantation as an established lifesaving therapy, graft survival rates of about 50% 5 years after transplantation demands the improvement of this approach. 13 Usually, the size, the complexity of the lung associated with several cell types, the low ischemic tolerance, and the unavailability of an universal gene delivery vector system compromise the efficacy of genetic engineering approaches. Nevertheless, the lung has already become a main target for gene therapy toward the correction of different diseases such as cystic fibrosis and α1-antitrypsin deficiency. 14 –16 Gene therapy of the lung is supported by the possibility of administering the vector through different routes. So far, these strategies have mainly focused on the in vivo modification of the lung epithelium, and therefore the conducting airways were used as the preferential administration route for therapeutic vectors rather than the pulmonary or bronchial arterial system, which could result in the undesired off-target modification of other tissues or organs. 17 –19
After solid organ transplantation, the graft's endothelium serves as the first line of contact between donor and recipient. The endothelium of a transplanted organ is the key cell layer responsible for triggering the direct pathway of allorecognition, may also be involved in triggering the indirect pathway, and is an important target during rejection. 20 Transplantation offers the chance to take advantage of the organs' inevitable ex vivo period and to utilize it for modifying their immunogenicity by genetic engineering. Until recently, this approach has been hindered by the equally unavoidable necessity for efficient cooling of the organs, which prevents efficient cell transduction with viral vectors. Ex vivo organ perfusion systems at normothermic conditions were shown to be powerful tools for evaluation and reconditioning of extended criteria donor organs. 21,22 This study extended the use of this technology as a strategy to genetically modify the endothelium of the lung efficiently without compromising its integrity. This study shows the efficient genetic engineering of a large and complex organ, the lung, during normothermic ex vivo lung perfusion (EVLP). In particular, stable silencing of MHC expression in the lung endothelium may significantly contribute to decrease the immunogenicity of the lung after allogeneic transplantation and combat the burden of rejection and immunosuppression.
Methods
Lentiviral vector constructs
A RNAi cassette was constituted by an H1 promoter and the short-hairpin RNA (shRNA) sequences targeting the transcripts for downregulating swine leukocyte antigen (SLA) expression. Either shRNA sequences targeting the swine beta2-microglobulin (β2m; shβ2m: 5′-CACGTGACTCTCGATAAGCC-3′) to silence SLA class I or the class II transactivator (CIITA; shCIITA: 5′-CTGCCACAGTACGACTTTGT-3′) to silence SLA class II were cloned into the
Lentiviral vector production
Large-scale production of lentiviral vector particles was performed in HYPERFlasks cell culture vessels (Corning, Corning, NY). Briefly, HEK293T cells were seeded into HYPERFlasks and cultured with Dulbecco's modified Eagle's medium 10% fetal calf serum until confluence was reached. Then, cells were co-transfected overnight with the shRNA-encoding vector, pRSV_Rev, pCDNA3.g/p.4xCTE, and pMD2.G. The next day, cells were washed, and fresh medium was added. After 48 h, supernatants were collected and centrifuged at 30,000 g for 3 h. Viral vector containing supernatants was titered using p24 enzyme-linked immunosorbent assay (Cell Biolabs, San Diego, CA).
Lentiviral vector–mediated lung transduction during normothermic ex vivo perfusion
Lungs of five 18–33 kg pigs were procured en bloc from the animals after low-potassium dextrane (Perfadex®; XVivo Perfusion AB, Göteborg, Sweden) solution cold-flush perfusion. Then, the lung blocks were mounted in a modified Organ Care System (OCS; TransMedics, Inc., Andover, MA) organ chamber by connecting the trachea and the pulmonary artery trunk to the respective OCS cannulas. In the modified OCS, the lungs were ventilated with a positive end expiratory pressure of 5, respiratory frequency of 8/min, and a tidal volume of 4 L/min. Perfusion of the lung was performed using a cardiac output of 0.7 L/min. Further, lungs were perfused with 1.5 L OCS solution (TransMedics, Inc.) by slowly warming up to 37°C within 15 min using a roller pump (Stöckert CAPS, Munich, Germany) set at 1.5 L/min flow. Then, 12 mg protamine sulfate (Sigma–Aldrich, St. Louis, MO) were infused into the circulating perfusion solution. Afterwards, 1 × 1011 shβ2m and 1 × 1011 shCIITA encoding vector particles were injected into the system and allowed to circulate for 2 h in three lung blocks. As control, the same number of viral particles encoding for the nonspecific shRNA (shNS) were used to perfuse the lungs of three pigs. Also, lungs of two pigs were perfused using the same conditions but without vector to serve as a further control. Then, the perfusion solution was substituted with porcine whole blood and 500 mL Steen solution (XVivo Perfusion AB). During the perfusion time, perfusate samples were collected for further analyses at the beginning and after every 30 min.
Preparation of endothelial cell cultures
After ex vivo perfusion, 1.5 cm
2
tissue samples were collected from 10 representative regions of the left lung (L1–L10) and right lung (R1–R10), as represented in Supplementary Fig. S1 (Supplementary Data are available online at
Real-time polymerase chain reaction
mECs were cultured for 1 week after isolation in EBM-2 medium. Afterwards, mECs were stimulated with tumor necrosis factor alpha (TNF-α) and interferon gamma (IFN-γ; 50 ng/mL; R&D Systems, Minneapolis, MN) for 48 h. Total RNA was isolated (RNease Mini Kit; Qiagen, Hilden, Germany) and reverse transcribed to cDNA using a High-Capacity cDNA Reverse Transcription kit (Applied Biosystems, Foster City, CA). Quantitative polymerase chain reaction was used to analyze β2m (ss03391154_m1) and SLA-DRA (ss03389945_m1; both from Thermo Fisher Scientific) transcript levels. As endogenous control, for normalization of mRNA levels, the HPRT gene (ss033388273_m1) was used. All samples were measured in duplicate.
Quantification of bioluminescence activity
Supernatants of endothelial cells cultured for 4 days were collected every 24 h for determination of bioluminescence activity. One μg/mL of the NL substrate furimazine (Promega) was added to the collected supernatants, and bioluminescence was measured with a luminometer (Berthold Technologies GmbH, Zug, Switzerland). Levels of bioluminescence were measured in independent lung regions (L1–L10 and R1–R10) or in the supernatants of cells of those regions pooled to represent the different lung lobes (LSL, LIL, RSL, RML, and RIL).
Analysis of the cytokine secretion profile during perfusion
Cytokines secreted into the perfusion solution during ex vivo lung perfusions were characterized using Luminex technology (Luminex Corp., Austin, TX). In particular, levels of porcine interleukin (IL)-1β, IL-2, IL-6, IL-8; IL-10, IL-12, TNF-α, and IFN-γ were quantified in the perfusate samples collected at the beginning and after every 30 min during organ perfusion.
Lung histology
After EVLP, random lung sections were collected, fixed in 4% paraformaldehyde solution, and embedded in paraffin. Sections were stained with hematoxylin and eosin and assessed by a specialized lung pathologist in a blinded manner.
Statistical analyses
Statistical analyses were performed using GraphPad Prism v5.0 (GraphPad Software, Inc., San Diego, CA). Data are presented as the mean ± standard deviation. For comparison of means between two groups, a two-tailed Student's t-test was used. p-Values <0.05 were considered significant.
Results
Detection of secreted luciferase activity in mEC cultures of lungs perfused with the lentiviral vector
The large size and 3D complexity of solid vascularized organs such as the lung remain challenging for the efficient perfusion and genetic engineering of the lung endothelium in all regions. Lentiviral vectors used for the genetic modification of the lung endothelium during normothermic ex vivo perfusion encoded for a secreted form of luciferase (NanoLuc, NL) as reporter gene (Fig. 1). Therefore, after perfusion, mEC cultures of 10 different regions of the right and left lungs (Supplementary Fig. S1) were established for detection of the reporter gene activity in the supernatants. The expression of the reporter gene continuously increased with time (Fig. 2). In comparison to mEC cultures derived from non-perfused lungs, significant increases (p < 0.001) in relative luminescence units were detectable in the supernatants of mECs perfused with NL-encoding vectors. Furthermore, mECs of both the right and left lungs as well of the different lobes expressed high levels of luciferase. Also, an increase in ML activity was detected in cell culture supernatants of mECs of lungs perfused with a control shNS-encoding vector. Analysis of the bioluminescence level in independent tissue samples indicates differences in the capacity of the vector to reach particular regions. However, no pattern for regions preferably transduced was observed for the left lung. In fact, the superior and inferior lobes of the left lung showed similar bioluminescence levels. In contrast, the middle lobe of the right lung showed higher bioluminescence levels in comparison to the superior and inferior right lobes, which may indicate preferential transduction of this region. These data indicate that this perfusion system enables the vector to reach and transduce all regions of the lung, but the accessibility of the vector to different regions may differ (Fig. 2C).
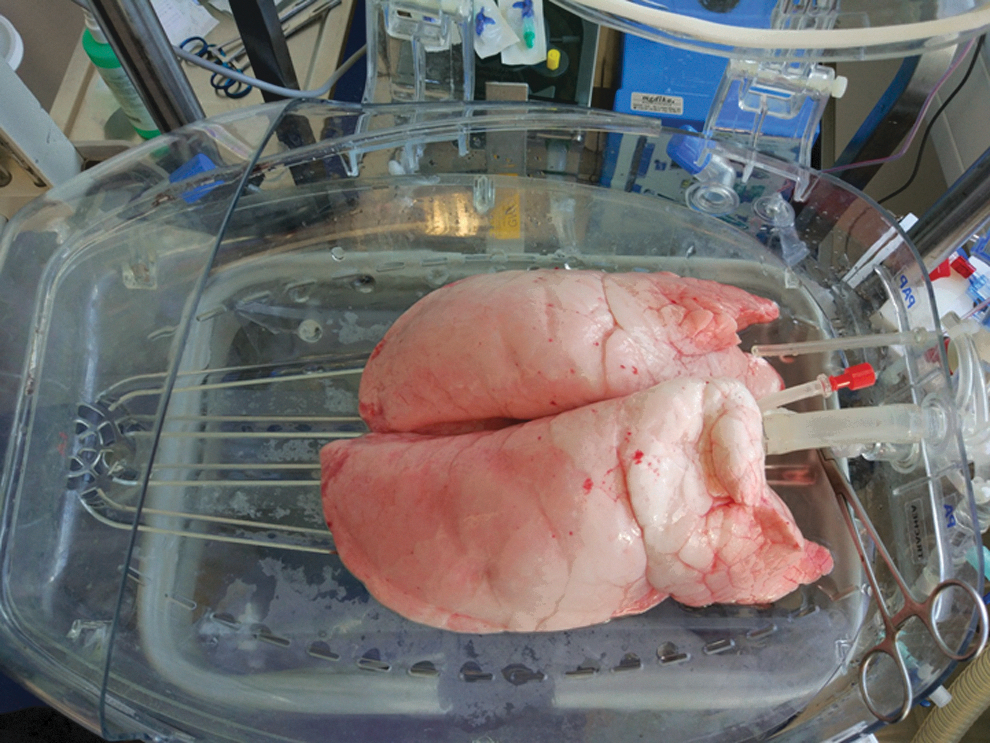
Genetic engineering of porcine lungs using a normothermic ex vivo perfusion system. The photo depicts a lung block connected to the perfusion system.

Genetic engineering of the lung endothelium. Lung blocks were perfused with a lentiviral vector encoding for swine leukocyte antigen (SLA)-specific short-hairpin RNAs (shRNAs; shβ2m/shCIITA) and nanoluciferase as a reporter gene. After perfusion, 10 representative tissue samples of different pulmonary regions of the left
Silencing SLA class I and class II transcripts in the lung endothelium
The use of optimized perfusion systems for the delivery of therapeutic vectors may become an efficient tool for regulation of targeted gene expression. Lung perfusion with shβ2m-encoding vectors resulted in downregulation by up to 60% in β2m transcript levels in mECs (Fig. 3A). Interestingly, the maximal silencing effect of the shRNA was observed upon mEC stimulation with IFN-γ and TNF-α. Lungs perfused without vector were able to upregulate β2m transcript levels by up to 13.7-fold, whereas a maximal increase of β2m of 5.1-fold was measured in the left superior lobe of lungs perfused with shβ2m-encoding vectors. mECs of the right inferior lobe could even only upregulate β2m levels by means of 3.6 ± 1.9-fold (Fig. 3B). The delivery of shCIITA into lung mECs caused a decrease by up to 96% in constitutive SLA-DRα transcript levels. Even under IFN-γ and TNF-α stimulation, shCIITA was able to prevent a strong upregulation of HLA class II (Fig. 3C). Non-transduced mECs cultured in the presence of IFN-γ and TNF-α were capable of upregulating HLA-DRα by up to 28.5-fold. In contrast, mECs of lungs exposed to shCIITA showed a significantly lower (p < 0.05) capacity to induce the transcription of SLA-DRα. A maximal upregulation of 12.7- and 12.9-fold upregulation of SLA-DRα was detected in the left superior lobe and right inferior lobe, respectively (Fig. 3D). SLA class I and class II transcript levels were similar to the control when shNS was used. These data show the feasibility of the vector-delivered shRNA to regulate gene expression in all lung regions.
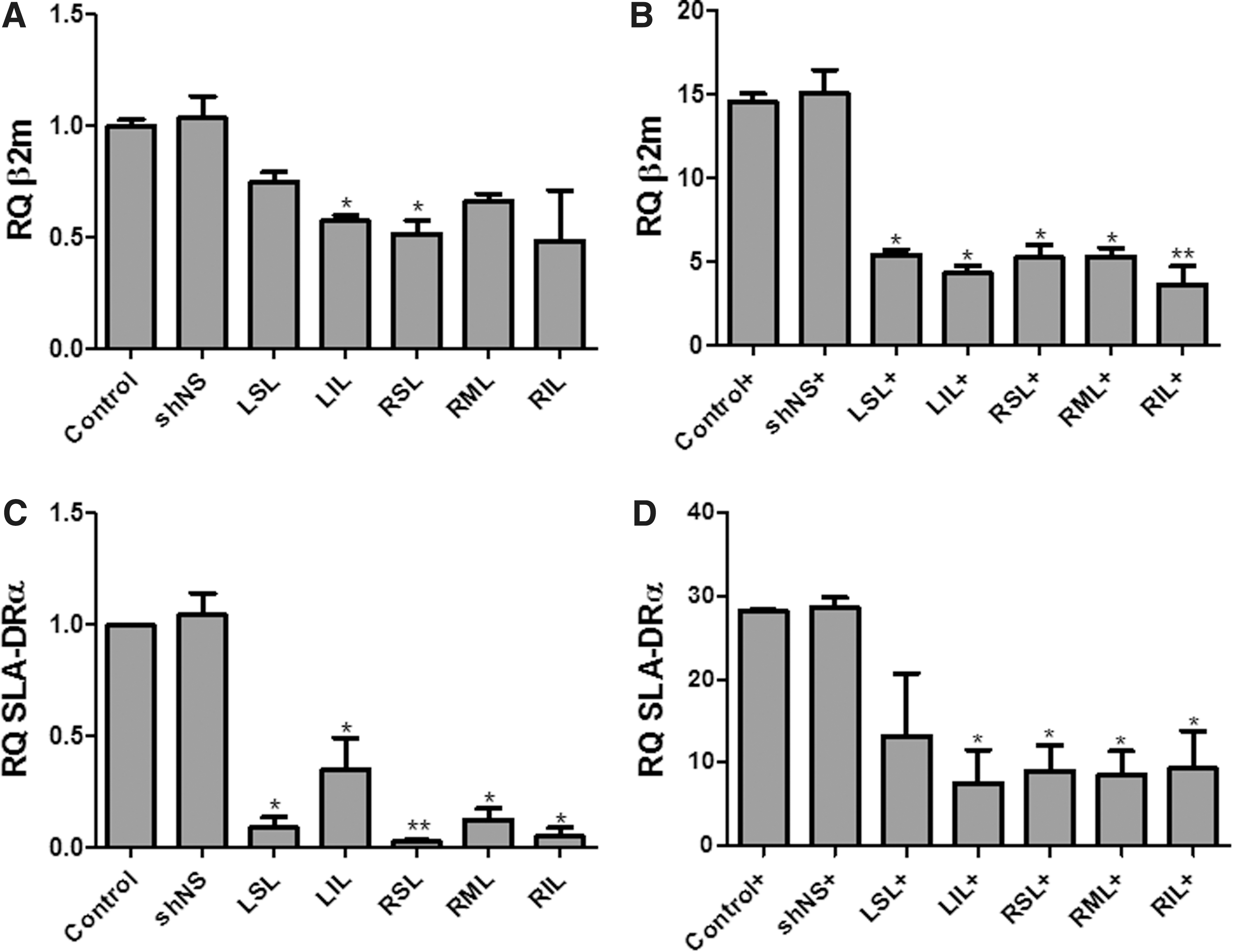
Silencing SLA class I and class II transcript levels on the lung endothelium. SLA class I and class II transcript levels were measured by real-time polymerase chain reaction.
Silencing SLA class I and class II protein expression in the lung endothelium
Antibody and T-cell receptor recognition of MHC molecules in the graft endothelium represent major triggers for allogeneic immune responses. Therefore, the effect of the shRNA delivery was evaluated at SLA protein level on mECs. shβ2m caused a 60.4 ± 19.2% (p < 0.001) decrease in SLA class I expression. After stimulation of mECs with IFN-γ and TNF-α, the levels of SLA class I proteins remained downregulated by 50.0 ± 17.03% (p < 0.001; Fig. 4). Similarly, SLA-DR expression was silenced by 45.6 ± 6.8% (p < 0.001) in unstimulated mECs perfused with shCIITA in comparison to the levels of expression derived from mECs of lungs perfused in the absence of the vector. Also, IFN-γ- and TNF-α-stimulated mECs showed a decrease in SLA-DR expression by up to 45% (Fig. 5). In comparison to non-transduced lungs, perfusion with shNS did not significantly alter SLA class I and class II expression (Figs. 4 and 5). These results show the feasibility of regulating protein expression levels after shRNA delivery in the lungs' endothelium.

Silencing SLA class I expression on the lung endothelium.
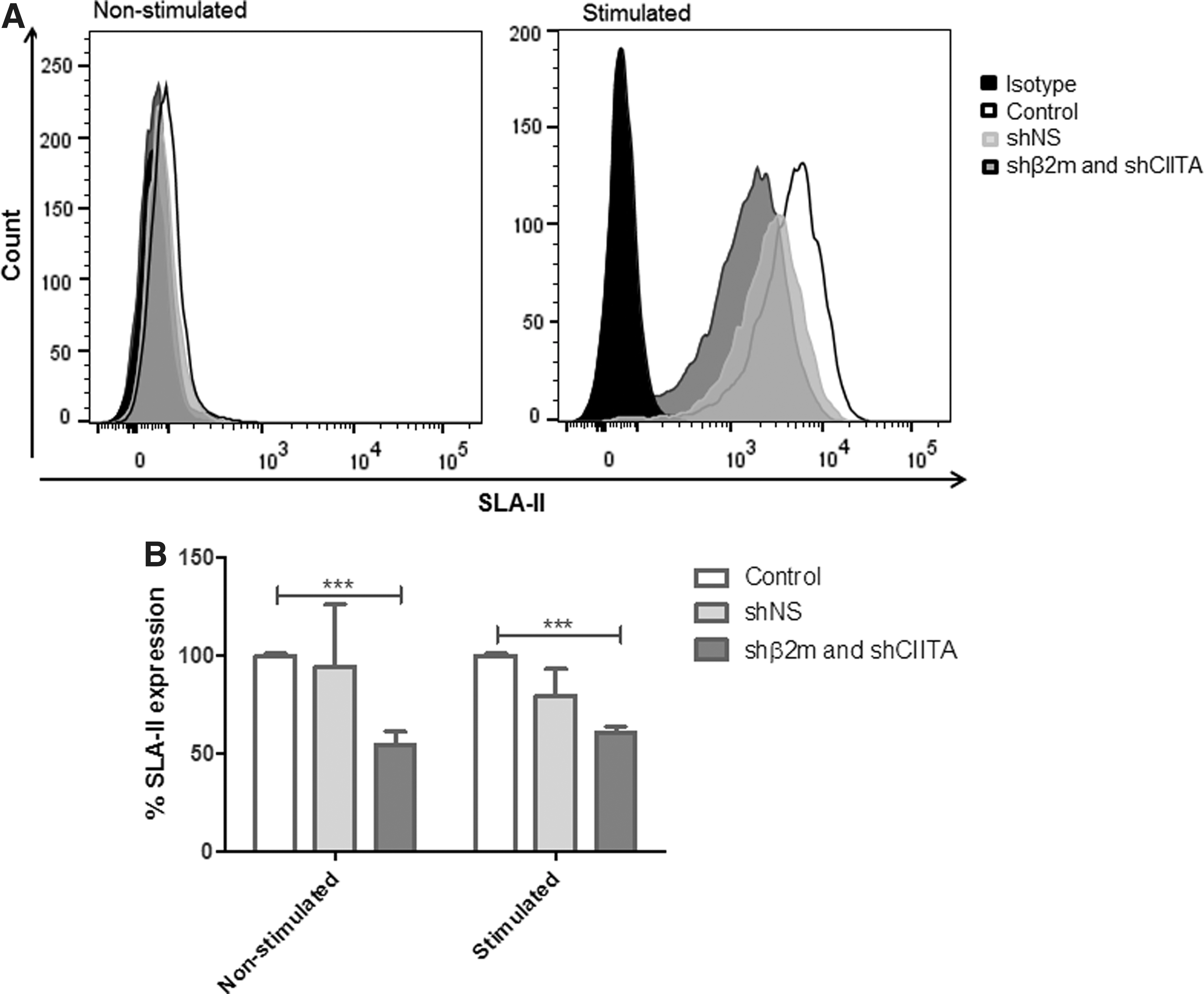
Silencing SLA class II expression on the lung endothelium.
Assessing the effect on lung quality of perfusion with lentiviral vectors
To evaluate any possible damaging effects on lung integrity of perfusion with the lentiviral vector, different assays were performed. Lactate dehydrogenase (LDH) levels were used as a marker for necrosis during perfusion. After 2 h of perfusion, levels of LDH were similar on the perfusates of lungs exposed or not exposed to the vector, indicating that perfusion with the vector does not induce cell injury (Fig. 6A). In addition, histological analyses after perfusion with the vector encoding for SLA-specific shRNAs showed similar tissue structure to shNS-expressing or non-transduced lungs (Fig. 6B). Perfusion of the lungs with the lentiviral vector increased levels of IL-6 by up to 0.106 pg/mL, IL-8 by up to 0.431 pg/mL, and TNF-α by up to 2.487 pg/mL (Fig. 6C). Secretion of IL-1β, IL-2, IL-10, IL-12, and IFN-γ was not detectable (data not shown). These results show that perfusion with lentiviral vectors does not induce tissue injury and induces secretion of low levels of pro-inflammatory cytokines.
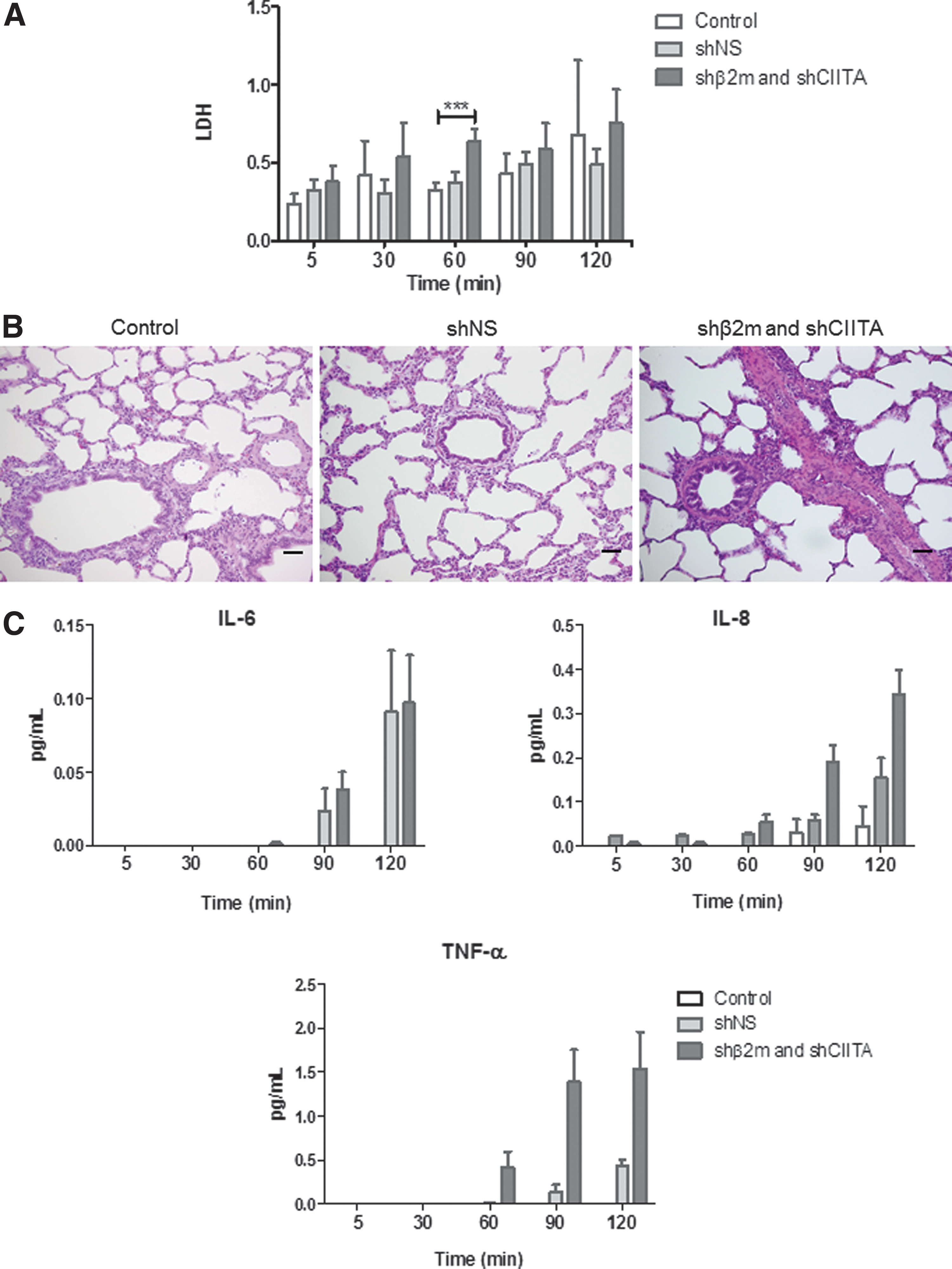
Assessment of quality parameters of the lung during perfusion.
Discussion
Modern transplantation medicine has made significant progress within the last decades due to a better immunological understanding of rejection and advances in immunosuppression. However, adverse effects of immunosuppressive therapy and a shortage of donor organs remain major limitations in transplantation. The severe side effects of lifelong immunosuppression to prevent graft rejection, including infections, malignancy, and kidney dysfunction, are a major burden for transplant recipients. 24 This study approaches this problem from a different perspective. Instead of inducing an immunological blindness of the recipient toward the allograft by immunosuppression or by expressing immunomodulatory molecules such as IL-10 or indoleamine 2,3-dioxygenase, 25,26 this study aimed to achieve immunological invisibility of the donor's organ. This may be performed by genetic modification of the graft during the inevitable ex vivo period of the allograft after explantation and until transplantation.
An allograft's immunogenicity is defined by the gene products of the MHC and the minor histocompatibility antigens (mHag) presented as peptides in MHC antigens. The mechanisms by which the immune system recognizes allogeneic cells centers around peptide-presenting MHC proteins on the cell surface, which are targeted by cytotoxic T cells and antibodies. MHC genes are highly polymorphic, resulting in thousands of variants in the population. After organ transplantation, MHC mismatches between donor and recipient are the main cause for graft rejection and dysfunction. Allogeneic cellular and humoral immune responses specific for mismatched HLA class I and class II proteins were shown to play a major role in the development of chronic rejection pathology, which is known to be the main obstacle to long-term graft survival. 27 Furthermore, in fully MHC matched pairs, rejection regularly occurs due to differences in the presented peptides (mHag).
Previously, the feasibility of stably downregulating MHC class I and class II antigens in different cell types and in the original 3D-structure of anatomical simple tissues such as the cornea was demonstrated. It was shown in vitro and in vivo that the downregulation of MHC expression generates a condition of immunological invisibility, which prevents the rejection of allogeneic cells after transplantation in MHC-incompatible settings. MHC-silenced cells prevented de novo alloimmune responses and protected the allogeneic cells against pre-formed humoral immune responses in vivo. 8 –12
The graft's vascular endothelium constitutes the first and most relevant immune recognition interface between the donor and recipient. The endothelium orchestrates the allogeneic innate and adaptive immune responses, which center around MHC proteins and the peptides presented as mHags. The mECs constitutively express MHC class I and class II, which initiate the immune recognition process and are targeted by de novo or pre-formed DSA as well as memory T cells. 20 Therefore, the organ endothelium, representing the most relevant immunologic interface in allogeneic transplantation, may be considered the major target for genetic engineering approaches aiming to improve graft survival. 28
The lung has already been used in in vivo and ex vivo gene therapy due to its ease of accessibility via the airways. Although hereditary diseases (e.g., cystic fibrosis and α1-antitrypsin deficiency) have attracted major attention, other experimental genetic engineering approaches for acquired diseases (e.g., asthma, allergic rhinitis, and cancer) have also been evaluated. However, such approaches have envisioned the organ epithelium as the therapeutic destination, while the endothelium has been so far difficult to manipulate during in vivo attempts. 29 In addition, the arterial system has been avoided as the delivery route in order to prevent undesirable vector-associated immune responses and off-target effects. 30 Lung gene therapy has mainly focused on adenoviruses (AdV) or adeno-associated viruses (AAV) as gene therapeutic vectors, though the expression of transgenes delivered by AdV or AAV is only transient. 31 –35 However, large numbers of lentiviral vector particles are more difficult to produce than AdV or AAV and could be associated with more severe off-target effects when applied in vivo. Thus, from the perspective of organ immune engineering, the lung poses several challenges due to its size and complexity.
From a clinical perspective, the long-term benefit of lung transplantation is hindered by chronic graft rejection and organ failure. 3,27 In contrast to other organs, spontaneous or induced operational tolerance remains elusive in lung-transplanted patients. 36 Hence, new therapeutic strategies are highly desirable to improve the long-term survival of pulmonary grafts. As MHC and MHC-mediated mHag mismatches are the main causes of allograft rejection and organ failure, this study was aimed at the permanent downregulation of MHC class I and class II expression in order to maintain a condition of future immunological invisibility during all time after transplantation. Recently, normothermic EVLP has become a well established tool originally designed to maintain organs after explantation in a warm, functioning state outside of the body to monitor and optimize their health. In addition, EVLP has even been shown to regenerate injured lungs toward expansion of organ utilization. 37 Although normothermic EVLP was initially developed to circumvent major drawbacks associated with static hypothermic preservation, this study evaluated the feasibility of genetically engineering lung endothelium by lentiviral transduction during normothermic ex vivo perfusion, as it enables the organ endothelium to be reached while preserving the organ.
Silencing MHC class I and class II expression in the lung endothelium is expected to decrease the organ's immunogenicity and support long-term graft survival. Importantly, as residual MHC class I expression might be required to prevent natural killer (NK) cell cytotoxicity, RNA interference technology was advisedly applied to silence MHC expression, instead of gene-editing technologies, in order to avoid complete MHC knockout. Previously, the necessity for residual HLA class I expression was demonstrated in human ECs to inhibit NK cell cytotoxicity. 11,12 Hence, in this study, a lentiviral vector was used for the delivery of shRNA targeting β2-microglobulin and CIITA for permanent downregulation of MHC class I and class II, respectively. Previously, it was shown that the downregulation of MHC antigens by 60% was sufficient to prevent allogeneic immune responses in vitro and in vivo. 8,38 However, future studies will be required to demonstrate if this MHC silencing effect on the endothelium is sufficient to prevent immune responses. Expression of the reporter gene NL in different representative regions of the right and left lung showed that the entire lung endothelial surface was exposed and transduced by the lentiviral construct. However, transductions efficiencies appear to vary between different regions, based on the bioluminescence levels detected on the cell culture of different lung regions. Nevertheless, no pattern of highly or less transduced regions was recognized, with exception of the middle lobe of the right lung, which showed higher luminescence levels in all lungs. These variations might be explained by differences in the perfusion efficiencies of the different regions, which might affect the capability of the vector to reach the different regions. In addition to the expression of the reporter gene NL, significant downregulation of SLA class I and class II transcripts in lung mECs of the different lobe areas confirmed the efficient transduction of the endothelium. Expression of both MHC class I and in particular class II is known to be strongly upregulated under inflammatory conditions, which increases the immunogenicity of the graft after allogeneic transplantation. 39 This study used IFN-γ and TNF-α to mimic inflammatory conditions. During inflammatory responses, IFN-γ and TNF-α are often simultaneously upregulated and regulate the expression of several genes such as MHC in a synergistically manner. 40 –44 Hence, a combination of both cytokines was used to simulate strong inflammatory conditions but without strongly increasing cell death (Supplementary Fig. S2). Under IFN-γ and TNF-α stimulation, mECs derived from transduced porcine lungs showed a substantially lower capacity to upregulate SLA class I and class II expression in comparison to mECs of non-engineered lungs. This indicates strong shRNA-mediated regulation of gene expression, even under strong inflammatory stimuli.
Previously, it was shown that EVLP may be used as a reconditioning strategy to monitor and improve the function of organs otherwise not suitable for transplantation. 45,46 Therefore, it is important that the use of gene therapeutic vectors does not compromise the beneficial EVLP effect. LDH has been used as a marker for cell and tissue damage. 47,48 Comparison of LDH levels in the perfusate of transduced lungs to those detected in non-manipulated organs indicates that the vector does not induce substantial tissue injury, as shown by the slight increase in LDH. Also, histological analyses did not reveal additional damage caused by the perfusion with lentiviral vectors. A fine-tuned balance between pro- and anti-inflammatory cytokines is crucial for wound healing after injury, as well as for the modulation of allogeneic immune responses. It has been demonstrated that EVLP induces, among other inflammatory mediators, 49 the secretion of pro-inflammatory cytokines such as IL-1β, IL-6, IL-8, MCP-1, and G-CSF in the lung tissue, even though this was within acceptable ranges for transplantation. 50 The impact of the increase in cytokine secretion during EVLP for graft survival after transplantation remains unclear. However, recent studies have already shown approaches to eliminate the negative effects of those cytokines produced during EVLP. 51 In the porcine model used in this study, IL-6, IL-8, and TNF-α were only detectable in the lungs perfused with lentiviral vectors, but at lower levels than those previously reported for conventional EVLP. 49 –51 However, the secretion of those cytokines may indicate activation of the endothelium, and its effect post transplantation will have to be investigated.
Altogether, the feasibility of efficiently engineering the lung endothelium during normothermic EVLP has been demonstrated. The use of lentiviral vectors permits permanent or conditional regulation of gene expression, which is of the utmost importance, as only the short ex vivo period between transplant procurement and recipient transplantation provides the unique opportunity for organ engineering without accepting systemic off-target effects. The use of ex vivo perfusion to deliver the lentiviral vector has been shown to be a powerful tool to circumvent the limitations associated with the production of a high amount of lentiviral vector particles required for the modification of large organs. The lung was chosen to demonstrate the feasibility of genetic engineering in this most challenging organ because of its extraordinary size, low ischemic tolerance, and complexity. Ex vivo perfusion is currently used in kidney, lung, liver and heart transplantation and should be considered as offering a realistic option for genetic engineering. In particular, reducing graft immunogenicity by silencing MHC class I and class II antigen expression may allow the generation of immunologically invisible organs and may become an important future strategy to improve graft survival and combat the burden of rejection and immunosuppression.
Footnotes
Acknowledgments
We are thankful to Susanne Aufderbeck for her excellent technical assistance.
Author Disclosure
The authors do not have conflict of interests.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.