
Abstract
Today, there are >500 published studies and 40 clinical trials to treat retinal disorders using gene therapy. The great majority of them rely on the use of adeno-associated virus vectors (AAV) for therapeutic gene delivery. Thus far, AAVs have an excellent safety profile in the clinic. Nevertheless, it is known that AAV-mediated gene delivery leads to toxicity at higher input doses in experimental gene therapy. This study reveals the factors that contribute to retinal toxicity after subretinal administration of AAV vectors in wild-type mice. The study shows that alongside the input dose, the nature of the transgene and the cells mediating the expression determine the extent of toxicity. Importantly, the study shows that AAV vectors encoding green fluorescent protein (GFP) used as controls in experimental gene therapy are toxic at doses as low as 5 × 109 vg, confounding the observed therapeutic effect in gene therapy paradigms. Altogether, the data show the importance of reducing input doses while increasing transgene expression levels via the use of more efficient capsids and promoters in order to avoid side effects in AAV-mediated gene therapy. Furthermore, the toxicity observed with AAV-GFP vectors imply a reinterpretation of previous gene therapy studies where the therapeutic effect was measured in relation to this control.
Introduction
Over the past 20 years, considerable effort has been invested in establishing safe and effective gene therapy approaches for a multitude of diseases. 1 Applications in the retina have been particularly successful, with positive outcomes and a lack of side effects confirming the suitability of the eye as a target organ. 2 The use of adeno-associated virus (AAV) vectors has contributed to the success of retinal gene therapy in the clinic. AAVs' ability to transduce a wide variety of tissues and mediate long-term expression of the transgene after a single in vivo administration has made it the vector of choice. Indeed, wild-type AAV is not associated with any disease pathology in humans and is also naturally replication defective, requiring a helper virus such as adenovirus to replicate. 3 AAVs are one of the simplest gene delivery vectors, containing only the transgene expression cassette flanked by two non-coding viral inverted terminal repeats (ITRs) enclosed in a capsid composed of three structural proteins: VP1, 2, and 3. 4 The simplicity of AAV vectors and their relatively low efficiency in transducing professional antigen presenting cells (APCs) (e.g., macrophages or dendritic cells) 5 –7 likely contribute to their generally low immunogenicity. 8 Moreover, transferred genomes tend to persist inside the cells primarily in an episomal, non-integrated form, reducing the risks of insertional mutagenesis. 9 All of these favorable properties account for the success of AAV in gene therapy thus far.
The great majority of clinical trials thus far have targeted the retinal pigment epithelium (RPE) cells, which are easily and efficiently transduced with vectors administered into the subretinal space. Indeed, RPE cells perform phagocytosis as one of their primary functions, and their AAV-mediated transduction is aided by the phagocytic properties of the RPE. Today, more translational studies are being undertaken targeting multiple retinal cell types, and there is a noticeable increase in the number of clinical trials being prepared, with diverse strategies from gene replacement 10 –14 to neuroprotection 15 to optogenetics, all of them using AAV as a vector. The nature of therapeutic proteins being expressed in these newer applications, however, demands higher-level gene expression in more difficult-to-transduce cell types than the RPE. These new studies are likely to put higher demands on AAV as a gene delivery vehicle, and this will naturally lead to the use of higher input doses. In increasing AAV input doses, several potential obstacles need to be taken into account. These include but are not limited to: (1) phenotoxicity, that is, problems arising from either overexpression or ectopic expression of the transgene; and (2) immunotoxicity, that is, harmful immune responses to either the capsid and/or to the transgene product. 16,17
Thus far, there have been a few studies reporting AAV gene delivery–related side effects in the retina, 12,18 –20 but the vector components that are accountable for the toxicity have not been investigated individually. This study probed the interplay between the viral dose, the viral capsid, the encoded transgene, and the promoter driving transgene expression in mediating toxic effects. It shows that the toxicity is directly correlated with the injected dose and can be entirely avoided at low doses. However, it has been seen that AAV vectors encoding green fluorescent protein (GFP) are toxic to cells at doses as low as 5 × 109 particles per eye—a dose frequently used in experimental gene therapy studies. 21 –32 The study also shows that toxicity is dependent on the transgene as a retinal protein (retinoschisin) delivered through AAV at the same dose led to less toxicity than GFP. These findings indicate that AAV-GFP vectors regularly used as controls in gene therapy studies can be toxic, creating a bias for the eye treated with the therapeutic transgene, urging a re-evaluation of some of the past studies. 25,28,29 The GFP-related phenotoxicity reported here should be taken into account in the design of future experimental gene therapies. AAV capsid protein is responsible for adverse effects, as non-coding AAV capsids caused toxicity at high doses. Therefore, the use of engineered AAVs with better infectivity and strong cell type–specific promoters should be favored over the use of higher doses with ubiquitous promoters.
Methods
AAV production
AAV vectors were produced as previously described using the co-transfection method and purified by iodixanol gradient ultracentrifugation. 33 AAV vector stocks were titered by quantitative polymerase chain reaction (PCR) 34 using SYBR Green (Thermo Fisher Scientific).
Animals and intraocular injections
The experiments were performed in accordance with the National Institutes of Health Guide for Care and Use of Laboratory Animals and approved by local ethics committees. Wild-type C57BL6/j mice (Janvier Labs) were used for this study. For ocular injections, mice were anesthetized by isoflurane inhalation. Pupils were dilated, and a 33-gauge needle was inserted into the eye to deliver 1 μL of AAV vector solution subretinally. Ophthalmic ointment (Fradexam) was applied after surgery. Eyes with extensive subretinal hemorrhage were excluded from the analysis. GFP expression was visualized using a Micron III ophthalmoscope after dilation of the pupils and under isoflurane anesthesia.
Electroretinogram recordings
Mice were dark adapted overnight and then anesthetized by intraperitoneal injection of ketamine (Kétamine 500; Virbac) and xylazine (Xylasine 2%; Rompun). Pupils were dilated, and mice were placed on a 37°C heated pad. Electrodes with contact lenses were positioned on the cornea of both eyes. A reference electrode was inserted into the forehead and a ground electrode into the back. For recording of scotopic electroretinograms (ERGs), stimuli range was between 0.1, 1, and 10 cd × s/m 2 . For recording of photopic ERGs, mice were initially exposed to a rod-saturating background for 10 min. Stimuli range was between 1, 10, and 30 cd × s/m 2 and was presented 60 times on a rod-saturating background. Data were analyzed with Espion ERG software, and then with GraphPad Prism.
Histology, immunohistochemistry, and microscopy
Mouse eyes were enucleated and immediately fixed in 10% formalin/4% formaldehyde for 2 h. After multiple washes and removal of the anterior parts, eyecups were immersed in phosphate buffered saline (PBS)/30% sucrose overnight at 4°C. Afterwards, they were embedded in optimal cutting temperature medium and frozen in liquid nitrogen. Vertical sections 10 μm thick were cut with a Microm HM560 cryostat (Thermo Fisher Scientific). After incubation in a blocking buffer, sections were incubated with primary antibodies overnight at 4°C: Iba-1 antibody (019-19741; Wako) and GFAP antibody (G3893; Sigma–Aldrich). After multiple washes of the sections, the secondary antibodies (Alexa Fluor 546 and 647; Thermo Fisher Scientific) were added for 2 h at room temperature, followed by several washes. Retinal cryosections were mounted in Vectashield mounting medium (Vector Laboratories) for fluorescence microscopy and visualized using an Olympus confocal microscope.
Results
Toxicity thresholds for subretinally administered AAV serotype 8 vectors encoding GFP
Subretinal injections have been the preferred administration route for gene delivery to photoreceptors and the RPE. Input dose increase is often envisioned to increase therapeutic gene expression in gene therapy settings. To investigate the possible toxicity of vectors as a function of input dose, an AAV serotype 8 (AAV8) vector was used, which is frequently used for targeting photoreceptors. The vector was produced at high titer and encoded GFP under a ubiquitous self-complementary (sc)CAG promoter. Subretinal injections were performed bilaterally at a dose of 5 × 1011 vg/eye in 1 μL volume in five wild-type C57BL6/J mice. Reporter gene expression was monitored on a weekly basis with fluorescent eye fundus imaging. High-level GFP expression was visible in all animals 2 weeks post dosing (Fig. 1A). Interestingly, GFP expression decreased, starting at 3 weeks, as seen on the eye fundus images (Fig. 1A and B and Supplementary Fig. S1; Supplementary Data are available online at

Effect of AAV8-scCAG-GFP injection on retinal structure and function.
AAV capsids and transgene overexpression together cause toxicity
The toxicity occurring at high AAV input doses can be attributed to the capsid and/or to the transgene product. To understand the causes of the observed phenotoxicity better, equal dose unilateral injections were performed with an AAV8 vector encoding GFP or the same vector where the transgene is oriented in reverse orientation in between loxP sites. This AAV (referred to as AAV8-flox) can only lead to expression in cells expressing Cre recombinase and therefore served as non-coding control in the experiments. About a 30% ERG amplitude decrease was observed with the AAV8-flox vector, suggesting that the capsid alone is responsible for a significant part of the toxicity. ERGs became un-recordable with an equal amount of AAV8-GFP, suggesting that the combination of high amounts of AAV capsid proteins and GFP is severely toxic to the retina.
Since GFP is a protein not normally found in the mammalian retina, the study investigated whether a retinal protein would also induce toxicity at high doses. To this aim, a high dose (1 × 1011 vg) of the same AAV vector encoding retinoschisin, a protein already expressed in the retina, was injected. It was found that there was about a 65% ERG decrease with high-dose AAV8 encoding retinoschisin (Fig. 2A). This suggests that even proteins normally expressed in the retina lead to toxicity when overexpressed or expressed in other cell types than those that usually produce the protein. Nevertheless, retinal proteins led to less severe toxicity compared to GFP, as shown by recordable ERGs in retinas overexpressing retinoschisin.
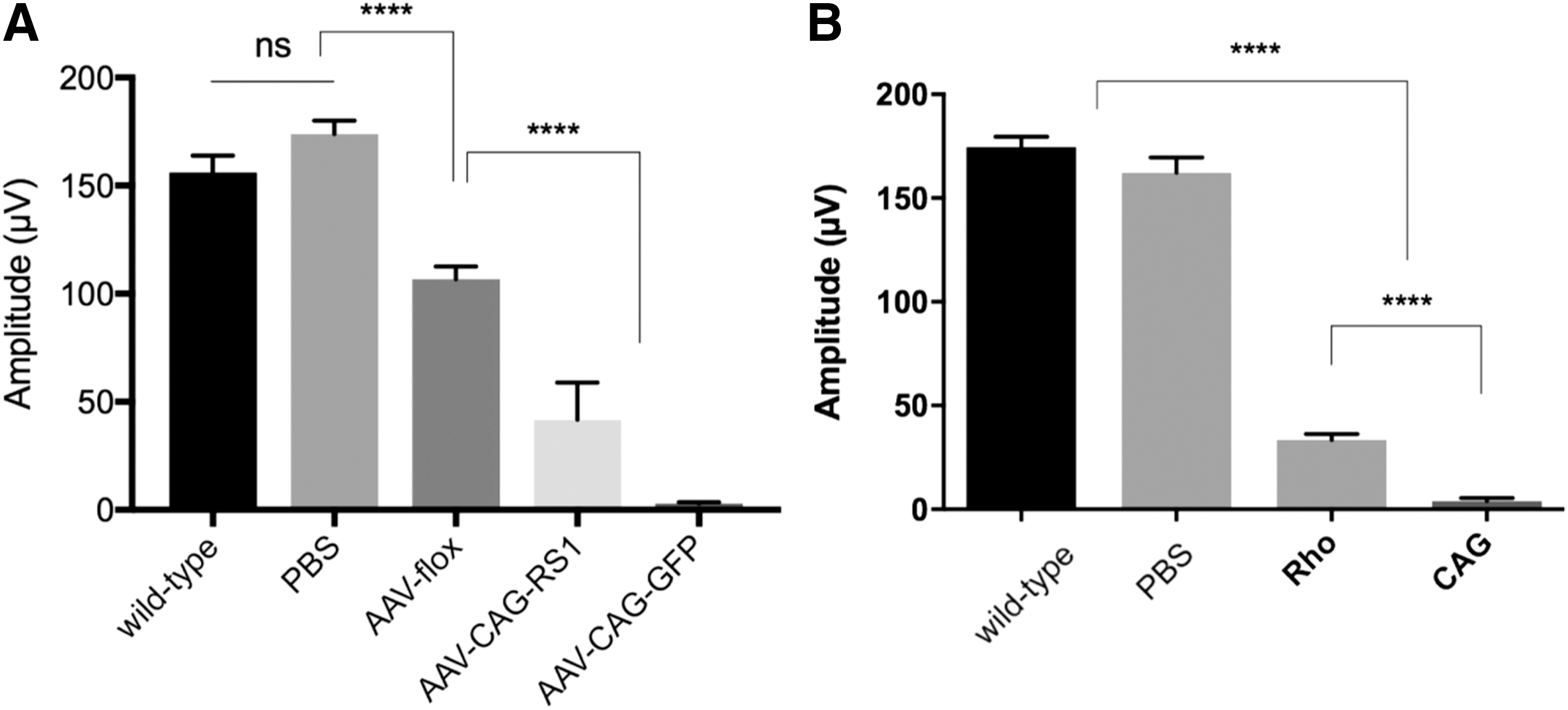
Functional retinal defects caused by high input AAV doses.
Since the cell types expressing GFP can play a role in immunotoxicity (i.e., harmful immune responses to GFP), the study then investigated whether a cell type–specific promoter could influence the toxicity. Expression of GFP was compared under two strong promoters: the ubiquitous CAG promoter versus the rod-specific rhodopsin promoter (1 × 1011 vg). Interestingly, GFP expression under the rhodopsin promoter was stronger (Supplementary Figs. S4 and S5), but GFP expressed under CAG promoter led to significantly more toxicity (Fig. 2B).
Local immune responses to high input doses of AAV-GFP
The toxicity observed in the current experiments can be attributed to immunotoxicity, that is, harmful immune responses to the capsid and the transgene product. Using AAV8-scCAG-GFP vector, GFP expression was found in the RPE, photoreceptors, and Müller glial cells involved in mediating immune reactions. Thus, the study investigated whether local immune responses were elicited within the retina. Two months after subretinal injections of AAV8-scCAG-GFP, retinal cryosections were prepared, and positive immunostainings were found for GFAP, a marker for Müller glial cell activation, and Iba-1, a marker for microglial cell activation. No labeling was observed with either marker in PBS-treated retinas (Fig. 3).

Local immune reactions to high dose AAV-GFP. Representative retinal cryosections showing GFAP and Iba1 immunostainings after subretinal injection of
Discussion
Gene therapy holds promise for treating patients with inherited diseases. It is generally agreed that AAVs have an excellent safety profile, are nontoxic, and display low immunogenicity. Nevertheless, preclinical and clinical experience has shown that AAV components can be recognized by the host immune system. 35 The extent of the impact of potential toxicity or inflammation on therapeutic efficiency remains poorly understood. In this context, it is important to investigate what aspects of AAV vectors can induce toxic effects in order to control them in gene therapy protocols. Indeed, there are now a few reports supporting acute toxicity from AAV at higher input doses, such as injections in the brain 36,37 or systemic injections to treat liver disorders, 38 but also when transducing embryonic stem cells in vitro, likely through disruption of DNA damage pathways. 39 Mechanism of AAV-induced toxicity is likely distinct for each scenario.
The present study showed that AAV is toxic to the retina at high input doses. High doses of AAV capsids without any transgene expression led to retinal toxicity, suggesting that the AAV capsid by itself accounts for part of the toxicity. Vectors encoding GFP were significantly more toxic, suggesting both the capsid and the transgene play a role in toxicity. Based on these data, GFP-encoding vectors should be carefully dosed when used as controls in gene therapy experiments, as high-level GFP expression in control retinas can give rise to a false therapeutic effect by increasing the difference between control and treated groups. The nature of the transgene also plays a role in toxicity, as a mammalian retinal protein led to less severe toxicity than foreign GFP. Interestingly, there was no direct correlation between the amount of GFP expression and the extent of toxicity, as restricting GFP expression to rods diminished toxic effects, despite providing higher expression levels. This suggests there is not only phenotoxicity due to overexpression but also that ectopic expression plays an important role. There are reports suggesting that the immune system plays a role in the toxic side effects observed at higher doses, 20 with cellular responses mediating clearance of the transduced cells. 35
One limitation of this study is the use of mice, as nonhuman primates (NHPs) are better models to assess immunogenicity and toxicity toward AAV. However, even in this rodent model that is presumed to tolerate intraocular AAV injections, dose-dependent structural and functional retinal defects were found, suggesting that more severe reactions could occur in primates. 18,20 Nevertheless, this study adds to the understanding of which AAV-related components correlate with side effects and informs the dosage thresholds to respect and factors to take into account in order to avoid toxicity in AAV-mediated gene therapy.
Several studies suggest dosing thresholds to maintain safety. 18,20 However, since there are numerous parameters that play a role on the efficacy of transduction and transgene expression—such as the promoter, the capsid, and other cis regulatory elements—it is difficult to define one single dose for all retinal gene therapy settings. The threshold dose needs to be determined for each vector and target tissue as a function of the administration route and animal model used. It has been shown in NHP studies that intravitreal injections of ≥1 × 1012 vg/eye is harmful and inflammatory when associated with GFP and ubiquitous promoters. 12,19 Subretinally, the threshold seems to be lower (around 1 × 1011 vg/eye in NHPs), 10,18,19 as higher doses were associated with inflammation, 20 also in patients, 40 likely because the vector is directly in contact with the target cells. In mice, the dose range is usually between 1 × 108 and 1 × 1010 vg. 10,41,42
Dose sparing using enhanced AAVs that allow low particle numbers to be used while maintaining satisfying expression levels is an attractive strategy to alleviate AAV-related toxicity issues. 43 Engineered capsids can be combined with cell type–specific promoters that restrict transgene expression to target cells, further refining the safety and efficacy of gene therapy. 43,44
Footnotes
Acknowledgments
This study was supported by Marie Curie CIG (334130, Retinal Gene Therapy), INSERM, Labex-Lifesenses (ANR-10-LABX-65), the Agence Nationale pour la Recherche—Recherche Hospitalo-Universitaire en santé (RHU; Light4Deaf), Fondation NRJ. H.K. was supported by a PhD fellowship from the AFM-Téléthon. We thank Julie Vendomèle for helpful discussion and advice. We thank Céline Winckler and Camille Robert for assistance with the production of plasmids and AAV vectors.
Author Disclosure
H.K. and D.D are inventors on two pending patent application on noninvasive methods to target cone photoreceptors using novel adeno-associated virus variants. D.D. is an inventor on a patent on adeno-associated virus virions with variant capsid and methods of use thereof with royalties paid to Avalanche Biotech. D.D. is a consultant for GenSight Biologics. C.C., L.P., and S.F. report no conflict of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.