
Abstract
Diabetes mellitus is associated with increased risk of heart failure. It has been previously demonstrated in mice that a single injection of adeno-associated virus 8 encoding urocortin 2 (AAV8.UCn2) increases glucose disposal in models of insulin resistance and improves the function of the failing heart. The present study tested the hypothesis that UCn2 gene transfer would reduce diabetes-related left ventricular (LV) dysfunction. Eight-week-old C57BL6 male mice were fed a Western diet (WD; 45% fat, 35% carbohydrate) for 40 weeks. At week 30, they received saline or AAV8.UCn2 (2 × 1013 genome copies/kg) via intravenous injection. Ten weeks after gene transfer, fasting blood glucose, glucose tolerance, and cardiac function were measured via echocardiography and in vivo measurement of LV contractile function, and the results were compared to those of mice fed normal chow (NC; 10% fat; 70% carbohydrate). The contents of key LV signaling proteins were also measured to probe mechanisms. WD increased 12 h fasting glucose (WD: 190 ± 11 mg/dL, n = 8; NC: 105 ± 12 mg/dL, n = 7; p = 0.0004). WD tended to reduce LV peak +dP/dt (p = 0.08) and LV peak –dP/dt (p = 0.05). LV ejection fraction was unchanged. Among WD-fed mice, UCn2 gene transfer reduced 12 h fasting glucose (WD-UCn2: 149 ± 6 mg/dL, n = 8; WD-Saline: 190 ± 11 mg/dL, n = 8; p = 0.012), increased LV peak +dP/dt (p < 0.001) and LV peak –dP/dt (p = 0.013), and reduced Tau (p < 0.02), indicating beneficial effects on systolic and diastolic LV function. In addition, among WD-fed mice, UCn2 gene transfer increased LV ejection fraction (p < 0.005) and the velocity of circumferential fiber shortening (p = 0.0005). Finally, a reduction was seen in fatty infiltration of the liver in WD-fed mice that had received UCn2 gene transfer. LV samples from WD-UCn2 mice showed increased phosphorylation of the protein kinase A catalytic domain (p = 0.03). In conclusion, UCn2 gene transfer increased LV systolic and diastolic function and reduced blood glucose in mice with diabetes-related LV dysfunction, indicating that UCn2 gene transfer may be of potential therapeutic benefit.
Introduction
Diabetes mellitus affects 9% of adults worldwide 1 and increases the risk of developing cardiovascular diseases. In addition, among adults with diabetes, heart disease death rates are two to four times higher. 2,3 Left ventricular (LV) dysfunction often accompanies diabetes, even in the absence of obstructive coronary atherosclerosis or hypertension. 4 Structural alterations of the LV associated with diabetes-related LV dysfunction often include increased wall thickness and fibrosis accompanied by diastolic or systolic dysfunction. 4,5 Uncontrolled high blood glucose, insulin resistance, and impaired cardiac insulin signaling contribute to the development of diabetes-related LV dysfunction. 6 Despite advances in understanding its pathophysiological mechanisms, diabetic-related LV dysfunction represents an unmet medical need, warranting development of new therapeutic approaches.
The mammalian corticotropin-releasing factor (CRF) family peptides consist of CRF and urocortins 1, 2, and 3. These hormonal peptides regulate stress responses and energy balance by activating their cognate receptors, corticotropin-releasing hormone receptor type 1 (CRHR1) and 2 (CRHR2), which are expressed in the central nervous system and periphery. 7,8 Among the CRF family peptides, UCn2 and UCn3 share 42% homology in amino acid sequence, and in vitro pharmacokinetic studies have demonstrated that both peptides preferentially bind to CRHR2 with small differences in receptor affinity. 9 –11 In preclinical and clinical studies, peptide infusion of UCn2, UCn3, and the related stresscopin showed beneficial cardiovascular effects in animal models and in clinical heart failure. 12 –16 Recent studies showed that sustained elevation of plasma UCn2 or UCn3 via intravenous (i.v.) delivery of an adeno-associated virus type 8 (AAV8) vector encoding UCn2 or UCn3 increases LV contractile function in mice. 17 –19 Interestingly, UCn2 and UCn3 have dissimilar effects on glucose metabolism. For example, UCn2 gene transfer increases insulin sensitivity and decreases fasting blood glucose in both normal and insulin-resistant mice, but UCn3 gene transfer has no effect on glucose disposal. 19,20 These results indicate that UCn2 and UCn3 elicit distinct physiological responses, despite similar selectivity and binding affinity to CRHR2. Furthermore, UCn2 is a unique CRF family peptide that can improve LV function as well as glucose homeostasis.
The present study investigated the effects of UCn2 gene transfer on mice with diet-induced abnormalities of LV function. The hypothesis that UCn2 gene transfer would attenuate the deleterious effects of a Western diet (WD) on LV function was tested.
Methods
Animals and protocol
Twenty-three C57BL6 male mice (8 weeks old) were obtained from the Jackson Laboratory (Bar Harbor, ME). Mice were allocated to two groups and provided with cereal-based normal chow (NC; control group, n = 7; Teklad laboratory diet; Envigo, Huntingdon, United Kingdom) or a higher-fat diet (n = 16; cat. #D12451, Research Diets, Inc., New Brunswick, NJ). The higher-fat diet has been referred to as a WD 21 because it is similar in composition to the diets of people in Western countries (kcal: fat 45%; carbohydrate 35%; protein 20%) and is higher in fat and lower in carbohydrate than NC (kcal: fat 10%; carbohydrate 70%; protein 20%) but lower in fat than the standard high-fat diet often used to induce insulin resistance in mice. 20
Mice were fed the WD for 40 weeks. At 30 weeks, WD-fed mice received i.v. saline (n = 8) or i.v. AAV8.UCn2 (2 × 1013 genome copies [gc]/kg; n = 8). Glucose homeostasis and cardiac function were measured 10 weeks after randomization (Fig. 1A) at the end of the 40-week period. A control group of mice fed NC was included for comparison. The primary test for efficacy was a group difference in WD-fed mice that received UCn2 gene transfer compared with the group that received saline. Data acquisition and analysis were blinded to the treatment group. The use of animals was in accordance with the Association for Assessment and Accreditation of Laboratory Animal Care guidelines and was approved by the Institutional Animal Care and Use Committee of VA San Diego Healthcare System.

Experimental protocol, adeno-associated virus 8 encoding urocortin 2 (AAV8.UCn2) plasma UCn2 after gene transfer.
AAV8 production and gene transfer
The construction and production of AAV8 encoding murine UCn2 with a chicken β-actin promoter (Fig. 1B) have been described previously. 18,19 Mice received 2 × 1013 gc/kg of virus diluted in saline (100 μL) through the jugular vein under anesthesia.
Plasma UCn2
Plasma was collected after a terminal study (10 weeks after gene transfer) and stored at −80°C. Plasma UCn2 levels were measured using mouse UCn2 enzyme immunoassay kits (cat. #KT-376; Kamiya Biomedical Co., Tukwila, WA) as instructed by the manufacturer.
Blood glucose and glucose tolerance testing
Mice were fasted for 12 h but were provided water ad libitum. Blood was drawn from a small tail incision, and glucose was measured using a Contour Blood Glucometer and test strip (Bayer [Ascensia Diabetes Care], Parsippany, NJ). After measuring 12 h fasting glucose, mice received glucose (2 g/kg) via an intraperitoneal (i.p.) injection, and blood glucose was measured every 30 min for 120 min.
Echocardiography
Mice were maintained in a lightly anesthetized state using 1% isoflurane. A Vevo 2100 system (FUJIFILM VisualSonics, Inc., Toronto, Canada) was used to assess chamber dimensions, ejection fraction (EF), and velocity of circumferential fiber shortening corrected for heart rate (VCFc), as previously described. 20 Vevo LAB v2.1 (VisualSonics) was used for measurement and analysis.
LV systolic and diastolic function
Mice were anesthetized with sodium pentobarbital (80 mg/kg, i.p.) and mechanically ventilated with oxygen (1 L/min). The right carotid artery was exposed to insert a 1.4F pressure-conductance transducer into the LV chamber followed by vagotomy, and LV pressure was measured. Two animals were excluded from the analysis due to abnormal responses during anesthesia, one each from the WD-Sal and WD-UCn2 groups. After the procedure, mice were killed by exsanguination under deep anesthesia with sodium pentobarbital (200 mg/kg), and blood and organs were collected.
Immunoblotting
LV samples were homogenized in 1 × lysis buffer (Cell Signaling Technology, Danvers, MA) containing protease and phosphatase inhibitors (Roche, Basel, Switzerland). Homogenates were sonicated on ice three times for 5 s each, and insoluble lysates were removed by centrifugation (13,000 × g for 10 min at 4°C). A total of 20–50 μg proteins were loaded per well and separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis. Separated proteins were transferred to a polyvinylidene fluoride membrane. After blocking with 5% nonfat dried milk in TBS-T for 1 h, the membrane was incubated with primary antibody diluted in TBS-T plus 5% bovine serum albumin overnight at 4°C. The primary antibodies used were: phospho-RYR S2808 (1:1,000; Abcam, Cambridge, United Kingdom), SERCA2a (1:1,000; Thermo Fisher Scientific, Waltham, MA), phospho-PLB S16 (1:5,000; Badrilla, Leeds, United Kingdom), phospho-TnI S23/24 (1:1,000; Cell Signaling Technology), phospho-PKA C T179 (1:1,000; Cell Signaling Technology), phospho-AKT S473 (1:1,000, Cell Signaling Technology), phospho-AMPKa T172 (1:1,000; Cell Signaling Technology), phospho-S6 S240/244 (1:1,000; Cell Signaling Technology), phospho-4EBP1 T37/46 (1:1,000; Cell Signaling Technology), and Vinculin (1:100,000; Sigma–Aldrich, St. Louis, MO).
Histology
Samples of LV and liver were fixed in 10% formalin overnight and then paraffin-embedded. Five-micron sections were slide mounted and stained with hematoxylin and eosin and Masson's trichrome. Microscopic images were scanned using an Axio slide scanner (Carl Zeiss, Oberkochen, Germany), and images were analyzed using ImageJ (NIH, Bethesda, MD).
Statistical analysis
All data acquisition and analyses were performed without knowledge of group identity. GraphPad Prism v7 (GraphPad Software, Inc., La Jolla, CA) was used for statistical analysis and calculation of area under the curve for data from glucose tolerance tests. Between-group comparisons were performed using Student's t-test (unpaired and two-tailed). Multiple tests were corrected by the Bonferroni method. The null hypothesis was rejected when p < 0.05.
Results
Morphometrics
Forty weeks after initiation of the dietary intervention (10 weeks after randomization to the UCn2 gene transfer group or the saline group), the mean body weight of the WD-Sal group had increased by 63% compared to the NC-Con group (Fig. 2A; p < 0.0001). Absolute (non-normalized) measures included: LV weight (NC-Con: 104 ± 3 g, n = 7; WD-Sal: 112 ± 3 g, n = 8, p = 0.07 vs. NC-Con; WD-UCn2: 109 ± 6 g, n = 8); liver weight (NC-Con: 1,466 ± 59 g, n = 7; WD-Sal: 2,571 ± 226 g, n = 8, p = 0.007 vs. NC-Con; WD-UCn2: 2,283 ± 197 g, n = 8); tibial length (NC-Con: 18.3 ± 0.1 mm, n = 7; WD-Sal: 18.2 ± 0.2 mm, n = 8; WD-UCn2: 18.1 ± 0.04 mm, n = 8). LV weight–tibial length ratio was increased 9% in WD-Sal mice compared to NC-Con (Fig. 2B; p = 0.03). Liver weight–tibial length ratio increased by 78% in WD-Sal mice compared to NC-Con (Fig. 2C; p = 0.0006). No group differences were seen in WD-fed mice that received UCn2 gene transfer compared to those receiving saline in terms of body weight, liver weight, or LV weight (Fig. 2A–C).

WD-fed mice: effects of UCn2 gene transfer on body, LV, and liver weight and glucose disposal.
Plasma UCn2
Plasma UCn2 concentration increased >20-fold 10 weeks after AAV8.UCn2 delivery (Fig. 1C; saline: 1 ± 0.2 ng/mL, n = 7; UCn2: 21 ± 2 ng/mL, n = 8; p < 0.0001).
Glucose disposal
Twelve-hour fasting glucose was elevated in mice on the WD (NC-Con: 105 ± 12 mg/dL; WD-Saline: 190 ± 11 mg/dL; p = 0.0004) and was reduced 10 weeks after UCn2 gene transfer (Fig. 2D; p = 0.012). Glucose tolerance testing showed decreased glucose disposal in WD-Sal compared to NC-Con (p < 0.003). WD-Sal mice showed impaired glucose tolerance compared to NC-Con mice (Fig. 2E; p < 0.003), and WD-fed mice that received UCn2 gene transfer tended to clear glucose more efficiently compared to WD-Sal mice (Fig. 2E; p < 0.06).
Echocardiography
Echocardiography measures showed no differences between NC-Con and WD-Sal mice. However, 10 weeks after randomization to UCn2 gene transfer or saline, group differences were evident. For example, in mice that received UCn2 gene transfer, EF (p < 0.005) and VCFc (p = 0.0005) were higher, and end diastolic diameter (p = 0.01) and end systolic diameter (p = 0.004) were lower compared to WD-fed mice that received saline (Fig. 3A–E).

Echocardiography.
LV function
Direct measurement of the peak rates of LV pressure development (+dP/dt) and decline (–dP/dt) showed that WD-Sal mice tended to have lower values (vs. NC-Con) in peak +dP/dt (Fig. 4A; a 17% decrease; p = 0.08) and in peak –dP/dt (Fig. 4B; a 19% decrease; p = 0.05). However, 10 weeks after randomization to either the UCn2 gene transfer group or the saline group, group differences were more striking. For example, in mice that received UCn2 gene transfer, peak +dP/dt (Fig. 4A; a 38% increase; p < 0.001) and peak –dP/dt (Fig. 4B; a 33% increase; p = 0.013) were higher compared to WD-fed mice that received saline. Tau was decreased (more rapid relaxation) in WD-fed mice that received UCn2 gene transfer (Fig. 4C; p < 0.02). No group differences were found in LV developed pressure or heart rate (Fig. 4D and E). LV end diastolic pressure tended to be lower in WD-fed mice that received UCn2 gene transfer (Fig. 4F; p < 0.06).
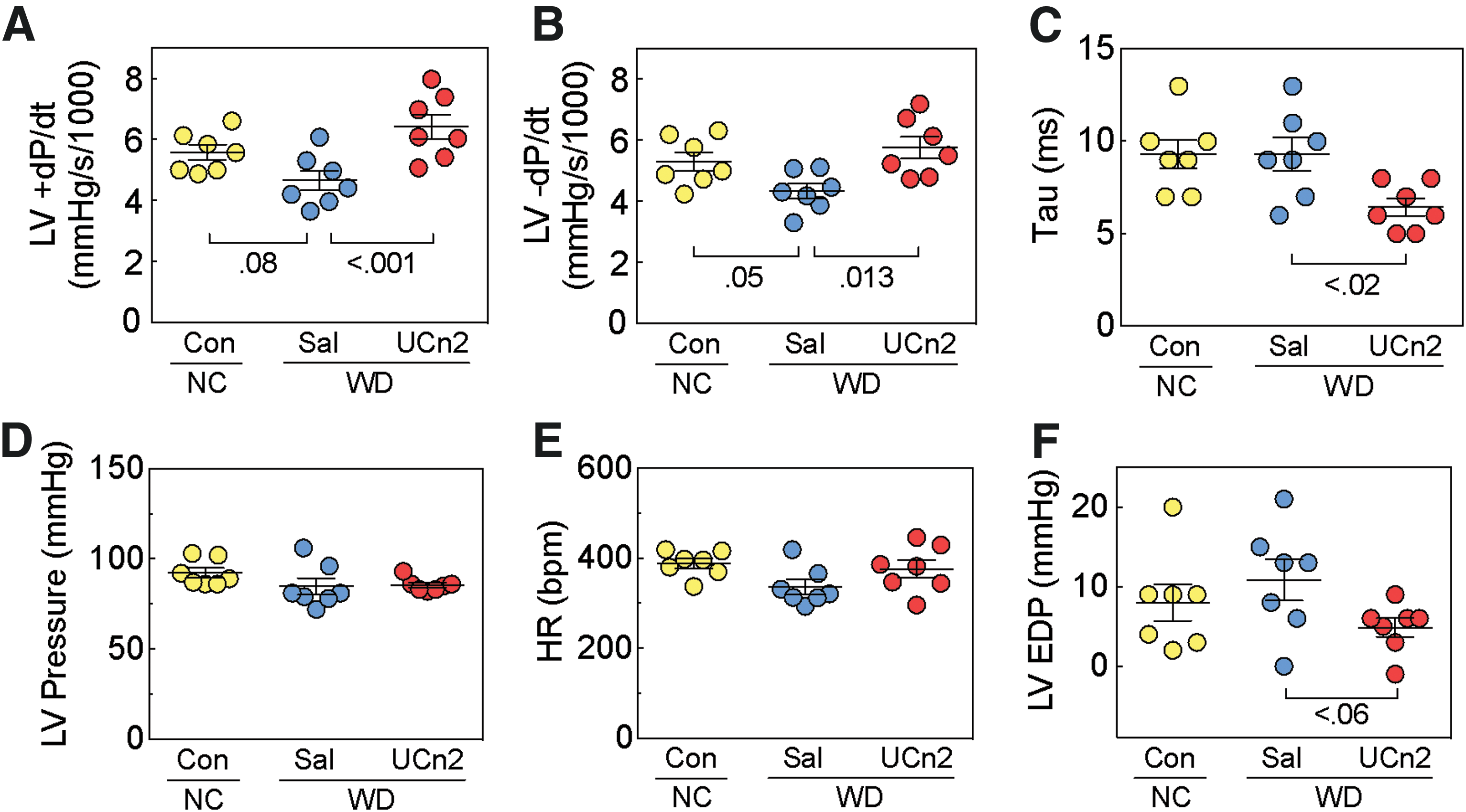
WD-fed mice: effects of UCn2 gene transfer on LV function.
Histological analysis
Liver fatty infiltration was compared by measuring the area of lipid droplets in the liver (Fig. 5A). WD-Sal mice showed increased fatty infiltration compared to NC-Con mice (p < 0.0002). WD-fed mice that received UCn2 gene transfer showed lower levels of fatty infiltration compared to WD-Sal mice (Fig. 5A; a 44% reduction; p < 0.008). Liver fibrosis was increased in the WD-Sal group compared to NC-Con mice (Fig. 5B; p < 0.02), and UCn2 gene transfer had no effect compared to WD-Sal mice. There were no group differences in LV fibrosis (Fig. 5B).

Histological analysis and immunoblotting.
Relevant LV signaling proteins
Immunoblotting was used to measure LV content of signaling proteins that govern LV contractility and cellular metabolism (Fig. 5C and Table 1). Phosphorylation at T179 in the protein kinase A (PKA) catalytic domain was increased in LV samples from WD-fed mice that received UCn2 gene transfer compared to WD-Sal mice (Fig. 5C and Table 1; p = 0.03). Phosphorylation at T172 in the AMPKα subunit was reduced in WD-fed mice that received saline compared with NC-fed mice (p = 0.022), and WD-fed mice that received UCn2 gene transfer tended to show increased phosphorylation of AMPKα (p = 0.13). No group differences were found in phospho-S2808 RYR2, SERCA2a, phospho-S16 PLB, phospho-S23/24 TnI, phospho-S473 AKT, phospho-S240 S6, or phospho-T37 4EBP1 (Fig. 5C).
Immunoblotting quantification
Data are the mean ± standard error. p-Values are from unpaired Student's t-tests (two-tailed).
p = 0.03 vs. WD-Sal.
p = 0.022 vs. NC-Con.
NC, normal chow; WD, Western diet; Con, control; Sal, saline; UCn2, urocortin 2.
Discussion
The most important finding of this study was that a single injection of AAV8.UCn2 increased LV function and glucose disposal in mice fed a higher-fat diet. The study tested whether UCn2 gene transfer would have beneficial effects on diabetes-related LV dysfunction. LV dysfunction was achieved using a higher-fat diet—one that is similar in fat content to a standard WD. 21 Mice that were fed this diet for 40 weeks (compared to NC with 10% fat) showed increased body weight and liver weight, fasting hyperglycemia, and impaired glucose tolerance. WD-fed mice tended to exhibit impaired rates of LV pressure development (+dP/dt) and impaired rates of LV pressure decline (–dP/dt) compared to mice fed NC. These relatively load-independent measures are sensitive and accurate measures of LV systolic and diastolic function. A major finding of this study was that WD-fed mice that received UCn2 gene transfer (compared to saline) had increased systolic and diastolic LV function 10 weeks later. This conclusion is based upon a blinded analysis of measures of glucose disposal and detailed analysis of cardiac function. These findings are important because the prevalence of diabetes-related LV dysfunction is increasing as the incidence of diabetes increases globally. 5 A one-time administration of an agent with enduring benefits on glucose disposal and LV function is an attractive and unique therapeutic strategy.
WD: glucose homeostasis and LV function
WD has previously been used to induce LV dysfunction in mice, 22,23 which was the rationale for using it. In the present study, mice fed WD for 40 weeks developed obesity, hyperglycemia, and impaired glucose disposal (Fig. 2). WD-fed mice also exhibited LV hypertrophy (see LV–tibial length ratio; Fig. 2B). Although LV EF showed no NC-Con versus WD-Sal group difference (Fig. 3A), LV peak +dP/dt and LV peak –dP/dt tended to be reduced (Fig. 4A and B). These less load-sensitive measures of LV function are superior to EF, an ejection phase index that is affected by loading conditions. 24
UCn2 gene transfer in WD-fed mice: glucose homeostasis
The mean plasma UCn2 concentration was 21 ± 2 ng/mL 10 weeks after AAV8.UCn2 delivery in WD-fed mice, a >20-fold increase compared to WD-fed mice that received saline (Fig. 1C). Increased plasma UCn2 in WD-fed mice was associated with reduced fasting blood glucose levels compared to WD-Sal mice (Fig. 2D; p = 0.012), and also tended to show improved disposal in glucose tolerance testing (Fig. 2E; p < 0.06). Hyperinsulinemic-euglycemic clamp studies would be required for a quantitative assessment of insulin sensitivity.
UCn2 gene transfer in WD-fed mice: LV function
It was previously demonstrated that UCn2 gene transfer increases LV function in normal mice 17 and also increases function of the failing heart (induced by myocardial infarction). 18 The present study found that UCn2 gene transfer has beneficial effects on LV function in mice with diabetes-related LV dysfunction induced by WD. WD-fed mice that received UCn2 gene transfer (WD-Sal) showed higher LV EF and VCFc (Fig. 3A and B). In addition, more detailed studies of LV function showed increased LV peak +dP/dt and LV peak –dP/dt in WD-fed mice that received UCn2 gene transfer (Fig. 4A and B), indicating important benefits on systolic and diastolic LV function. The relative increases of 38% and 33% on peak +dP/dt and peak –dP/dt, respectively, would be expected to have important physiological effects on heart function in clinical settings. Tau, the time constant of LV pressure decline, was decreased (i.e., it was more rapid) in WD-fed mice that received UCn2 gene transfer (Fig. 4C). These data indicate that UCn2 gene transfer reverses the decline in LV systolic and diastolic function seen in WD-induced diabetes-related LV dysfunction (Fig. 4A and B). The extent of LV hypertrophy, however, was not reduced by UCn2 gene transfer (Fig. 2B).
Diabetic-related LV dysfunction is associated with impaired insulin signaling, abnormal metabolic responses, and abnormalities in cardiac myocyte Ca2+ handling. 25,26 In vitro assays using isolated cardiac myocytes or cell lines showed that UCn2 peptide treatment increases cyclic adenosine monophosphate (cAMP) production, AMPK activity, phosphorylated phospholamban, and phosphorylated TnI via CRHR2 activation. 10,27 It has previously been shown that UCn2 gene transfer increases LV SERCA2a and improves Ca2+ handling in cardiac myocytes isolated from both normal and failing hearts of mice. 17,18 These studies also showed increased β-adrenergic receptor-stimulated cAMP production and PKA activity in cardiac myocytes isolated from normal mice that had received AAV8.UCn2, but not cardiac myocytes isolated from failing hearts. 17,18 The present study found that phosphorylation at T172 in the AMPKα subunit is decreased in LV homogenates from WD-fed mice that received saline compared to NC-Con mice, indicating reduced activation of AMPK signaling (Fig. 5C and Table 1). The phosphorylated PKA catalytic subunit was increased in LV homogenates from WD-fed mice that received UCn2 gene transfer (Fig. 5C and Table 1). There was no group difference in SERCA2a content in this study (Fig. 5C). Taken together, increased PKA activation may have contributed to increased LV contractility in WD-fed mice that received UCn2 gene transfer. Additional studies to determine the precise molecular mechanisms underlying these beneficial effects are underway.
Limitations
The favorable effects of UCn2 gene transfer on LV function in WD-fed mice likely require UCn2 activation of CRHR2. It was previously shown that deletion of CRHR2 abrogates the beneficial effects of UCn2 gene transfer on glucose disposal. 20 In that study, euglycemic clamps were used in insulin-resistant mice, and it was found that UCn2 gene transfer increases skeletal muscle insulin sensitivity and reduces hepatic glucose production. UCn2 increases Glut4 translocation in skeletal myotubes as effectively as insulin. 20 Whether the beneficial effects of UCn2 on LV function reflect improved glucose disposal and consequently a reduction of the deleterious effects of sustained hyperglycemia on LV function or a direct effect of UCn2 on Ca2+ handling in cardiac myocytes 17 –19 or both will require additional studies. However, there were additional benefits associated with reduced hyperglycemia following UCn2 gene transfer in the present study, including a 44% reduction in fatty infiltration of the liver (Fig. 5A; p < 0.008). Finally, a one-time therapy that improves both glucose disposal and LV function would be a welcome addition to current clinical therapies.
In conclusion, UCn2 gene transfer via a one-time i.v. injection of AAV8.UCn2 increased LV systolic and diastolic function, reduced blood glucose, and reduced fatty infiltration of the liver in mice with diabetes-related LV dysfunction, indicating that UCn2 gene transfer may be of potential therapeutic benefit in clinical settings.
Footnotes
Acknowledgments
We thank Eileen D'Souza for administrative support and the VA Histology Core for tissue processing. This study was supported by a grant from NHLBI (P01 HL066941) and Merit grants from the Department of Veteran's Affairs (1I01 BX001515-03 and 1I01 BX003774-01A1). Funding agencies played no role in the study design, conducting of the experiments, or interpretation of the data.
Author Disclosure
H.K.H. is cofounder and unpaid consultant of Renova Therapeutics. Renova did not provide financial support and was not involved in the studies. The other authors declare no competing interests.