
Abstract
(Pro)renin receptor (PRR) is a novel component of the renin–angiotensin system that has been demonstrated to be involved in cardiovascular diseases. Recent research reported that diabetic cardiomyopathy (DCM) may be accompanied by high expression of PRR, indicating that PRR may be a potential therapeutic target for DCM. However, the exact mechanisms of PRR in DCM have not been completely clarified. This study hypothesized that PRR is involved in the pathological progression of DCM and can exacerbate myocardial fibrosis and cardiac dysfunction. Inhibition of PRR expression may alleviate these pathological changes. In this study, in vivo experiments were performed in Wistar rats, and in vitro experiments were carried out in rat cardiac fibroblasts. After establishing an in vivo DCM model, the rats were divided into a control group, DCM group, adenovirus scrambled short hairpin RNA group, and adenovirus PRR short hairpin RNA group to observe further the effects of PRR RNA interference (RNAi) silencing on the pathogenesis of DCM. The results showed that PRR RNAi silencing decreased myocardial fibrosis and improved cardiac function in DCM. The study also observed the effects of PRR RNAi silencing on high glucose stimulated cardiac fibroblasts, and the results showed that PRR RNAi silencing inhibited the expression of type I collagen, type III collagen, and transforming growth factor beta. It was concluded that PRR plays a key role in the pathological progression of DCM and that inhibition of PRR expression achieved by specific PRR RNAi silencing offers a new therapeutic approach for DCM. The underlying mechanisms of these effects may be associated with the ERK signaling pathway and oxidative stress.
Introduction
Diabetic cardiomyopathy (DCM) is a major cardiac complication of diabetes. DCM causes a set of structural and functional abnormalities in the heart and thus increases morbidity and mortality in diabetic patients. 1 Some molecular mechanisms, including myocyte apoptosis, cardiac fibrosis, oxidative stress, cardiac inflammation, left ventricular dysfunction, and heart failure, have been demonstrated to be involved in the pathogenesis of DCM. 2,3 It is well known that a hyperactive renin–angiotensin system (RAS) is a unique feature of the pathological progression of diabetes. 4 The activation of RAS, particularly local RAS, can further trigger a series of characteristic pathological processes that exacerbate DCM. 5
(Pro)renin receptor (PRR), which is a 350 amino acid protein with a single transmembrane domain, is a novel component of the RAS. A previous study found that PRR is widely expressed in various organs, including the heart, kidney, and brain. PRR has been reported to be involved in the pathological progression of some cardiovascular diseases, including hypertension, myocardial infarction (MI), and accelerated heart injury. 6,7 Renin and its precursor prorenin binding to PRR further enhance renin activity and fully activate the biologically inactive prorenin peptide. 8 The binding of PRR to its ligand promotes the production of Ang II and subsequently activates Ang II-dependent pathways. Moreover, PRR can also activate Ang II-independent pathways, including the mitogen-activated protein kinase (MAPK) signaling pathway, which is further involved in the pathological processes associated with the diseases. 9 Remarkably, several studies have demonstrated that Ang II-independent pathways play a significant role in the end-organ damage of cardiovascular diseases such as diabetes and hypertension. 10
A previous study showed that blocking PRR could improve blood pressure in spontaneously hypertensive rats (SHRs) and alleviate the pathological alterations to a certain extent. 11 The same function has also been demonstrated in mice with MI. 12 A recent report manifested that PRR expression is significantly increased in DCM, 13 which has shed light on the importance of PRR for the development of DCM. However, the role of PRR in DCM has not yet been clarified.
To detect the effects of PRR RNA interference (RNAi) silencing on the pathogenesis of DCM, in vivo and in vitro experiments were performed. A DCM rat model was established and then divided into four groups: a control group, DCM group, adenovirus (Ad) scrambled (SC) short hairpin RNA (shRNA) group, and Ad-PRR-shRNA group. Myocardial fibrosis levels and cardiac function in the four groups were detected in vivo. The effects of RNAi silencing on the high glucose–induced protein expression of collagen I, collagen III, and transforming growth factor beta (TGF-β) were also observed in vitro in cardiac fibroblasts. These in vivo and in vitro experiments aimed to evaluate the effects of the inhibition of PRR expression by PRR RNAi silencing on left ventricular remodeling and cardiac function in DCM and to explore the underlying mechanism.
Methods
PRR shRNA construction
Three rat PRR shRNA fragments, including PRR-shRNA-547, PRR-shRNA-691, and PRR-shRNA-274, and a SC shRNA were designed and constructed by GenePharma Company (Shanghai, P.R. China). Recombinant Ad was used to carry these shRNA fragments, and Ad-PRR-shRNA and Ad-SC-shRNA were successfully established.
H9C2 cells were used to select the most efficient PRR-shRNA fragment among these three types of PRR-shRNA. The result by Western blot demonstrated that Ad-PRR-shRNA-547 was the most efficient fragment. The selected sequence of PRR is GCTCCGTAATCGCCTGTTTCA.
Experimental animals
Sixty male 8-week-old Wistar rats weighing 200–250 g were obtained from Shandong University Animal Center and maintained in an animal room with controlled temperature (22 ± 2°C) and humidity (55 ± 5%) under a 12 h/12 h light/dark cycle.
The rats were randomly divided into the control group (n = 15) and the treatment group (n = 45). The rats in the treatment group were used to construct a DCM model by a single intraperitoneal (i.p.) injection of streptozotocin (STZ; 65 mg/kg). One week after STZ administration, rats with blood glucose levels >11.1 mmol/L and classical clinical symptoms were considered to be diabetic rats. Then, 12 weeks after STZ administration, the diabetic rats were further divided into three groups: the DCM group, the Ad-SC-shRNA group, and the Ad-PRR-shRNA group (n = 15 per group). The rats in the Ad-SC-shRNA group, Ad-PRR-shRNA group, and DCM group received an intravenous (i.v.) injection of recombinant Ad containing SC-shRNA (1 × 109 pfu), PRR-shRNA (1 × 109 pfu), or phosphate-buffered saline (PBS), respectively, via the tail vein. All rats were maintained on a normal diet after the i.v. injection. Then, 4 weeks after the Ad injection, all rats were anesthetized with an i.p. injection of 10% chloral hydrate (300 mg/kg). The heart was irrigated with normal saline from the left ventricle to all parts of body until the liver tissue became white, and the hearts were excised and washed with normal saline. The myocardium of the left ventricle was collected for pathological and biochemical analyses. During the experiments, one rat in the DCM group and one rat in the Ad-PRR-shRNA group died.
The animal protocol complied with the Guide for the Care and Use of Laboratory Animals from the National Academy of Sciences published by the U.S. National Institutes of Health (NIH publication no. 86-23, revised 1996) and with the Principles of Laboratory Animal Care formulated by the National Society for Medical Research. The protocol was approved by the Institutional Animal Care and Use Committee of Shandong University.
Echocardiography measurements
At the end of the experiment, the rats in the control group, DCM group, Ad-SC-shRNA group, and Ad-PRR-shRNA group were subjected to echocardiography using a Vevo770 imaging system (Visual Sonics, Toronto, Canada). The rats in the four groups underwent general anesthesia with 10% chloral hydrate (300 mg/kg, i.p.), and the fur in precordial region was removed from the skin. M-mode signals were collected, and the left ventricular ejection fraction (LVEF), the ratio of flow Doppler E wave to A wave amplitude (E/A), the left ventricular end diastolic diameter (LVEDD), and the left ventricular end systolic diameter (LVESD) were measured. All measurements represent the mean of five consecutive cardiac cycles.
Histology and morphometric analyses
The myocardial tissues were fixed in 4% paraformaldehyde for >24 h, embedded in paraffin, and cut into sections 4.5 μm thick. The sections were stained with hematoxylin and eosin (H&E) and Masson's trichrome staining.
Immunohistochemical staining
The 4.5-μm-thick sections obtained from paraffin-embedded myocardial tissues from the four groups were deparaffinized, and antigen retrieval was performed at a temperature >95°C, which was achieved in a microwave oven with 3 min of high heat and 13 min of low heat. Nonspecific reactions were blocked with goat serum for 15 min at 37°C. Then, the sections were incubated with primary polyclonal anti-PRR, anti-collagen I, anti-collagen III, anti-TGF-β, anti-NOX2, and anti-NOX4 antibodies (Abcam, Cambridge, MA) overnight at 4°C. All specific primary antibodies were diluted in 1 × PBS, and the dilution ratio was 1:200. The sections were incubated with a horseradish peroxidase (HRP)-labeled secondary antibody (Abcam) for 20–30 min at 37°C. The negative controls were treated without the primary antibody. The samples were viewed under a confocal FV 1000 SPD laser scanning microscope (Olympus, Tokyo, Japan).
Western blot analysis
Total proteins were extracted from myocardial tissues from the four groups, separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis, and transferred onto polyvinylidene difluoride (PVDF) membranes. After blocking nonspecific proteins with 5% skim milk, the PVDF membranes were incubated overnight at 4°C with specific antibodies diluted in Western blot primary antibody diluent at a ratio of 1:1,000. The specific primary antibodies included anti-p-ERK, anti-t-ERK (Cell Signaling Technology, Danvers MA), anti-(pro)renin receptor (Abcam), anti-NOX2 (Abcam), and anti-NOX4 (Abcam) antibodies. Glyceraldehyde 3-phosphate dehydrogenase (GAPDH; Abcam) and HRP-conjugated secondary antibodies (Abcam) were used to bind to the specific antigen–antibody complex. The immunoreactive bands were visualized by an enhanced chemiluminescence reagent. Densitometry was performed with a customized imaging analysis system Image J.
RNA extraction and real-time polymerase chain reaction
The total RNA was isolated from the myocardial tissues and cardiac fibroblasts using TRIzol Reagent (Thermo Fisher Scientific, Waltham, MA). In total, 1 mL was extracted from each sample. Completely dissociated tissues and cells were incubated with trichloromethane with one fifth of the volume of TRIzol after 5 min of placement. The samples were mixed thoroughly for full reaction and then centrifuged at 12,000 g for 15 min at 4°C. An equal volume of isopropanol was added to the supernatant, followed by centrifugation, and the RNA was purified with anhydrous ethanol. Finally, the extracted RNA was diluted in diethyl pyrocarbonate water, and the concentration was determined. cDNA was synthesized from 1 mg of total RNA by using a Prime Script 1st Strand cDNA Synthesis Kit (Takara Bio, Inc., Shiga, Japan).
Gene expression was analyzed by real-time polymerase chain reaction (PCR) using a LightCycler 480 sequence detection system (Roche, Basel, Switzerland). The samples were analyzed in triplicate. The primers for rat PRR and GAPDH were purchased from BioSune Biotechnology Company (Shanghai, P.R. China), and the experimental conditions followed the manufacturer's cycling parameters. The following primer sequences were used: PRR, forward 5′-TCTGTTCTCAACTCGCTCCC-3′, reverse 5′-TCTCCATAACGCTTCCCAAG-3′; GAPDH, forward 5′-TCTCTGCTCCTCCCTGTTCT-3′, reverse 5′-ATCCGTTCACACCGACCTTC-3′.
GAPDH was used as a reference gene. The relative quantities of the amplified cDNAs were analyzed by a LightCycler 480 (Roche), and the target values were normalized to GAPDH mRNA.
Nicotinamide adenine dinucleotide phosphate oxidase activity
Myocardial tissue homogenate was produced from the myocardial tissues in the four groups. Then, the nicotinamide adenine dinucleotide phosphate (NADPH) oxidase activity was measured in each group using an NADPH oxidase assay kit (Nanjing Jiancheng Bio-Engineering Institute, Nanjing, P.R. China). Absorption value was detected at a wavelength of 600 nm for 20 s and 80 s after the end of the reaction.
Blood glucose, blood pressure, and body weight
Blood samples from the rats in all four groups were collected from the tail vein, and blood glucose levels were measured by a one-touch glucometer (Life Scan, Milpitas, CA) at the end of the experiment.
Systolic blood pressure, diastolic blood pressure, and mean arterial pressure of the rats in the four groups were measured with a noninvasive tail-cuff system (Softron, Tokyo, Japan), and all rats were in a relaxed state during the blood pressure measurements. The evaluation of each group was repeated three times.
The body weights of the rats in the four groups were detected by an electronic scale (Seca Company, Hamburg, Germany) at the end of the experiment. All measurements were repeated three times.
Cell culture and treatments
Cardiac fibroblasts were extracted from the myocardium of 1- to 3-day-old neonatal rats. First, the whole heart was cut into the smallest pieces possible, and the myocardial matrix was removed with collagenase. Next, based on the difference in growth rates, cardiac fibroblasts were distinguished from myocardial cells. After discarding the supernatant and adding myocardial fibroblast culture medium, the cells were cultured in a humidified atmosphere of 5% CO2 and 95% air at 37°C until a typical growth pattern was observed. The culture medium consisted of endotoxin free Dulbecco's modified Eagle's medium (Thermo Fisher Scientific) supplemented with 5 mM glucose and 10% fetal bovine serum (Thermo Fisher Scientific). The cardiac fibroblasts were identified by cardiac fibroblast vimentin expression detection, and second- to third-generation cardiac fibroblasts were transplanted into six-well cell plates for further experiments.
To determine the influence of high glucose on PRR expression in cardiac fibroblasts, the cardiac fibroblasts were divided into three groups: the normal glucose group (5.5 mM glucose), the high glucose group (25 mM glucose), and the mannitol high permeability control group (19.5 mM mannitol +5.5 mM glucose). PRR expression levels were detected by immunofluorescence staining. PRR expression in the cardiac fibroblasts following high glucose stimulation was observed in a time-dependent manner at 0, 2, 6, 12, and 24 h.
Ad-shRNA transfection and treatments in vitro
The cardiac fibroblast (1 × 108) was transplanted onto six-well plates. The cells were grown to 80–90% confluence for further treatment. The cardiac fibroblasts were cultivated in culture medium without serum for >12 h first, and then Ad-PRR-shRNA and Ad-scrambled-shRNA were transfected into the Ad-PRR-shRNA group and Ad-SC-shRNA group, respectively. The quantity of Ad-shRNA to be transfected in each well was determined by the cell number to obtain a multiplicity of infection (MOI) value up to 150:1. This MOI value was determined by previous experiments. The target sequence of Ad-PRR-shRNA was the same as that used in the rat experiments. After 12 h of incubation with the transfection reagent, the cells were switched to normal medium for another 12 h for recovery and prepared for further experiments.
The cardiac fibroblasts from the high glucose group were divided into four groups: the high glucose group, the Ad-SC-shRNA group, the Ad-PRR-shRNA group, and the losartan group. Both the Ad-SC-shRNA group and the Ad-PRR-shRNA group were transfected with Ad-shRNA following the methods described above. Then, high glucose stimulation was applied for another 12 h. In the losartan group, the cardiac fibroblasts were first pretreated with losartan (20 μM; Abcam) in normal glucose culture medium for 1 h and then treated with 25 mM glucose and drugs for another 12 h.
Immunofluorescence staining
The cardiac fibroblasts were seeded in 24-well chamber slides and treated with normal glucose medium, mannitol high permeability medium, and high glucose medium for 24 h. PRR protein expression in the control group, high glucose group, and high permeability group was detected by immunofluorescence staining. First, the cardiac fibroblasts in each group were fixed with 4% paraformaldehyde. Then, the fibroblasts were blocked with 10% bovine serum albumin and stained with an anti-PRR antibody (dilution ratio 1:250; Abcam) overnight at 4°C. After applying fluorescein isothiocyanate–conjugated immunoglobulin G, the cell nuclei were stained with 6-diamidino-2-phenylindole, and the samples were sealed with anti-fluorescence quenching agent (Abcam). The immunofluorescence intensity was visualized under a fluorescence microscope (Leica, Wetzlar, Germany).
Enzyme-linked immunosorbent assay
Cardiac fibroblasts stimulated with high glucose were divided into four groups: the high glucose group, the Ad-SC-shRNA group, the Ad-PRR-shRNA group, and the losartan group. The cell supernatant from each group was collected for enzyme-linked immunosorbent assay (ELISA) to detect the expression of type I collagen, type III collagen, and TGF-β in the different groups. All measurements were performed according to the manufacturer's instructions (Abcam). The experiment was repeated at least three times per group. The optical density (OD) value of the sample was determined at a wavelength of 450 nm by an enzyme standard instrument. The relative factor concentrations were calculated according to standard sample OD values and concentrations.
Statistical analysis
Statistical analyses were performed using IBM SPSS Statistics for Windows v19.0 (IBM Corp., Armonk, NY). All data are expressed as the mean ± standard deviation of at least three representative independent experiments. The differences were analyzed by one-way analysis of variance. A p-value of <0.05 was considered significant.
Results
PRR RNAi silencing decreased PRR expression in vivo
At the end of the experiment, PRR expression in the control group, DCM group, Ad-SC-shRNA group, and Ad-PRR-shRNA group was evaluated by PCR and Western blot (Fig. 1). The PCR results showed that PRR mRNA expression in the DCM group and Ad-SC-shRNA group was significantly upregulated compared to that in the control group. However, this upregulation was significantly alleviated in the Ad-PRR-shRNA group (Fig. 1A). The PRR protein expression in the DCM group and Ad-SC-shRNA group detected by Western blot was also significantly increased compared to that in the control group. However, the Ad-PRR-shRNA group displayed a significant decrease compared to the DCM group and the Ad-SC-shRNA group (Fig. 1B and C).

(Pro)renin receptor (PRR) expression in four groups of rats.
Immunohistochemistry was also used to evaluate PRR protein expression in the four groups. The results showed that PRR expression in the Ad-PRR-shRNA group was statistically lower than that in the DCM group and the Ad-SC-shRNA group (Fig. 1D and E).
H&E staining was measured in the four groups. The representative H&E staining of myocardium tissues in the four groups is shown in Fig. 1F. In addition, the basic characteristics of animals in the four different groups were also evaluated and are shown in Table 1.
Basic characteristics of animals in the four different groups
p < 0.01 vs. control group; # p < 0.05 vs. DCM group, Ad-SC-shRNA group.
DCM, diabetic cardiomyopathy; Ad, adenovirus; SC, scrambled; shRNA, short hairpin RNA; PRR, (pro)renin receptor.
PRR RNAi silencing improved cardiac function in DCM
At the end of week 16, the rat cardiac function parameters in the control group, DCM group, Ad-SC-shRNA group, and Ad-PRR-shRNA group were detected by echocardiography (Fig. 2A and Table 2).

Cardiac function data in four groups of rats 4 weeks after PRR shRNA interference.
Cardiac function data in the four different groups
p < 0.01 vs. control group; # p < 0.05 vs. DCM group, Ad-SC-shRNA group.
LVEDD, left ventricular end diastolic diameter; LVESD, left ventricular end systolic diameter; E/A, ratio of flow Doppler E wave to A wave amplitude; EF, ejection fraction.
The results showed that the LVEF, which is an important indicator of systolic function in the left ventricle, was much lower in the DCM group and the Ad-SC-shRNA group than in the control group. In contrast, the LVEF in the Ad-PRR-shRNA group was statistically higher than that in the DCM group and the Ad-SC-shRNA group (Fig. 2B and Table 2). Regarding the heart size–associated parameters LVEDD and LVESD, the results showed that LVEDD was significantly increased in the DCM group and the Ad-SC-shRNA group compared to that in the control group. However, the administration of PRR RNAi silencing in the Ad-PRR-shRNA group resulted in a significant decrease in the LVEDD compared to that in the DCM group and the Ad-SC-shRNA group (Fig. 2C and Table 2). Furthermore, the LVESD was lower in the Ad-PRR-shRNA group than that in the DCM group and the Ad-SC-shRNA group (Fig. 2D and Table 2). Moreover, the diastolic function was also evaluated, and the results showed that the E/A ratio was statistically higher in the Ad-PRR-shRNA group than that in the DCM group and the Ad-SC-shRNA group (Fig. 2E and Table 2).
PRR RNAi silencing attenuated myocardial fibrosis in DCM
Masson's trichrome staining results showed that the Ad-PRR-shRNA group treated with PRR RNAi silencing had much lower collagen expression than the DCM group and the Ad-SC-shRNA group (Fig. 3A and B).
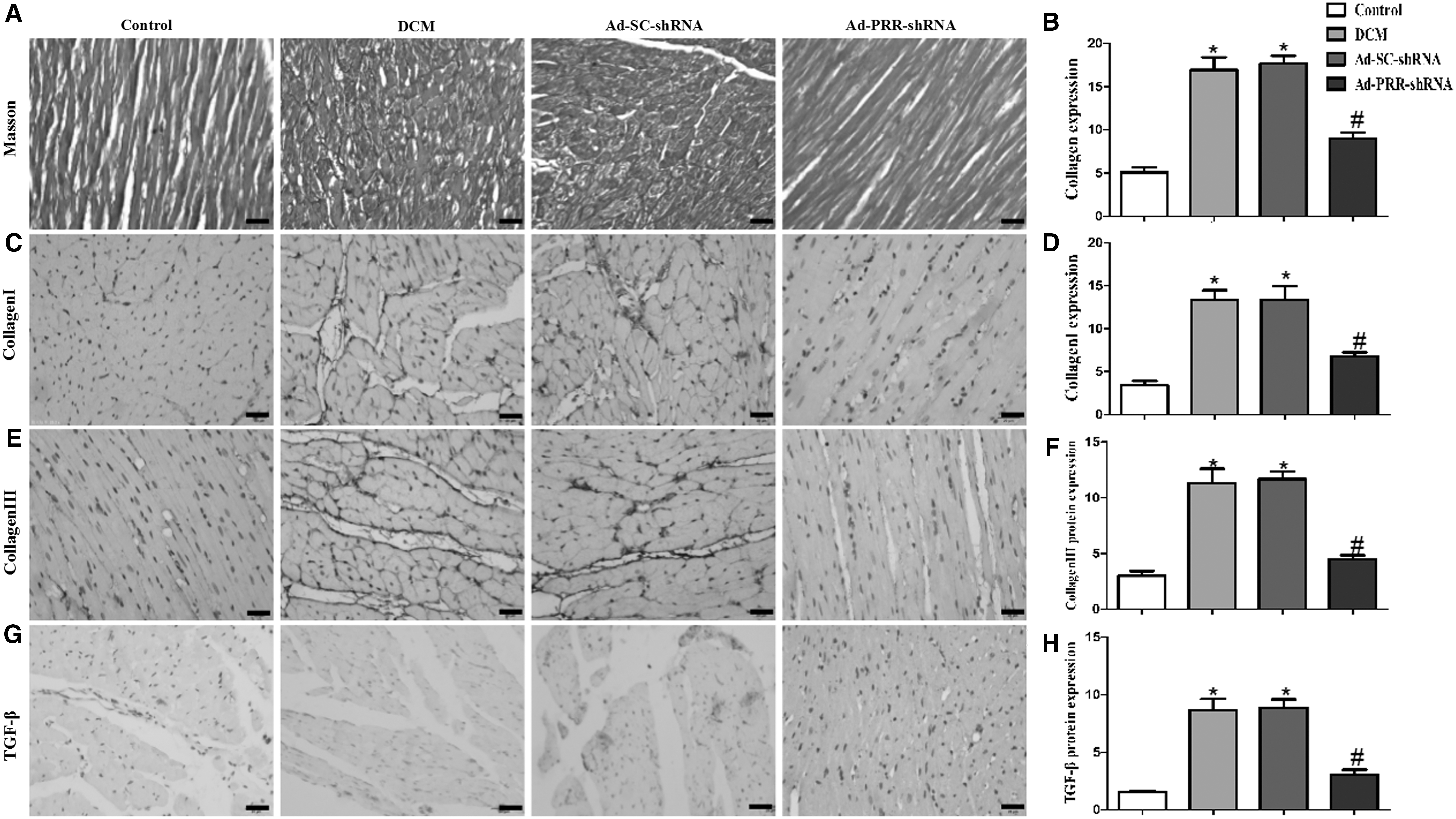
Masson's trichrome staining of the myocardium and collagen protein expression in four groups of rats.
The protein expression of type I and type III collagen in the four groups was detected by immunohistochemical staining. The results showed that the collagen I protein expression level in the Ad-PRR-shRNA group was significantly lower than that in the DCM group and the Ad-SC-shRNA group (Fig. 3C and D). The collagen III protein expression level in the Ad-PRR-shRNA group was also lower than that in the DCM group and the Ad-SC-shRNA group (Fig. 3E and F). Then, the results by immunohistochemical staining also showed that the TGF-β protein expression level in the DCM group and the Ad-SC-shRNA group was higher than that in the control group. In contrast, PRR RNAi silencing in the Ad-PRR-shRNA group decreased its expression (Fig. 3G and H).
PRR RNAi silencing attenuated the myocardial oxidative stress response
The NOX2 and NOX4 protein expression measured by immunohistochemical staining was upregulated in the DCM group and the Ad-SC-shRNA group compared to that in the control group. However, these high protein expression levels were significantly alleviated in the Ad-PRR-shRNA group (Fig. 4A and C). The Western blotting results displayed the same trend, showing that NOX2 and NOX4 protein expression in the Ad-PRR-shRNA group was lower than that in the DCM group and the Ad-SC-shRNA group (Fig. 4E and H).

Oxidative stress response level, total ERK, and phosphorylated ERK protein expression in four groups of rats.
The analysis of NADPH oxidase activity showed that the myocardial NADPH oxidase activity in the DCM group and the Ad-SC-shRNA group was significantly higher than that in the control group. Moreover, the NADPH oxidase activity in the Ad-PRR-shRNA group was lower than that in the DCM group and the Ad-SC-shRNA group (Fig. 4I).
PRR RNAi silencing attenuated myocardial ERK signaling activation
The level of ERK phosphorylation as detected by Western blot was statistically higher in the DCM group and the Ad-SC-shRNA group than that in the control group. However, this upregulation was mitigated by the administration of PRR RNAi silencing in the Ad-PRR-shRNA group (Fig. 4J).
PRR RNAi silencing significantly alleviated PRR expression in vitro
First, the changes in PRR expression following high glucose stimulation were observed in three groups: the high glucose group, the normal glucose group, and the mannitol high permeability control group. Expression was evaluated by immunofluorescence staining. The results showed that the PRR expression in the high glucose group was much higher than that in the normal glucose group and the mannitol high permeability control group (Fig. 5A and B). The PCR and Western blot results also showed that high glucose stimulation upregulated PRR expression in a time-dependent manner. In addition, 12 h after high glucose stimulation, PRR expression reached a peak (Fig. 5C and D).

PRR expression in high glucose stimulated cardiac fibroblasts.
Second, the effects of PRR RNAi silencing on PRR expression were observed. As revealed by Western blot analysis, the results showed that using PRR RNAi silencing in the Ad-PRR-shRNA group significantly decreased PRR expression compared to that in the high glucose group and the Ad-SC-shRNA group in vitro (Fig. 6A).
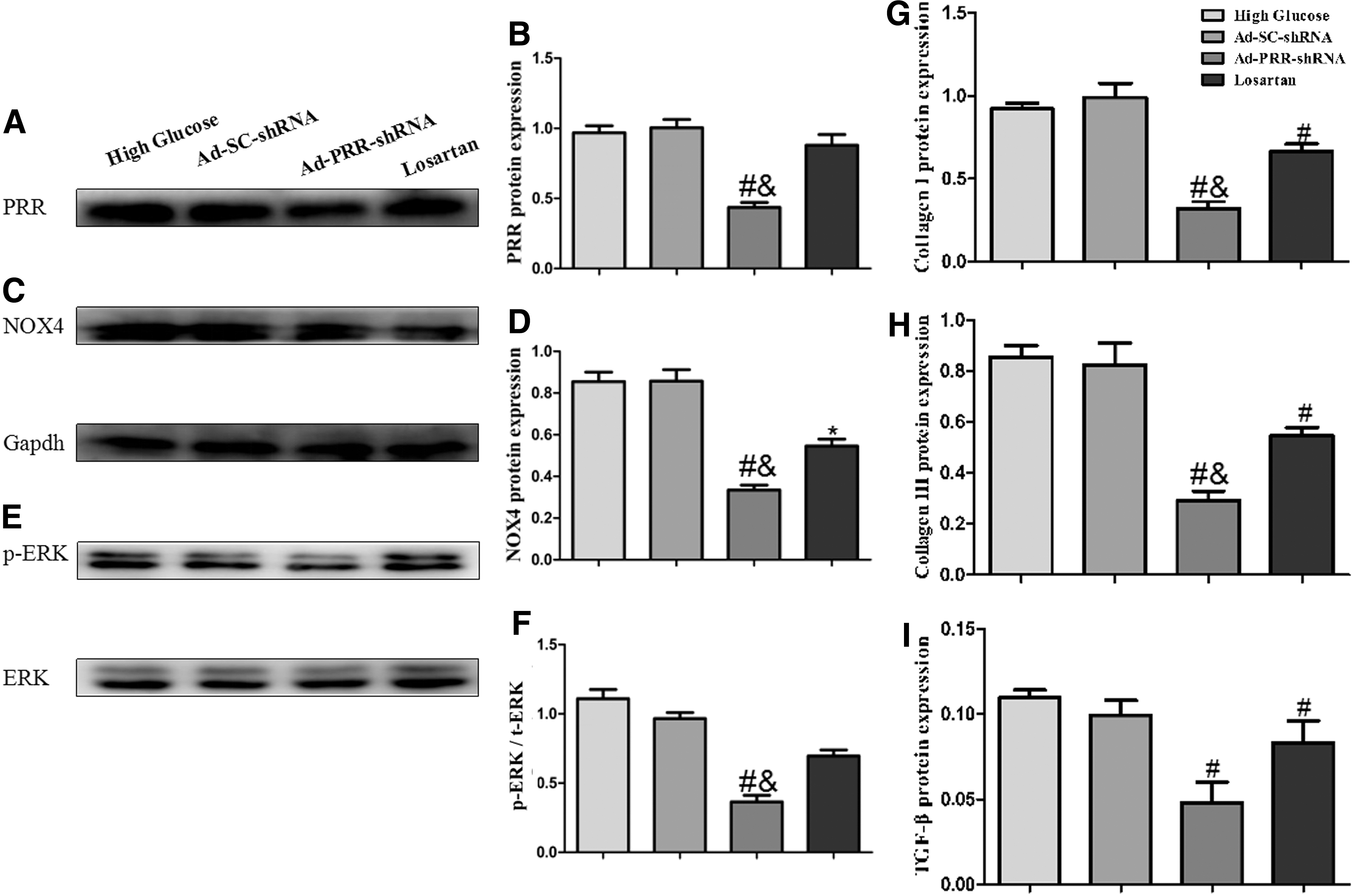
PRR, NOX4, total ERK, and phosphorylated ERK protein expression and fibrosis levels in the four groups in vitro.
PRR RNAi silencing significantly alleviated high glucose–induced fibrosis in vitro
The collagen I and collagen III protein expression levels in the high glucose group, Ad-SC-shRNA group, Ad-PRR-shRNA group, and losartan group were measured by ELISA. The results showed that collagen I and collagen III were significantly upregulated in the high glucose group and the Ad-SC-shRNA group, but these expression levels were statistically lower in the Ad-PRR-shRNA group and the losartan group. Moreover, the expression of collagen I and collagen III in the Ad-PRR-shRNA group was lower than that in the losartan group (Fig. 6G and H). ELISA analysis also showed that the expression of TGF-β was lower in the Ad-PRR-shRNA group and the losartan group than that in high glucose group and the Ad-SC-shRNA group (Fig. 6I). In addition, TGF-β protein expression was not statistically different between the Ad-PRR-shRNA group and the losartan group (Fig. 6I).
PRR RNAi silencing significantly alleviated high glucose–induced oxidative stress and ERK activation in vitro
As revealed by Western blot analysis, high glucose–induced expression of NOX4 was significantly alleviated in the Ad-PRR-shRNA group and the losartan group. However, the Ad-PRR-shRNA group exhibited a more significant reduction than the losartan group (Fig. 6C).
The results by Western blot also showed that ERK phosphorylation levels were decreased in the Ad-PRR-shRNA group compared to those in the high glucose group and the Ad-SC-shRNA group. Furthermore, the ERK phosphorylation levels in the Ad-PRR-shRNA group were lower than that in the losartan group (Fig. 6E).
Discussion
This study found that the inhibition of PRR expression by PRR RNAi silencing in vivo alleviated myocardial fibrosis and left ventricular remodeling and improved cardiac function. In vitro, PRR shRNA attenuated collagen and TGF-β expression in cardiac fibroblasts following high glucose stimulation. In addition, PRR RNAi silencing alleviated high glucose–induced oxidative stress upregulation and ERK signaling pathway activation. The results indicate that inhibiting PRR expression by PRR RNAi silencing could be a new efficient treatment for DCM.
DCM has a high morbidity and mortality and is a major cardiac complication in patients with diabetes. Systematic and local RAS activation is an initial pathological process in DCM that triggers a series of pathological changes, such as myocardial fibrosis, LV remodeling, and heart failure. PRR is a novel component of RAS that combined with renin or prorenin is involved in several pathological processes associated with cardiovascular diseases. 14 Fukushima et al. reported that PRR plays a crucial role in MI. 14 Hayakawa et al. suggested that high salt intake induced high PRR expression in hypertension and could exacerbate cardiac fibrosis and result in early heart injury. 15 Some authors have reported that overexpression of PRR in the mouse atrium induced significant dilation and fibrosis and was associated with a high incidence of sudden death. 16 However, blockade of PRR expression could be beneficial for some cardiovascular diseases. 17 These studies suggest that PRR induction could be a common pathological finding in heart injury and may be a novel therapeutic target for cardiovascular diseases.
Recent research reported that PRR expression is significantly increased in DCM. 13 The present study found that PRR expression in the myocardium was notably increased in DCM rats at both the mRNA and protein levels. This finding is consistent with previous reports. In contrast, after PRR RNAi silencing by Ad-PRR-shRNA, the expression of PRR at both the mRNA and protein levels was significantly decreased in the Ad-PRR-shRNA group compared to that in the high glucose group. In vitro, the PRR RNAi silencing also decreased PRR expression in the high glucose stimulated cardiac fibroblasts.
The most important pathological feature of DCM is the accumulation of extracellular matrix (ECM) proteins, especially collagen. 18 Upregulated PRR expression in the diabetic kidney has been shown to exacerbate renal fibrosis and result in end-stage renal disease. 19 In contrast, a recent study found that the administration of a PRR inhibitor diminished cardiac fibrosis in SHRs and reversed diabetic nephropathy. 20 Ellmers et al. also demonstrated that blocking PRR after MI in mice alleviated cardiac fibrosis and improved cardiac dysfunction. 12 However, few studies have investigated the involvement of PRR in the pathogenesis of DCM.
In the present study, blocking PRR expression by PRR RNAi silencing exerted the same protective function in DCM. PRR RNAi silencing ameliorated the accumulation of ECM proteins, including type I collagen and type III collagen, accumulated in DCM. An important factor regulating collagen production in DCM is TGF-β, and the experimental results showed that PRR RNAi silencing can downregulate its high expression under the high glucose condition. Gonzalez et al. found that PRR knockdown in collecting duct cells prevented the upregulation of profibrotic proteins. 21 The present study also showed that PRR RNAi silencing decreased the high glucose–induced upregulation of collagen I and collagen III in cardiac fibroblasts, which is consistent with previous observations.
Recent studies have shown that PRR binding to its ligands activates not only the Ang II-dependent pathway but also the Ang II-independent pathway, which is involved in pathological progression of cardiovascular disease. The overexpression of PRR in losartan-treated animals has been reported to result in myocardium fibrosis in vivo and Ang II-independent pathway activation in vitro. 22 The present study found that PRR RNAi silencing has a more effective function in inhibiting cardiac fibrosis than losartan, supporting previous studies. These results indicate that PRR RNAi silencing might decrease collagen production and further alleviate DCM-induced cardiac fibrosis.
The echocardiography results showed that PRR RNAi silencing improved systolic and diastolic function as shown by increased LVEF and decreased LVEDD and LVESD values, indicating that PRR RNAi silencing enhanced systolic function. Similarly, rats treated with PRR RNAi silencing had a higher E/A ratio than DCM rats, indicating that PRR RNAi silencing also enhanced diastolic function.
The following putative mechanisms may underlie the benefits of PRR RNAi silencing: inhibition of the oxidative stress response and ERK signaling pathway blockade. Previous studies have reported that diabetes-induced oxidative stress response activation could further accelerate cardiac fibrosis and cardiac function injury. 23 Hua Peng et al. demonstrated that PRR could upregulate oxidative stress by the Ang II-dependent and -independent pathway in neuronal cells to mediate blood pressure. 9,23 In the present in vivo study, PRR RNAi silencing downregulated NOX2 and NOX4 protein expression in the myocardium of diabetic rats, and NADPH oxidase activity was also mitigated by PRR blockade. This inhibitory function was also observed in the experiment with cardiac fibroblasts, and the results showed that PRR RNAi silencing attenuated the high glucose–induced oxidative stress response upregulation and was much more effective than losartan.
PRR has been reported to induce the Ang II-independent MAPK pathway, which is involved in the pathological progression of cardiovascular diseases. Previous studies have shown that PRR blockade abolishes the increased MAPK activation in diabetic kidneys, further relieving pathological progression. 24 The present experiment showed that PRR RNAi silencing decreased the ERK phosphorylation level, which was significantly upregulated in DCM. On the basis of the present results and the results of previous studies, it can reasonably be believed that PRR RNAi silencing attenuates cardiac fibrosis and cardiac dysfunction in DCM by inhibiting the ERK signaling pathway.
In conclusion, diabetes induces high PRR expression in the myocardium and exacerbates fibrosis progression, resulting in cardiac dysfunction in DCM. PRR RNAi silencing exerts significant therapeutic effects on attenuating myocardial fibrosis and improving cardiac function in vitro and in vivo. The possible mechanisms may involve the inhibitory effects on ERK signaling pathway activation and oxidative stress upregulation, which are induced by PRR-mediated Ang II-dependent and -independent MAPK pathway activation. The inhibition of PRR expression could be a new therapeutic intervention for DCM in the future.
Footnotes
Author Disclosure
No competing financial interests exist.