
Abstract
Poorly differentiated thyroid carcinoma cells tend to be more aggressive and show enhanced glucose uptake which could be exploited for anti-cancer strategy. Previously, we identified hexokinase 2 (HK2) as a direct target of miR-143. In our current study, the effects of miR-143 on glucose metabolism and tumor biological behavior were investigated in FTC-133 cells which is a poorly differentiated thyroid carcinoma (PDTC). Additionally, tumor-bearing mice xenografts of PDTC were constructed, with encapsulated miR-143 agomir being administered intravenously. 18F-FDG microPET-CT scanning was used for the evaluation of therapeutic efficacy. The tumor-restrained effect of miR-143 was demonstrated in PDTC. Furthermore, microPET/CT imaging exhibited a reduction of 18F-FDG uptake in tumors, corresponding to the downregulated expression of HK2 in tissues. In summary, our results suggest that miR-143 can be an alternative treatment for PDTC and the specific assessment of therapeutic response to miR-143 can be achieved by 18F-FDG microPET/CT in advanced thyroid carcinoma xenografts.
Introduction
As the most common endocrine malignancy, the incidence of thyroid cancer continues to rise worldwide over the past decade. Generally, patients with differentiated thyroid carcinoma (DTC), which accounts for approximately 90% of cases, have favorable prognosis via standard therapeutic approach including surgical operation, radioactive iodine (RAI) treatment, and thyrotropin suppression. 1 Nevertheless, only a minority of cases show poorly differentiated thyroid carcinomas (PDTC), which predominantly originates from the variants of DTC, with impaired or lack of avidity for RAI. 2,3 These advanced cases are frequently more aggressive with worse prognosis, with a mean survival of 3.2 years following diagnosis. 3 Although tyrosine kinase inhibitors such as Sorafenib and Lenvatinib have been shown to be effective in improving the progression-free survival of RAI-refractory thyroid cancer patients, there is little evidence revealing the prolongation of the overall survival (OS). 4,5 Besides, the deleterious side effects cannot be ignored. Consequently, new therapeutic alternatives are required.
One of the prominent features of cancer is a high rate of aerobic glycolysis, which helps to distinguish cancer cells from normal cells and is used extensively for imaging tumors in vivo using 18 F-fluorodeoxyglucose ( 18 F-FDG) positron emission tomography/computed tomography (PET/CT). 6 Many studies have shown that 18 F-FDG uptake is inversely related to RAI uptake and tumor differentiation in thyroid cancer, namely iodine/FDG “flip-flop phenomenon.” 7 –9 Consequently, PDTC is often characterized by damaged RAI uptake and increased cellular glucose metabolism, suggesting the potentiality to target glycolysis in PDTC treatment. The first rate-limiting reaction in the glycolytic pathway is considered to be the most significant step in glucose metabolism. This step is catalyzed by hexokinases; among these is hexokinase 2 (HK2), which is aberrantly expressed at high levels in many cancers and is being suggested as a glycolytic target in cancer therapy. 10,11
MicroRNAs (miRNAs) are small RNAs that function as regulators of gene expression by negatively regulating protein translation at post-transcriptional level. 12,13 miR-143 plays a tumor-suppressive role and represses multiple target genes of signaling pathways involved in proliferation, metastasis, and apoptosis. 14,15 miR-143 is abnormally expressed in a variety of cancers including thyroid cancer. The downregulation of miR-143-3p was found to be related to the aggressive behavior in cases of DTC, 16 suggesting the possibility of miR-143 as a therapeutic target in PDTC.
Previously, we identified the glycolytic gene HK2 as a direct target of miR-143. 17 However, whether miR-143-HK2 axis is related to the elevated glycolysis of PDTC is unknown, as is the potential therapeutic role of miR-143 in PDTC. In the present study, we explored the therapeutic potentiality of miR-143 in PDTC xenografts and demonstrated that 18 F-FDG microPET/CT could be exploited for visual evaluation during miR-143 treatment.
Materials and Methods
Cell culture
The FTC-133 human poorly differentiated follicular thyroid carcinoma cell line 18,19 and the 8505C human anaplastic thyroid carcinoma (ATC) cell line were obtained from the European Collection of Animal Cell Cultures (Salisbury, UK). The C643 human ATC cell line was obtained from Typical Culture Preservation Commission Cell Bank, Chinese Academy of Sciences (Shanghai, China). FTC-133 cells were cultured in Dulbecco's Modified Eagle's Medium/Ham's F12 (DMEM/F12) medium and cells of 8505C and C643 were maintained in Roswell Park Memorial Institute (RPMI) 1640 medium, supplemented with 10% fetal bovine serum (FBS), 100U/mL penicillin and 100μg/mL streptomycin at 37°C, 5% CO2.
RNA oligonucleotide and cell transfection
miR-143 mimic and scrambled negative control RNA (Ctrl RNA) (RiboBio, Guangzhou, China) were at the molarity of 50 nM when the cells were transfected with Lipofectamine 2000 (Invitrogen, Carlsbad, CA) in accordance with the manufacturer's instructions.
RNA isolation and qRT-PCR assays
The RNeasy Mini Kit (Qiagen, Hilden, Germany) was used for extracting the total RNA from cells and tissues. The PrimeScript RT reagent (Takara, Shiga, Japan) and SYBR Premix Ex Taq II (Takara) were used for quantitative real-time PCR (qRT-PCR) assays, separately with β-actin and U6 small nuclear RNA as endogenous references for HK2 and miR-143 quantification. The main sequences used in the study are listed below:
HK2 forward primer, 5′-AAGGCTTCAAGGCATCTG-3′;
HK2 reverse primer, 5′-CCACAGGTCATCATAGTTCC-3′; miR-143 forward primer, 5′-ACACTCCAGCTGGGTGAGATGAAGCACTGT-3′; and miR-143 reverse primer, 5′-CTCAACTGGTGTCGTGGAGTCGG-3′.
The relative levels of HK2 mRNA and miR-143 were analyzed with the Ct method. The supplemental sequences of primers used in qRT-PCR are listed in Supplementary Table S1.
Cell proliferation, apoptosis, and transwell migration assays
FTC-133, 8505C, and C643 cells were prepared for further assays after being transfected with RNA oligonucleotide (miR-143 mimic/Ctrl RNA). In brief, equal numbers of cells were seeded in 96-well plates. 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) assays were performed to evaluate cell viability, and Annexin V-FITC Apoptosis Detection Kit (Sigma, St. Louis, MO) was used with flow cytometry analysis to assess cell apoptosis. Experiments were carried out in triplicate. Cell migration assays were performed in 24-well transwell plates with PET membrane filters (BD Biosciences, Franklin Lakes, NJ) separating the upper and lower chambers. The bottom contained DMEM/F12 or RPMI-1640 with 10% FBS; the upper contained serum-free DMEM/F12 or RPMI-1640 with PDTC and ATC cells being plated in for further migration assay.
Measurement of glucose consumption, lactate production, glucose-6-phosphate production, extracellular acidification rate, and in vitro 18 F-FDG uptake
Glucose consumption and lactate production were measured using a glucose assay kit (Sigma) and a lactate assay kit (BioVision, Milpitas, CA) as previously described. 17 The rates of glucose consumption and lactate production were normalized to cell number. Glucose-6-phosphate (G6P) was analyzed using a glucose-6-phosphate assay kit (Sigma) according to the recommended protocol. The extracellular acidification rate (ECAR) was determined on an XF24 Extracellular Flux analyzer (Seahorse Bioscience) according to the manufacturer's instructions. The data was analyzed by XFe-24 software and normalized to the protein content. In 18 F-FDG uptake assays, the FTC-133, 8505C, and C643 cells were pretreated in the glucose-free medium for glucose starvation. Six hours later, cells were incubated with 18 F-FDG in the glucose-free medium at an initial concentration of 37 kBq/mL. After incubation of FTC-133 cells for 30, 60, 90, and 120 min while incubation of 8505C and C643 cells for 120 min, the cells were rinsed with cold phosphate-buffered saline (PBS) and harvested by trypsinization. 18 F-FDG radioactivity was measured by a gamma counter and exhibited as percent 18 F-FDG uptake.
Xenograft model and in vivo therapeutic experiments
Animal experiments were performed under the U.S. National Institutes for Health Guide for the Care and Use of Laboratory Animals (1996) and in compliance with the institutional ethical guidelines for animal experimentation. FTC-133 cells (2 × 106) were subcutaneously injected into the right flank of 6-week-old female BALB/c-nu mice to construct PDTC xenografts. The mice were randomly divided into three groups (n = 3). When the tumor volumes reached around 100–130 mm3, Cy3-labeled miR-143 agomir, and Ctrl RNA agomir (RiboBio, Guangzhou, China) were encapsulated by MaxSuppressor™ In Vivo RNA-LANCEr II (BIOO Scientific, Austin, TX), a neutral lipid emulsion composed of neutral lipid, nonionic detergent, oil, and small molecules, followed by the ultrasonic disruption. 20,21 Subsequently, the miRNA agomirs were administered to PDTC xenografts by tail vein injection at the dose of 1.5 mg/kg once every 3 days for six treatments. Tumor volumes were monitored every 3 days throughout the treatments.
MicroPET/CT imaging
MicroPET/CT scanning and image analysis were separately conducted before the first treatment and after six treatments, utilizing an Inveon Acquisition Workplace (Siemens, Munich, Germany) and an Inveon MM Platform (Siemens). PDTC tumor-bearing mice were fasted for at least 6 h before the injection of 18 F-FDG. Firstly, the mice were anaesthetized with 2% isoflurane in O2 gas, followed by an injection of 18 F-FDG (each tumor-bearing mouse was given 3.7 − 7.4 Mbq 18 F-FDG intravenously in 100 μL saline). Forty minutes later, a 5-min CT scan and a 10-min PET scan were conducted successively while the mice were kept in the state of continuous anaesthesia with 1.5% isoflurane in O2 gas. For semiquantitative analysis, the three-dimensional regions of interest (ROIs) were strictly delineated over the borders of the tumors under the guidance of CT images. The maximum standardized uptake value (SUVmax) and the mean standardized uptake value (SUVmean) were acquired automatically once the ROIs were delineated.
Toxicity assessment
Body weights of mice were monitored every 3 days throughout the study. At the end of the treatments, blood samples were collected from the mice to evaluate the hematological and biochemical markers including white blood cells counts, hemoglobin, platelet counts, alanine aminotransferase, aspartate aminotransferase, and urea. After the mice being sacrificed, tumor and organ tissues (heart, brain, lung, liver, kidney, stomach, and muscle) were removed for frozen sections with nucleus counterstained by DAPI. Cy3 signals were detected using fluorescence microscope to assess the distribution of miRNA.
Western blotting and immunohistochemistry
The assays were performed using standard experimental procedures. Antibodies for HK2, GAPDH, Ki-67, and Caspase-3 were from Cell Signaling Technology (Danvers, MA). Horseradish peroxidase–conjugated goat anti-rabbit immunoglobulin G and β-actin antibodies were from Sigma. Paraffin-embedded tumor tissues from xenografts were sectioned to a thickness of 5 mm. Sections were dewaxed and rehydrated in concentration gradient ethanol and then distilled water. Routine hematoxylin and eosin staining was performed. After being incubated with the primary antibody (anti-HK2, anti-Ki-67, and anti-Caspase-3) overnight at 4°C, the sections were washed with PBS and then incubated with horseradish peroxidase–conjugated secondary antibodies. Subsequently, peroxidase substrate DAB kit was used for visualization in accordance with the manufacturer's instructions (Thermo Fisher Scientific, Waltham, MA).
Statistical analysis
The results are expressed as mean ± standard deviation. The relationships between treatment groups and their paired controls were analyzed using Student's t-test. A p-value <0.05 indicated that the difference was statistically significant.
Results
miR-143 inhibits glycolysis of PDTC by downregulating HK2 expression
We first examined whether the HK2 gene could be regulated by miR-143 in FTC-133 cells, which, being refractory to the uptake of RAI, falls into the category of poorly differentiated follicular thyroid carcinoma. As a result, HK2 mRNA and protein expression were found to be markedly suppressed when FTC-133 cells were transfected with miR-143 mimic (Fig. 1A–C). Moreover, miR-143 overexpression significantly attenuated the rates of glucose consumption and lactate production, as well as the levels of G6P in FTC-133 cells (Fig. 1D–F). Using the Seahorse XF24 Extracellular Flux analyzer, we noticed that ECAR, which reflects the overall flux of glycolysis, was remarkably suppressed by miR-143 (Fig. 1H). Additionally, overexpression of miR-143 resulted in marked reduction of in vitro 18 F-FDG uptake at 120 min (Fig. 1G), suggesting that miR-143 restrained glycolysis of PDTC by negatively regulating HK2 expression.
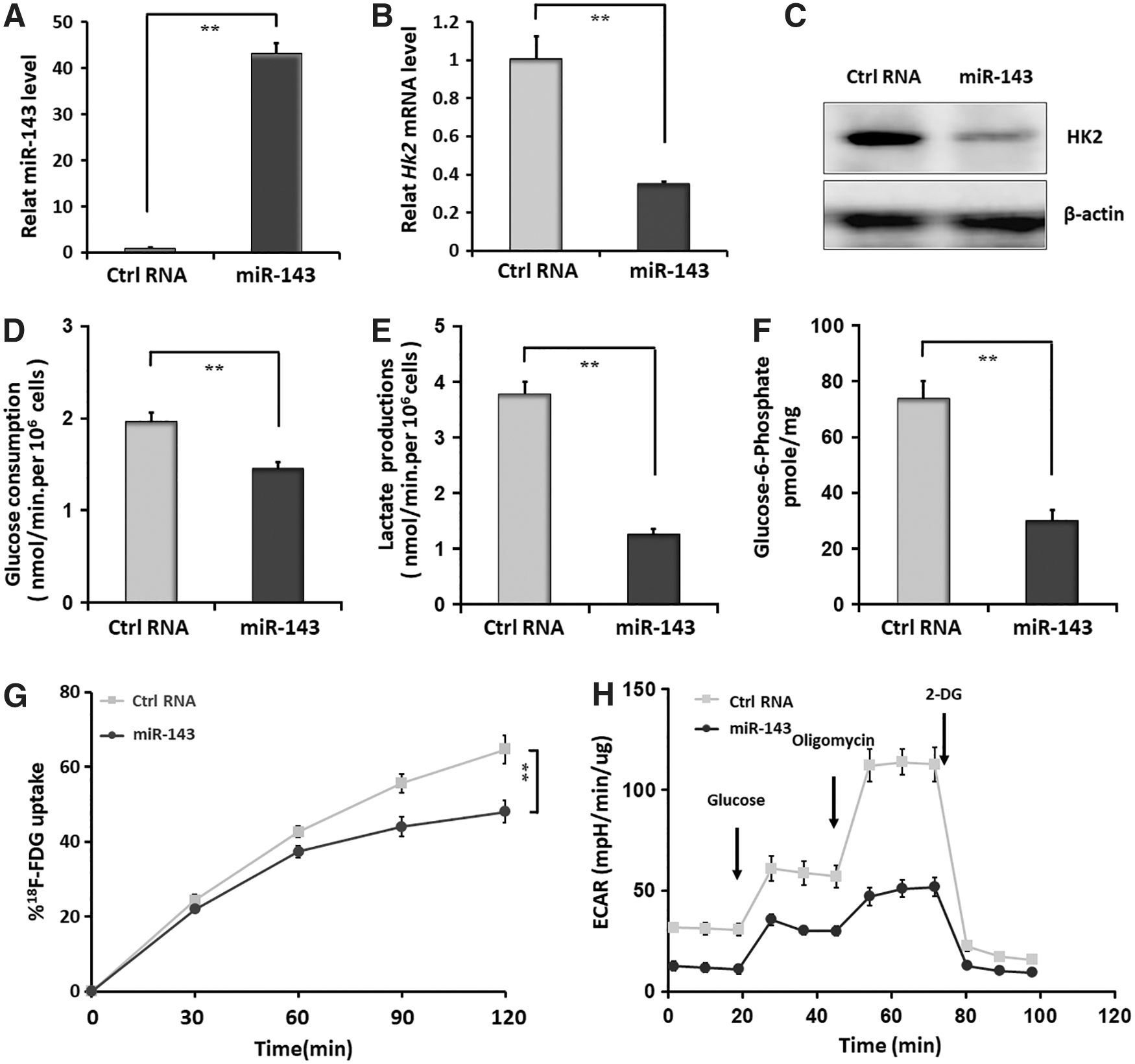
MicroRNA (miR)-143 represses glycolysis of poorly differentiated thyroid carcinomas (PDTC) by downregulating hexokinase 2 (HK2) expression.
miR-143 suppresses PDTC cells proliferation, migration, and induces apoptosis
To verify the tumor-restrained effect of miR-143 on PDTC cells, we first performed MTT assays, followed by Transwell migration and flow cytometry assays to assess the in vitro effects of miR-143 on PDTC collectively. The results showed that transfection of miR-143 mimic significantly suppressed the proliferative and migratory abilities of FTC-133 cells compared with the negative control group (Fig. 2A and B). In addition, miR-143 further exhibited a potent pro-apoptotic effect on FTC-133 cells (Fig. 2C). These results support the tumor-inhibitory effect of miR-143 on PDTC cells.
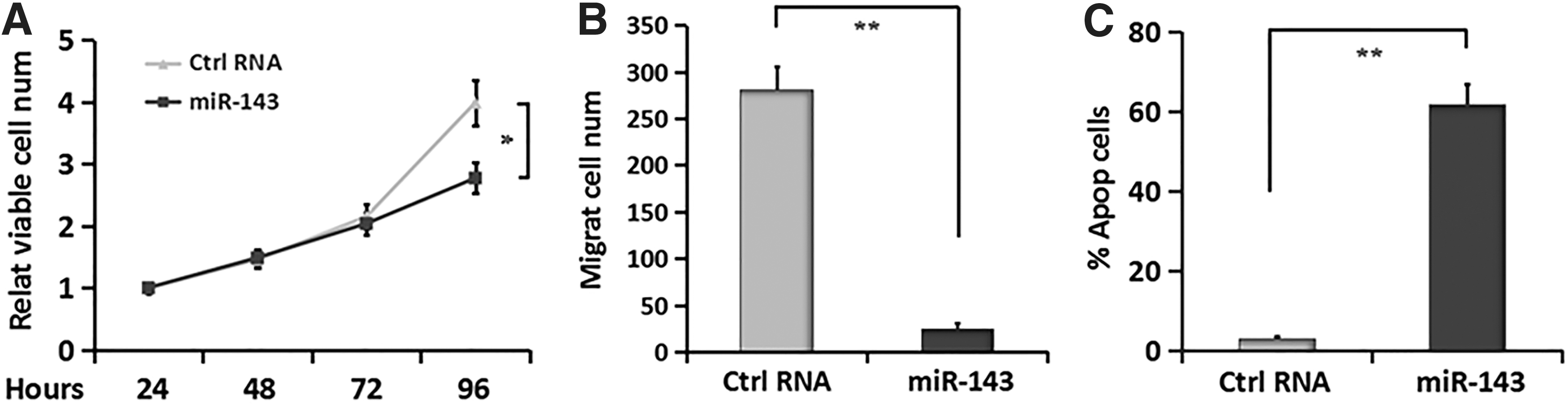
miR-143 inhibits proliferative and migratory abilities of PDTC cells and induces apoptosis.
miR-143 inhibits glycolysis of ATC by downregulating HK2 expression, suppresses ATC cell proliferation and migration, and induces apoptosis
To further confirm the effects of miR-143 on glycolysis and tumor biological behavior in advanced thyroid carcinoma, we used the 8505C and C643 ATC cells which are highly aggressive, with lack of avidity for RAI resembling PDTC for further investigation. Firstly, we studied whether HK2 gene could be regulated by miR-143 in 8505C and C643 cells. The results showed that expression of both HK2 mRNA and protein were significantly downregulated when the ATC cells were introduced with miR-143 mimic (Fig. 3A and B). Furthermore, miR-143 overexpression evidently attenuated in vitro 18 F-FDG uptake of 8505C and C643 cells at 120min (Fig. 3C), which can be utilized to detect and visualize glucose consumption. These results demonstrated that miR-143 inhibited glycolysis of ATC by negatively regulating HK2 expression. Then we examined the effect of miR-143 on tumor biological behavior in ATC cells using MTT, transwell migration, and flow cytometry assays. As a result, compared with the negative control groups, miR-143 overexpression markedly repressed the proliferative and migratory abilities of 8505C and C643 cells and further induced cell apoptosis (Fig. 3D–F), supporting the tumor-inhibitory effect of miR-143 on ATC cells.

miR-143 represses glycolysis of anaplastic thyroid carcinoma (ATC) by downregulating HK2 expression, inhibits proliferative and migratory abilities of ATC cells and induces apoptosis.
miR-143 exerted a tumor-suppressive effect on PDTC xenografts
Next we explored systemic treatment of PDTC xenografts with miR-143 agomir. The PDTC tumor-bearing mice were intravenously injected with Cy3-miR-143 agomir or Cy3-Ctrl RNA agomir which were encapsulated into a neutral lipid emulsion and disrupted by a sonicator successively. When the tumor volumes reached around 100–130 mm3, the formulation was administered once every 3 days for a total of six treatments. 18 F-FDG microPET/CT was performed before the first treatment and after six treatments (Fig. 4A). Tumor volumes were measured during the treatment cycle. The tumor growth curve revealed that miR-143 markedly slowed the rates of tumor growth in PDTC xenografts (Fig. 4C). After six treatments, tumors from both the miR-143-treated and negative control groups were resected for further assays. As qRT-PCR assays proved (Fig. 4B), miR-143 was remarkably overexpressed in the treatment group. Additionally, results of immunohistochemistry assays showed that proliferation-related gene Ki-67 was markedly downregulated in the miR-143-treated group compared with the negative control group, whereas a distinct upregulated expression of apoptosis-related gene Caspase-3 was observed in the tumors of treatment group (Fig. 4D). Altogether, the above results further support the growth inhibitory role of miR-143 in PDTC xenografts.

Systemic treatment of PDTC xenografts with miR-143.
Evaluating the therapeutic effect of miR-143 in PDTC xenografts using 18 F-FDG microPET/CT
18 F-FDG microPET/CT scanning was performed for further visualization of the therapeutic efficacy of miR-143 in PDTC xenografts. As is shown in the representative images (Fig. 5A), mice treated with Ctrl RNA agomir exhibited a striking increase in tumor 18 F-FDG uptake, whereas in the miR-143-treated group, tumor 18 F-FDG uptake increased slightly. Once the ROIs were delineated, SUVmax and SUVmean were acquired for semiquantitative analysis. Following six treatments, the miR-143-treated group showed a mild change in both SUVmax and SUVmean at the terminal point. Moreover, as can be seen from the post-treatment results, the miR-143-treated group revealed a significantly lower level of both SUVmax and SUVmean in PDTC xenografts, compared with the negative control group (Fig. 5B). Additionally, as shown in the representative images acquired by microPET/CT, the tumor size of miR-143-treated xenografts was smaller than that of the negative control group (Fig. 5A), further confirming the inhibitory effect of miR-143 on PDTC. Subsequently, tumors from both groups were resected for immunohistochemistry assays. The results showed that the expression of glycolysis-related gene HK2 was distinctly downregulated in the miR-143-treated group (Fig. 5E). Moreover, compared with the negative control group, tumors of miR-143-treated group exhibited a downregulation of HK2 mRNA and protein expression which were separately verified by qRT-PCR and Western blot (Fig. 5C and D). Collectively, the above results support the feasibility of evaluating the therapeutic effect of miR-143 in PDTC xenografts using 18 F-FDG microPET/CT.

Assessment of miR-143-exerted therapeutic effects on PDTC xenografts using
18
F-FDG microPET/CT.
Toxicity evaluation of miR-143 agomir treatment in PDTC
We next assessed the therapeutic toxicity of miR-143 agomir by measuring the weights of mice every 3 days throughout the treatment cycle and carrying out clinical chemical tests at the end of the therapy. When the treatments were finished, tumor tissues and the organ tissues were further resected from the PDTC tumor-bearing mice to assess the distribution of miR-143 agomir. As shown in Fig. 6A, there is a slight fluctuation of body weights in all the three groups of mice. However, no significant weight loss was observed in the miR-143-treated group compared with the controls. The results of clinical chemical tests showed that therapeutic delivery of miRNA agomirs caused a mild decrease in white blood cells counts and platelets, while a slight rise of aspartate aminotransferase and urea were observed compared with the PBS-treated group. This might result from the chemistry of agomir or liposome. Furthermore, therapeutic delivery of encapsulated miR-143 agomir slightly reduced hemoglobin and elevated alanine aminotransferase, compared with both controls (Fig. 6C). Despite the changes caused by miR-143 formulation, there is no significant difference among the groups, indicating that therapeutic delivery of miR-143 has minimal toxicity on PDTC xenografts. We further examined the distribution of miR-143 agomir in different tissues by detecting Cy3 signals. As we expected, an evident distribution of Cy3-miR-143 agomir was observed in tumor tissues. Moderate formulation was found to scatter in lung, liver, kidney, and stomach tissues, while only slight fluorescence was detected in heart, brain and muscle tissues (Fig. 6B).
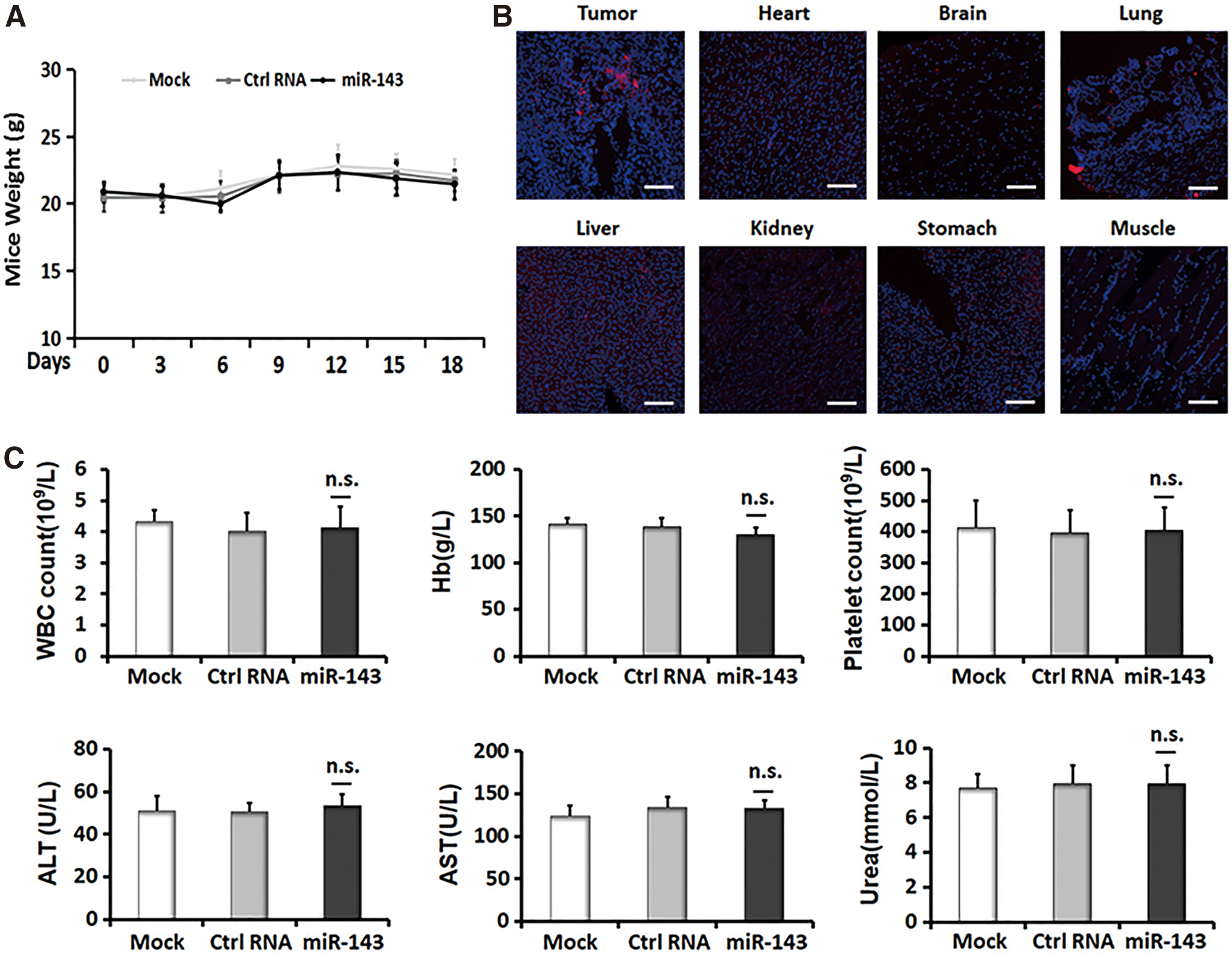
Toxicity assessment of systemic treatment of PDTC xenografts with miR-143.
Discussion
Elevated glucose metabolism proves to be one of the distinctive traits of cancer cells, providing the raw material for further anabolic processes to produce nucleic acids, proteins, etc. that cancer cells demand. Multiple glycolytic enzymes are aberrantly expressed in tumor cells to maintain the micro-environmental homeostasis, leading to an extensive exploration of targeting glycolytic enzymes as anti-cancer strategy. 22,23 Ja Seung Koo et al. recently screened glycolysis-related protein expression in different subtypes of thyroid tumors. 24 HK2-positive proportion was observed higher in PDTC than in DTC. Besides, elevated HK2 expression was also observed in follicular thyroid carcinoma (FTC) compared with follicular adenoma. The results are in accord with the “flip-flop phenomenon,” which has been observed in the process of thyroid cancer dedifferentiation, enabling the tumors with decreased avidity for RAI and increased glucose metabolic activity. The above studies support our idea of treating PDTC by targeting HK2 to interfere with glucose metabolism. Members of the miR-143 family were involved in multiple signaling pathways by targeting different genes. Guo et al. found that miR-143 suppressed the proliferation and metastasis of endometrial cells by targeting MAPK1, which modulated CYCLIN D1, c-MYC, etc., the downstream substrates of the MAPK/ERK pathway. 25 Fan et al. showed that miR-143 and miR-145 regulated the epithelial–mesenchymal transition process and depressed the metastasis of gastric cancer by targeting MYO6. 26 Additionally, GATA6, CIAPIN1, and FOSL2 were all demonstrated to be the target genes of miR-143. 27 –29 In the current study, we examined whether the above genes could be regulated by miR-143 in PDTC using qRT-PCR assays. As a result, mRNA expression levels of FOSL2 and CIAPIN1 were significantly suppressed by miR-143 both in vitro and in vivo (Supplementary Fig. S1). FOS-like antigen 2 (FOSL2), member of the activator protein 1 transcription factor family, is closely related to cell proliferation, adhesion, invasion, and metastasis. 29,30 As a novel apoptosis inhibitor, cytokine-induced apoptosis inhibitor 1 (CIAPIN1) depletion proves to inhibit proliferation and trigger more apoptosis of chronic myeloid leukemia cells, further decrease ERK5 phosphorylation and NF-κB activity. 31 These findings support the importance of FOSL2 and CIAPIN1 in the biological behavior of tumors, suggesting that miR-143 could probably exert a tumor-suppressive effect by targeting FOSL2 and CIAPIN1 in PDTC.
In our previous work, HK2 proved to be the target gene of miR-143 in triple-negative breast cancers. 17 This finding was also verified in multiple tumors such as oral squamous cell carcinoma, prostate carcinoma, and renal cell carcinoma. 32 –34 Our further studies indicated that systemic delivery of miR-143 agomir targeting tumor metabolism was an effective anti-tumor strategy in triple-negative breast cancers. 35 PDTC often exhibits a characteristic of “increased glycolysis,” which could be confirmed by 18 F-FDG PET/CT. 8 However, it is unknown whether miR-143-HK2 axis is related to the elevated glycolysis of PDTC. In the current research, we noticed that compared with normal thyroid cells, PDTC cells harbor the reduced level of miR-143 and increased mRNA and protein expression of HK2 (Supplementary Fig. S2). These results indicate that abnormal glycolysis of PDTC is associated with the lack of miR-143. Overexpression of miR-143 was found to reduce HK2 expression remarkably, consistent with the attenuation of the rates of glucose consumption and lactate production as well as G6P, ECAR, and in vitro 18 F-FDG uptake. The proliferative and migratory abilities of PDTC cells were dramatically inhibited, while the cell apoptosis was observed to significantly increase. We further confirmed the suppressive effects of miR-143 on glycolysis and tumor biological behavior in 8505C and C643 ATC cells, which are highly aggressive and not sensitive to RAI, belonging to advanced thyroid carcinoma resembling PDTC. Moreover, the in vivo therapeutic delivery of miR-143 verified the glycolysis-suppressive and tumor-restrained role of miR-143 in PDTC xenografts.
18 F-FDG PET/CT scanning has been regarded as an efficient imaging tool. It has been widely exploited for tumor detecting, staging, restaging, monitoring the response of patients in anti-cancer therapy, and predicting the prognosis. In thyroid cancer cases, 18 F-FDG PET/CT scanning was preferentially recommended in high-risk DTC patients with elevated serum Tg (generally >10 ng/mL) and negative RAI imaging. Additionally, 18 F-FDG PET/CT scanning is also recommended in patients with an aggressive histological subtype, such as poorly differentiated, tall cell and Hürthle cell thyroid cancer. 36 Studies showed that the sensitivity and specificity of 18 F-FDG PET/CT in detecting the non-131I-avid thyroid cancer lesions were as high as 80% or more. 37 Besides, 18 F-FDG PET/CT could sensitively prompt the dedifferentiation tendency of DTC lesions. 38 In our present work, the therapeutic efficacy of miR-143 in PDTC xenografts was assessed using 18 F-FDG PET/CT scanning. Our results indicated that six treatments with Cy3-miR-143 agomir dramatically suppressed tumors 18 F-FDG uptake (reflected by SUVmax and SUVmean) compared with the negative control group, while the values showed little change compared with the pretreatment in the miR-143-treated group. Our findings were verified by immunohistochemistry, qRT-PCR, and Western blot assays, proving that HK2 was inhibited by miR-143, further supporting that HK2 is an independent predictor of 18 F-FDG uptake. 39 Furthermore, tumors observed in the miR-143–treated group grew slower than that in the negative control group. This observation coincides with the upregulation of Caspase-3 marker and the downregulation of Ki-67 marker in the stained tumor sections. Altogether, the systemic treatment efficacy of miR-143 in PDTC xenografts could be specifically assessed by 18 F-FDG microPET/CT.
To sum up, the potentiality of miR-143 as a therapeutic target for PDTC was demonstrated in our current study. Moreover, the efficacy evaluation of miR-143 treatment in PDTC xenografts could be achieved by 18 F-FDG microPET/CT. Further investigation should be carried out to take the different subtypes and molecular features of thyroid cancer into consideration. And the problem of targeting property requires further exploration.
Footnotes
Acknowledgments
This work was supported by grants from the National Natural Science Foundation of China (Nos. 81271610 and 81501499).
Author Disclosure
No competing financial interests exist.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.