
Abstract
Reconstitution of healthy endothelial cells in vascular beds offers opportunities for mechanisms in tissue homeostasis, organ regeneration, and correction of deficient functions. Liver sinusoidal endothelial cells express unique functions, and their transplantation is relevant for disease models and for cell therapy. As molecular targets for improving transplanted cell engraftment and proliferation will be highly significant, this study determined whether ETA/B receptor antagonism by the drug bosentan could overcome cell losses due to cell transplantation–induced cytotoxicity. Cell engraftment and proliferation assays were performed with healthy wild-type liver sinusoidal endothelial cells transplanted into the liver of dipeptidylpeptidase IV knockout mice. Transplanted cells were identified in tissues by enzyme histochemistry. Cells with prospective ETA/B antagonism engrafted significantly better in hepatic sinusoids. Moreover, these cells underwent multiple rounds of division under liver repopulation conditions. The gains of ETA/B antagonism resulted from benefits in cell viability and membrane integrity. Also, in bosentan-treated cells, mitochondrial homeostasis was better maintained with less oxidative stress and DNA damage after injuries. Intracellular effects of ETA/B antagonism were transduced by conservation of ataxia telangiectasia mutated protein, which directs DNA damage response. Therefore, ETA/B antagonism in donor cells will advance vascular reconstitution. Extensive experience with ETA/B antagonists will facilitate translation in people.
Introduction
The ability of transplanted endothelial cells (EC) to replace proteins, repair vessels, and help regenerate organs is important for tissue engineering and cell and gene therapy. 1 –3 The hepatic microcirculation represents one of the largest vascular beds in the body. Understanding biological mechanisms in its reconstitution are of particular significance in view of the rising prevalence of liver diseases where pathophysiological processes include roles of EC. 4 Whereas multiple subsets of EC have been characterized for different functions in arteries, veins, or sinusoids within tissues, 5,6 liver sinusoidal endothelial cells (LSEC) represent a distinct cell type with unique properties. 4 Besides phenotypic variations from other EC, expression of coagulation factors (e.g., FVIII), cytokines (e.g., GCSF, VEGF, and others), or scavenger functions (e.g., hyaluronic acid receptors) are also characteristic of LSEC. 2,7 –9 Major roles of LSEC include support of hepatic development, homeostasis, and regeneration, 9 –11 and their losses in adult liver due to injury or inflammation during the course of diseases is harmful. 2,4,12,13 Injuries in LSEC may also arise due to anticancer or other drugs, toxins, and radiation. Such vascular damage may be restrictive for completing (cancer) therapies and might lead to disabling liver conditions (e.g., sinusoidal obstruction syndrome). 4 Transplantation of healthy LSEC to repair endothelial damage in these and other situations could be helpful. The potential of LSEC transplantation for correcting FVIII deficiency in hemophilia mice has been established. 2 However, for effective reconstitution of vascular beds, superior engraftment and proliferation of transplanted cells will be critical.
Vasoconstrictors, cytokines, chemokines, or receptors are major contributors to endothelial damage in tissues. 2,13 –18 Among these, the most potent vasoconstrictor, endothelin (ET)-1, has been particularly noteworthy. 19 Expression of ET-1 leads to considerable ischemic injury in multiple tissues. 13,20 –24 The role of ET-1 in tissue damage led to characterization of ETA and ETB receptors and development of several antagonists. 19 Of these, ETA/B antagonism by bosentan (BOS) is effective for treating pulmonary hypertension, cardiac hypertrophy, and other conditions. 25 –27 Experimental studies in conditions of hypoxia and inflammation with antagonism of ETA (darusentan) or ETA/B (BOS) indicated that these drugs may protect transplanted hepatocytes. 21,22 Expression of ETA/B in LSEC has been verified by autoradiographic localization of radiolabeled ET-1, including in receptor subtype knockout mice. 28 Nonetheless, whether ETA/B antagonism may benefit transplanted EC has been unknown.
Normally, ET-1 is expressed at low levels to maintain vascular tone. 19,29 Excessive amounts of ET-1 are removed by intracellular uptake via ETB receptors, although this causes oxidative stress (as in conditional EC-specific transgenic mice). 30 –32 Moreover, ET-1 may damage mitochondria (as in aortic smooth muscle). 33 Without healthy mitochondria, cell survival and cell division are impaired. 34 Depletion of mitochondrial DNA may even cause organ failure. 35 –38 Recently, ataxia telangiectasia mutated (ATM) protein, which oversees DNA protection, has been associated with mitochondrial homeostasis. 39 –41 This molecular context should be significant for the role of ET-1 in transplanted cells.
To this end, studies were performed by transplanting donor LSEC with or without ETA/B antagonism into knockout mice lacking dipeptidyl peptidase (DPP)-IV activity, since these offer convenient assays for liver repopulation. 15 The study determined whether ETA/B antagonism in donor cells will improve vascular reconstitution with attention to the following mechanisms: (1) by improving endothelial viability under stress or injury conditions; (2) by enhancing engraftment and proliferation of transplanted cells; and (3) by exerting effects on mitochondrial health and DNA damage response related to the ATM pathway. These aspects revealed ETA/B antagonism was highly significant for reconstituting hepatic sinusoids.
Methods
Drugs and chemicals
Drugs, chemicals, and reagents, including phenytoin (Phen), rifampicin (Rif), monocrotaline (MCT), naproxen (Nap), and thalidomide (Thal), were obtained from Sigma–Aldrich (St. Louis, MO). Water-soluble sodium salt of BOS was provided by Actelion Pharmaceuticals (Allschwil, Switzerland). Drugs were dissolved in normal saline, except for Nap, which was dissolved in 20% ethanol (2 mg/mL), and Thal, which was dissolved in dimethylsulfoxide (25 mg/mL).
Animals and procedures
Animal Care and Use Committee of Albert Einstein College of Medicine approved protocols in compliance with the National Institutes of Health and other regulations.
Donor C57BL/6 mice, 8–10 weeks old, were from National Cancer Institute (NCI; Bethesda, MD). DPP-IV−/– mice, 6–7 weeks old, were from Animal Models, Stem Cells, and Cell Therapy Core of the Liver Research Center. Mice were housed under 16 h/8 h light/dark cycles with unlimited access to pelleted chow and water. Male and female mice were distributed in equal numbers for cell engraftment studies. Mice received 20 mg/kg Thal or 9 mg/kg Nap 2h before and 200 mg/kg MCT 16–24 h before cell transplantation. Males were obligatory for liver preconditioning in repopulation assays. This required Phen, Rif, and MCT over 4 days, as previously described. 15
Mouse anesthesia used ketamine/xylazine. Mice received 0.75–1 × 106 LSEC in 0.25 mL serum-free Dulbecco's modified Eagle's medium (DMEM; GIBCO, Grand Island, NY) via the portal vein with 27-gauge needles. Hemostasis was secured by compression. For Kupffer cell (KC) phagocytosis, carbon was given 1 h before sacrifice, as previously described. 17
LSEC isolation
Multiple donors were used. The liver was perfused via the portal vein with 5 mL/min of collagenase (Worthington Biochemical Corp., Lakewood, NJ) or Liberase (Thermo Fisher Scientific, Fredericks, MD), as described. 42 Hepatocytes were removed by centrifugation. Gradients of Percoll™ (Sigma–Aldrich) were used to isolate purified LSEC, as previously described. 8 Initial viability was determined by trypan blue dye. Cells were examined by glycogen staining (hepatocytes), retinoid autofluorescence (hepatic stellate cells [HSC]), and F4/80 staining (KC), as previously described. 43 For CD31 immunostaining, cells were fixed in cold acetone for 10 min, and incubated in PE-conjugated anti-mouse CD31 antibody (1:100; Pharmingen BD Biosciences, San Diego, CA). Nuclear staining used Hoechst 33258 (Sigma–Aldrich).
Initial LSEC viability by trypan blue dye exclusion was 75–90%. The majority of LSEC were CD31+ (75–80%), which was similar to cell profiles previously described. 43 Cell isolates did not contain any hepatocytes or HSC. Presence of contaminating KC was infrequent (2–5%). As engraftment in liver of transplanted KC is inefficient and these do not proliferate, their presence was inconsequential. 12
ETA/B receptor antagonism by BOS and time-to-transplantation
Immediately after isolation, LSEC were incubated with gentle mixing in serum-free DMEM containing 10–5 M BOS for 60 min at 4°C on ice. 13,21,23 After incubating, free BOS was removed by washing cells twice in plain DMEM. Control cells were incubated in DMEM alone. Cells were transplanted within 1 h after isolation and drug treatment. There was no intervening step of culture.
Cell viability and cytoskeletal integrity in vitro
All experiments were repeated at least three times from separate donors. To identify live and dead cells, incubation for 10 min with 1 μg/mL propidium iodide (PI; Sigma–Aldrich) was used. Assays for cell attachment used 4 × 105 LSEC/cm 2 of collagen-coated dishes in Medium-199 (Sigma–Aldrich) with 100 μg/mL Endothelial Cell Growth Factor and 20% fetal bovine serum (M-199) for 4 h. Cells were counted by Operetta high-content imaging (PerkinElmer, New York, NY). For F-actin and vinculin staining, cells were plated in collagen-coated dishes for 2 h. For F-actin, cells were fixed with ethanol, permeabilized with 0.1% Triton X-100 (Sigma–Aldrich) in phosphate-buffered saline (PBS) for 10 min, blocked in 3% bovine serum albumin (BSA) for 30 min, and then incubated with phalloidin in 1% BSA (1:250, Phalloidin Cruz Fluor™ 488 Conjugate; S.C-363791; Santa Cruz Biotechnology, Santa Cruz, CA) for 30 min. For vinculin, cells were incubated with anti-vinculin in 1% BSA (1:250, NB600-1293; Novus Biologicals, Littleton, CO) overnight at 4°C followed by TRITC-conjugated anti-mouse immunoglobulin G (IgG; 1:250, T5393; Sigma–Aldrich) for 1 h at room temperature. Nuclei were counterstained by Hoechst 33258.
For angiogenic tube formation, 1 × 105 LSEC were plated per cm 2 in Matrigel-coated 24-well dishes in M-199 medium (see above). Cells were maintained in culture for up to 48 h. Multiple microphotographs were obtained after overnight culture, 24 h, and 48 h. Cell migration from early clusters, intermediate tube-like structures, and characteristic angiogenic tubes were scored in microphotographs. Each condition was represented in three to six replicates.
Mitochondrial membrane potential and analysis of oxidative stress with reactive oxygen species
LSEC were seeded at 5–8 × 104 per well in 96-well collagen-coated dishes in DMEM for 1–2 h. Oxidative stress was induced by exposure to 0.01–1 mM hydrogen peroxide (H2O2) for 1 h in 5% CO2 at 37°C. To detect reactive oxygen species (ROS), cells were incubated for 1 h with 25 μM dihydrorhodamine 123 (Sigma–Aldrich), and microphotographs for green fluorescence were obtained by Axiovision microscope (Carl Zeiss Microscopy LLC, Thornwood, NY). For mitochondrial membrane potential (MMP), cells were incubated with JC-1 dye (Cayman Chemical, Ann Arbor, MI) for 1 h at 37°C according to the manufacturer's instructions, plus 6 μg/mL Hoechst 33258 for staining nuclei. Aggregation of JC-1 was recorded by shift from green to red epifluorescence with microphotography. Image analysis used a Cytation5 instrument. To identify the role of ATM, its kinase activity was inhibited by 10 mM caffeine (Sigma–Aldrich) or 10 μM of either KU-55933 or KU-60019 antagonists (Cayman).
DNA damage pathways
Cells were fixed in ethanol for cytostaining. For visualizing 8-oxo-dG adducts, cells were treated with 250 ng/mL RNAse for 1 h at 37°C; DNA was denatured in 4 M HCl for 7 min followed by neutralization with 50 mM Tris base for 2 min. After blocking with 10% goat serum (Life Technologies, Frederick, MD) in PBS for 1 h, cells were incubated with mouse anti-8-oxo-dG (1:1,000, cat-4354-MC-050, Clone 2E2; Trevigen, Inc., Gaithersburg, MD) overnight. Then Alexa Fluor® 647-conjugated goat anti-mouse IgG (1:100, cat. #4410; Cell Signaling Technology, Danvers, MA) was applied for 1 h at room temperature. For other stainings, fixed cells were blocked/permeabilized in 5% goat serum, 0.2% Triton X-100 (Sigma–Aldrich) in PBS for 1 h, and incubated overnight at room temperature with anti-phospho-ATM (1:100, 560007; BD Pharmingen, San Diego, CA), phospho-Chk2 (1:100, ab59408; Abcam, Cambridge, United Kingdom), or anti-phospho-NBS1 (1:100, PA5-34385; Thermo Fisher Scientific) followed by Alexa Fluor®647-conjugated anti-mouse or anti-rabbit IgG (Cell Signaling Technology). Cells were counterstained with DAPI. Imaging used an Axiovision microscope.
Tissue studies
Samples were frozen in methylbutane at −80°C for 5 μm cryosections. For carbon uptake in KC, ethanol-fixed sections were stained with hematoxylin. Histochemistry for DPP-IV was as described previously. 13,22,23 In some studies, DPP-IV staining was modified by fixing sections for 5 min in cold 95% ethanol/glacial acetic acid (99/1 v/v) on ice, followed by 5 min in 95% ethanol at 4°C. Samples were incubated in DPP-IV substrate for 30 min at 37°C, followed by 3 min in 0.1 M CuSO4. This was followed by post fixing in 4% paraformaldehyde. Nuclei were stained with toluidine blue (Sigma–Aldrich).
Morphometric analysis
For cell culture assays, multiple images were analyzed per condition with 200–1,000 cells each. Animal groups comprised multiple mice (n = 6 each). In each mouse, three sections from same liver lobes were scored with 50–100 microscopic fields in multiple animals per condition (n = 3–6). Carbon-containing KC were counted under 200× magnification. Transplanted cells were counted under 400× magnification. Liver areas with transplanted cells were measured under 100× and quantitated by ImageJ (NCI). Areas of transplanted cell foci were analyzed by quantile distributions with box plots. Linear regression for liver repopulation kinetics was as described previously. 13
Statistical methods
Each experimental analysis used three to six mice. Data are shown as means ± standard error of the mean. Significances were determined by a t-test, Mann–Whitney rank sum test, analysis of variance with Tukey's test, or Spearman's correlation with GraphPad Prism v7 (GraphPad Software, Inc., La Jolla, CA). p-Values of <0.05 were considered significant.
Results
Superior performance of donor cells for engrafting in the liver should have been apparent with assays for viability and cytoskeletal integrity under cytotoxic conditions. The former was elucidated by demonstrating live cells under oxidative stress following exposure to H2O2; the latter was examined by cell attachment to extracellular matrix components (ECM) and cell aggregation with production of angiogenic structures.
ETA/B antagonism improved LSEC viability in culture
In BOS-treated LSEC versus untreated controls, live or dead cell assays with PI revealed more live cells under basal conditions immediately after drug treatment (Supplementary Fig. S1). When cells were exposed to H2O2, BOS-treated LSEC maintained greater viability compared to control cells (p < 0.05, ANOVA, n = 3 replicates). This indicated ETA/B antagonism imparted greater robustness to LSEC.
After culture in dishes, LSEC attached well. Virtually all cells, both with or without BOS treatment stained for CD31, thereby excluding contaminating cell types (Supplementary Fig. S2A). Testing for cell adherence in collagen-coated dishes indicated this was superior for BOS-treated LSEC (13 ± 3% greater vs. untreated control LSEC; p < 0.005; Supplementary Fig. S2B). Similarly, analysis of cytoskeletal microfilament structure by the pattern of F-actin distribution was informative. These actin filaments contribute toward maintenance of cytoskeletal structure in a dynamic fashion that allows cell migration and shape alterations during cell contractions or division. These cytoskeletal structures are also critical for physiological trafficking of intracellular molecules. In drug-untreated control LSEC, F-actin was either not localized or was mostly disrupted. By contrast, BOS-treated LSEC exhibited F-actin in far more cells—encircling cells either partially or fully throughout cytoskeletal areas (Supplementary Fig. S3). This indicated superior preservation after BOS treatment of cell integrity.
To determine whether superior viability and cytoskeletal integrity will lead to activation of outside-in signaling after cell attachment, development of focal adhesion complexes was examined. These are formed by engagement of ECM (integrin) receptors and recruitment of intracellular kinases, such as vinculin. 44 Prevalence of vinculin-containing focal adhesion complexes was low in untreated LSEC (Supplementary Fig. S4). By contrast, focal adhesion complexes in BOS-treated LSEC increased by several fold (p < 0.02).
These cytoskeletal effects of ETA/B antagonism were further examined in Matrigel angiogenesis assays, where cell migration and cell membrane reorganization are required. After attachment to Matrigel, BOS-treated LSEC exhibited greater ability than untreated control LSEC to organize in clusters and early- or later-stage angiogenic tubes over 16–48 h (increased by 5.4 ± 0.6-fold; p < 0.05, ANOVA, n = 3; Supplementary Fig. S5). This established that BOS stabilized multiple events and processes in LSEC that together had major effects on their viability and performance under adverse conditions, setting the stage for testing in vivo.
Engraftment and proliferation assays in DPP-IV knockout mice
For engraftmen analysis, liver preconditioning was compared to established conditions of MCT (liver endothelial disruption), 15 Thal (anti-cytokine plus endothelial disruption), 13 or Nap (cytoprotective factor release). 24 To allow completion of cell engraftment, mice were analyzed 1 week after cell transplantation (Fig. 1A). In DPP-IV knockout mice, which lack DPP-IV enzyme activity, wild-type transplanted cells are readily identified. 15 In case of hepatocytes, DPP-IV enzyme activity is localized in bile canaliculi, which produces a distinct staining pattern in their apical domain away from the basolateral domain next to liver sinusoids, as noted in donor liver (Fig. 1B). By contrast, LSEC are located within hepatic sinusoids and express DPP-IV throughout the plasma membrane. The absence of background staining allowed localization of each DPP-IV+ transplanted LSEC. These engrafted LSEC exhibited characteristic elongated morphology in liver sinusoids. The distinct expression pattern of DPP-IV in the plasma membrane of LSEC versus bile canaliculi of hepatocytes allowed separation of these cell types. In mice preconditioned with MCT, transplantation of control LSEC resulted in engraftment of 0.5 ± 0.3 DPP-IV+ LSEC per portal area. Transplantation of BOS-treated LSEC in mice with preconditioning using Thal, Nap, or MCT increased engraftment. After MCT preconditioning, engraftment of BOS-treated LSEC was 1.5- and 3-fold greater than in mice with Thal or Nap preconditioning, respectively, and was 3.8-fold more than that of drug-untreated control LSEC, (p < 0.05, ANOVA, n = 6 each; Fig. 1C).

Engraftment of transplanted liver sinusoidal endothelial cells (LSEC) after 1 week.
To examine whether superior engraftment of BOS-treated LSEC could have resulted from modulation of hepatic inflammation after cell transplantation, KC activation was studied in cell recipients. The extent of KC activation as demonstrated by carbon uptake analysis increased similarly after transplantation of LSEC with or without BOS (Supplementary Fig. S6). This indicated that BOS treatment did not alter cell interactions in the liver. The ability of BOS-treated LSEC to engraft better, despite inflammation in the recipient liver, corresponded to their superior tolerance to injury in vitro.
To examine the effects of ETA/B antagonism and superior cell engraftment on subsequent proliferation of transplanted LSEC, liver repopulation assays were performed in mice preconditioned with Rif, Phen, and MCT (Fig. 2A). In this situation, proliferation in healthy transplanted LSEC is promoted by MCT-induced DNA damage in native cells that restricts their survival and/or cycling. 2 These effects in native cells are synergistically amplified by Rif and Phen. 15 Groups of mice were established for LSEC transplantation with or without BOS treatment (n = 6 each). After 6 weeks, tissue analysis for transplanted LSEC indicated these proliferated within hepatic sinusoids (Fig. 2B). Transplanted LSEC often lined entire sinusoids. Compared to drug-untreated controls, BOS-treated LSEC formed significantly larger areas (Fig. 2C). These areas in BOS-treated LSEC were 30 ± 10% larger than control LSEC (p < 0.001, t-test). The divergence in tissue areas replaced by BOS-treated and control LSEC indicated proliferation was accelerated in the former (p < 0.02, Spearman correlation; Fig. 2D). Overall, reconstitution of liver sinusoids by BOS-treated LSEC approached 10.4 ± 3.5% compared to 2.7 ± 0.5% in drug-untreated control LSEC (p < 0.008, t-test; Fig. 2E). Therefore, superior viability and cytoprotection after ETA/B antagonism had profound effects on LSEC proliferation. To define mechanisms underlying this benefit, the focus was on the effects of BOS on mitochondrial homeostasis.
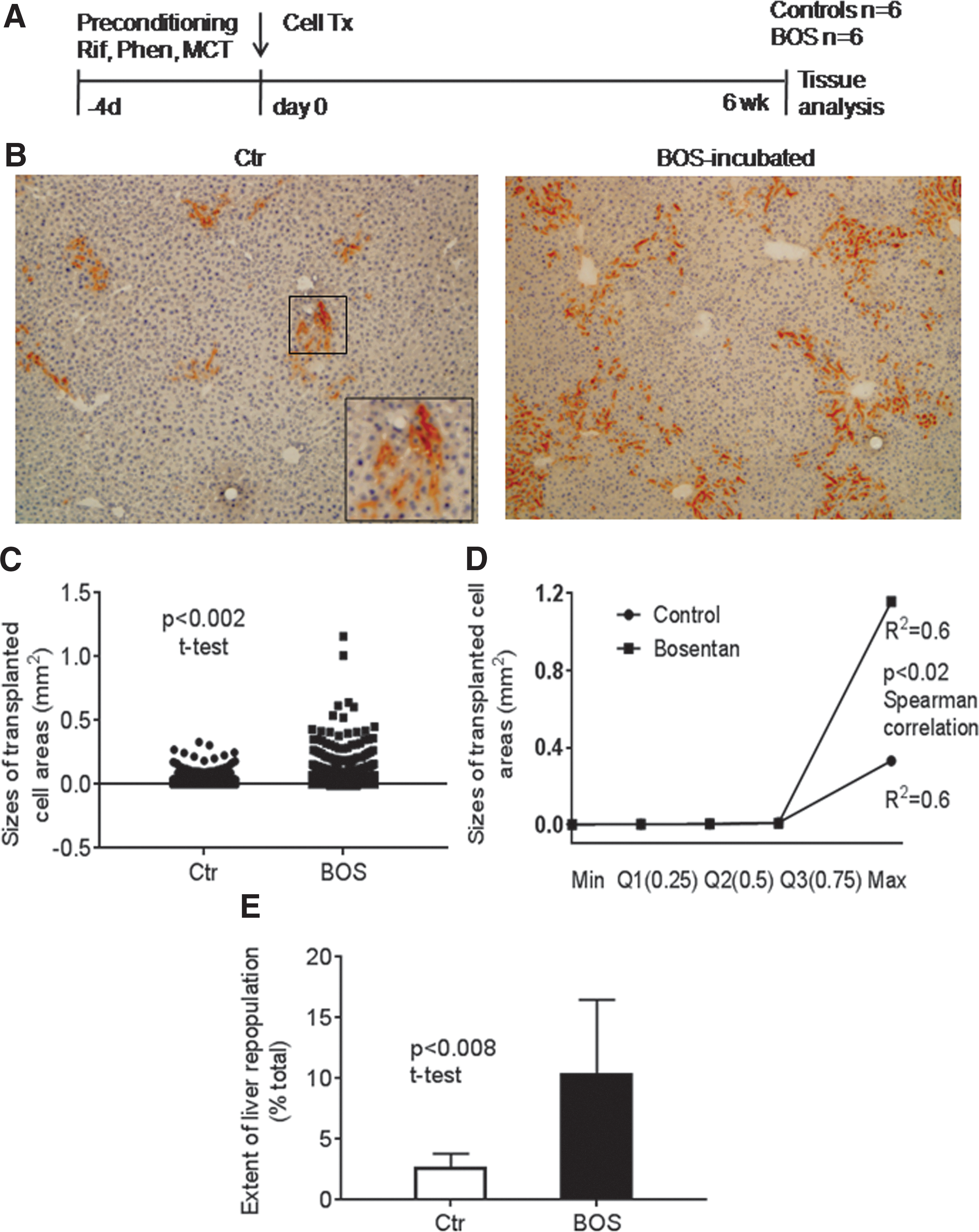
Liver repopulation kinetics in rifampicin (Rif)/phenytoin (Phen)/MCT-conditioned mice.
Mitochondrial viability and the ATM pathway were affected by ETA/B antagonism
As preservation of MMP is an excellent indicator of mitochondrial health, this property was examined first. Use of JC-1 dye for elucidating effect of oxidative stress, as previously demonstrated for other cell types, 45 indicated MMP levels in BOS-treated LSEC, even under basal conditions, were greater than control LSEC (Fig. 3A). Despite H2O2 injury, MMP levels were higher in BOS-treated LSEC compared to controls (p < 0.05, ANOVA, n = 3 replicates; Fig. 3B).

Analysis of mitochondrial membrane potential (MMP) levels with JC-1 dye in freshly isolated LSEC.
This was in agreement with effects of ETA/B antagonism in mitochondria. To substantiate this finding, metabolic activity of mitochondria was examined with dihydrorhodamine dye, which provides information about oxidizable substrates generating ROS. Under basal conditions, ROS levels were similar in control or BOS-treated LSEC (Fig. 4A.). With oxidative stress following H2O2 in control LSEC, ROS increased by several fold. However, BOS-treated LSEC withstood H2O2without increases in ROS levels (p < 0.0001, ANOVA). To reveal whether cells were protected from deleterious consequences of ROS, 8-oxo-dG adducts were examined, which result from oxidative DNA damage (Fig. 4B). In control LSEC, under basal and H2O2 conditions, 8-oxo-dG adducts were abundant, whereas these were significantly less in BOS-treated LSEC (p < 0.0001, ANOVA). Therefore, ETA/B antagonism improved mitochondrial health. This correlated with superior cell viability and cytoskeletal integrity in vitro and superior engraftment and proliferation in vivo.
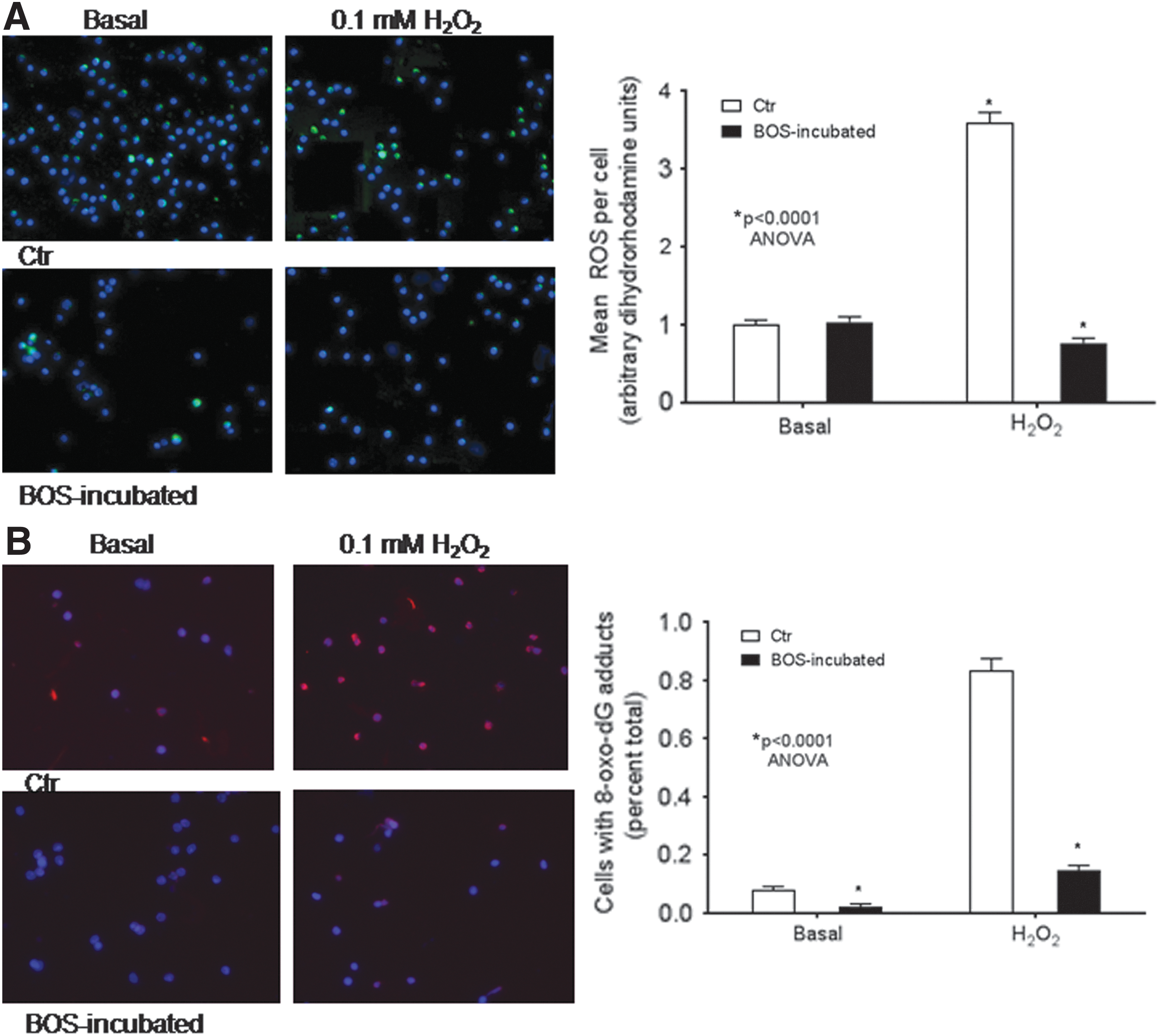
Effects of BOS on reactive oxygen species (ROS) and 8-oxo-dG DNA adducts in freshly isolated LSEC. The assays used LSEC with or without BOS treatment followed by culture over 1–2 h.
In DNA damage settings, the ATM pathway regulates cell survival and proliferation. 45 Recently, ATM and partners were found to be critical for mitochondrial biogenesis and also function. 39,40,46,47 To identify whether these processes were involved in ETA/B antagonism, phosphorylated ATM was localized, along with NBS1, a component of MRE11/RAD50/NBS1 complex in DNA double-strand breaks, and Chk2 kinase, a downstream ATM transducer and mitosis inhibitor after DNA damage. 48 In control LSEC, pATM was largely absent, and Chk2 or NBS1 were expressed in 12–15% under basal conditions with further increases after H2O2 (Fig. 5A and Table 1). By contrast, in BOS-treated LSEC, these events were reversed, with pATM in more cells and pNBS1 and pChk2 in fewer cells (p < 0.05, ANOVA; Fig. 5B and Table 1).
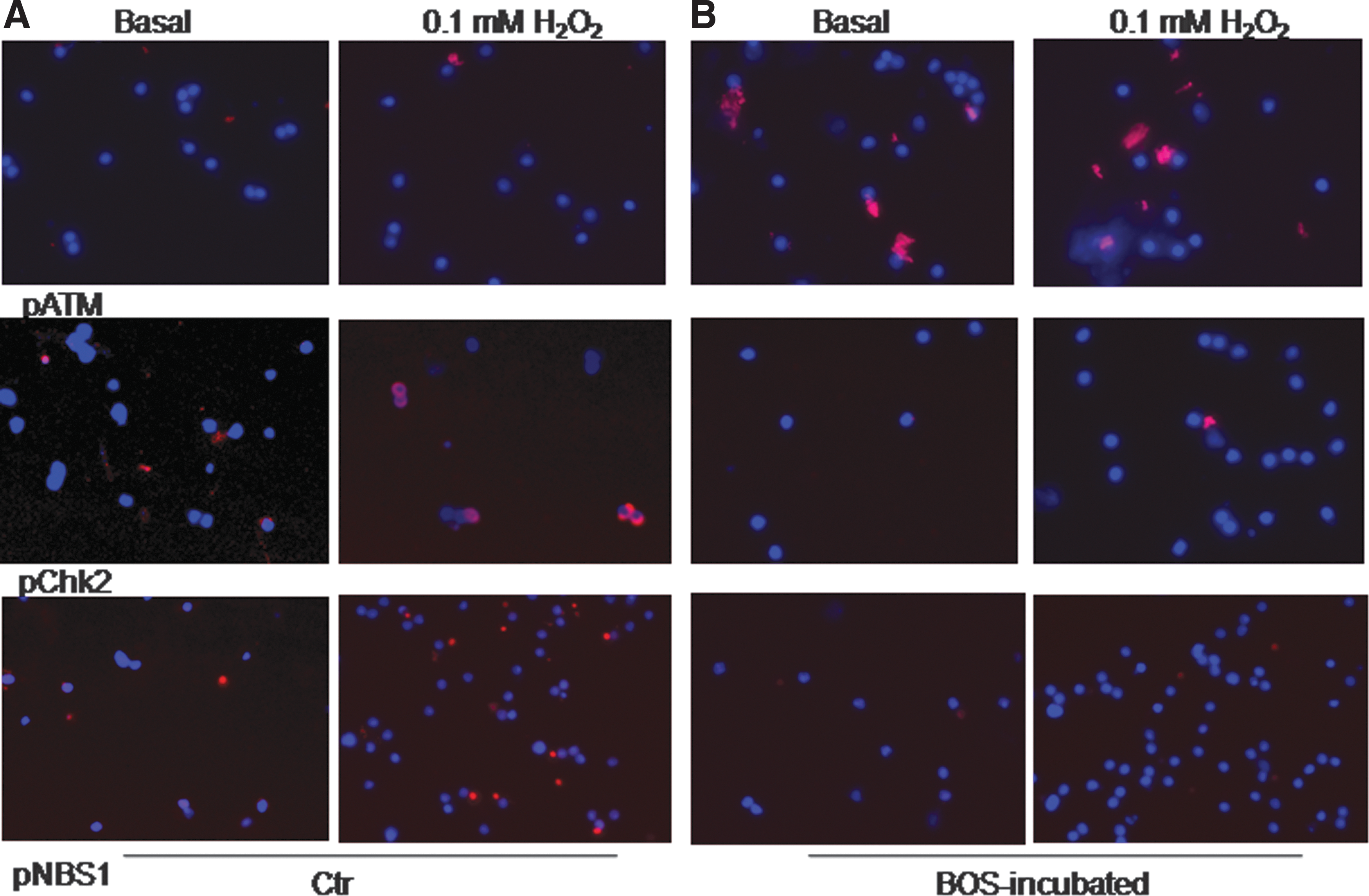
Effects of BOS on ATM pathway in LSEC. Microphotographs for pATM, pChk2, and pNBS1 expression in LSEC (red) under basal and H2O2 conditions. The assays used cells in culture over 1–2 h. Original magnification 400 × . Hoechst dye counterstain for nuclei.
Prevalence of cells with protein expression related to DNA damage response a
Morphometric analysis of multiple images per condition (n = 3 replicates).
BOS, bosentan; ANOVA, analysis of variance.
To reveal whether ATM accounted for mitochondrial homeostasis and superior MMP in BOS-treated LSEC, loss-of-function studies were undertaken with ATM kinase blockers. Inhibition of ATM kinase activity by any of 10 mM caffeine or 10 μM each of KU-55933 or KU-60019 antagonists markedly decreased MMP, thus verifying the direct role of ATM in mitochondrial homeostasis in BOS-treated LSEC (Fig. 6).

Effect of ATM kinase inhibition on MMP in LSEC with or without BOS.
Discussion
These studies provide firm evidence for protection of LSEC by ETA/B antagonism with BOS. This evidence includes maintenance of cell viability and cytoskeletal integrity under injury conditions. The benefits emanated from preservation of mitochondrial homeostasis by recruitment of the ATM pathway. These fundamental mechanisms related to ETA/B antagonism allowed transplanted LSEC to survive and proliferate better in mice.
The benefits of ETA/B antagonism in LSEC will have extended to interference in cytokine-mediated inflammation, which may be triggered by exposure of cells to ET-1 or other inflammatory mediators, as depicted schematically (Fig. 7). Importantly, inflammatory cytokines and growth factors may regulate the ATM pathway, along with other related events and processes during liver injury. 45,48,49 For instance, deficiency of ATM is responsible for damage to mitochondria, and characterizes a significant mechanism for failed liver regeneration in acute liver failure. 45 As loss of LSEC is integral to such liver injuries, restoring health to EC with drugs should be highly significant. The ability of drug-modified LSEC for reconstitution of liver will be significant for potential cell therapy applications in hepatic injury. This cell transplantation approach will be translatable, since BOS is widely used in clinical practice.
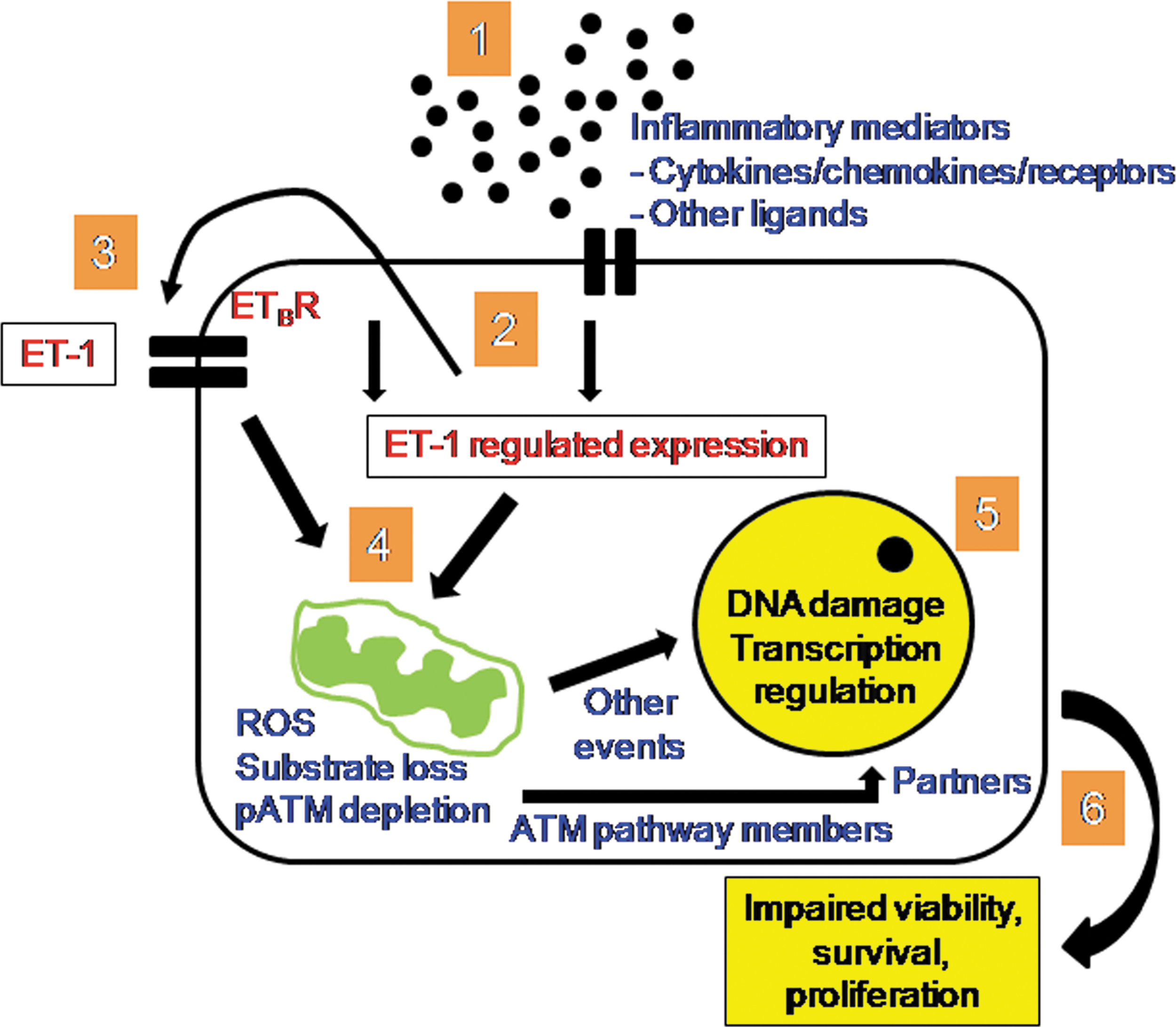
Representation of intracellular pathway effects in BOS-treated LSEC. In the setting of inflammation following exposure to cytokines, chemokines, and so on (1), BOS will have interfered with autocrine or paracrine effects of ET-1 through ETA/B receptor antagonism (2 and 3). This would decrease loss or depletion of oxidizable substrates and generation of intracellular ROS. Simultaneously, conservation of ATM protein would provide gains in MMP through recruitment of additional transducers and partners for mitochondrial homeostasis (4). Decreases in ROS and mitochondrial homeostasis would lead to avoidance of cellular and mitochondrial DNA damage, along with maintenance of physiological gene expression profiles (5). This would yield net gains for cell viability and cytoskeletal integrity culminating in better cell survival under adverse conditions and also acceleration of cell proliferation under suitable conditions (6).
Inflammatory effects of ET-1 are well recognized. 19 In case of EC, overexpression of ET-1 under endothelial-preferred promoters (Tie-1 or Tie-2) leads to increases in oxidative damage. 30 –32 Similarly, inflammatory cytokines induce ROS in isolated LSEC. These results are echoed in published findings obtained with FLS.12 cells, which originated from fetal mouse liver and express hematopoietic properties, since IL-3-dependent inflammation in these cells led to depletion of oxidizable substrates with impairments in mitochondrial function and also cell viability. 50 After repletion of oxidizable substrates, this damage reversed in FLS.12 cells. As ETA/B antagonism with BOS decreased oxidative stress and improved mitochondrial health in LSEC, this verified ET-1 was important for these processes.
Biogenesis and fission of mitochondria are essential requirements for cell cycle progression and completion of mitosis. 34 Drivers of mitochondrial fusion and fission (i.e., Drp-1 [drosophila] or DNM1L [human] and FIS1) are coordinately regulated during cycling by cell division kinase (CDK)-1/cyclin B or cyclin E. 51 Loss of Drp-1 may produce mitochondrial damage with hyperfusion and ATM-dependent cycling arrest in G2/M. 52 Interference in these processes from ET-1 is recognized in various cell types, where ET-1 overexpression increases mitochondrial fission. 33 Low MMP in LSEC may decrease cellular energy levels to cause arrest in G1/S. 51,53 As ETA/B antagonism improved mitochondrial health, these mechanisms should help explain superior engraftment and proliferation of BOS-treated LSEC.
Moreover, ATM directs mitochondrial homeostasis by regulating mitophagy.
39
Severe oxidative stress may by itself degrade ATM.
54
Thus, conservation of ATM in BOS-treated LSEC improved MMP and cell membrane
Superior engraftment of BOS-treated LSEC emphasizes the value of ETB antagonism for cell transplantation. Cells are exposed to inflammation during both isolation and transplantation. 18 Organ conditioning for cell transplantation introduces other adversities through tissue damage or inflammatory cell activations. 15 Therefore, superior outcomes in engraftment and proliferation of BOS-treated LSEC underscore the significance of cytoskeletal and mitochondrial integrity. Although BOS exerts antimitotic effects in cancers, 27,46 induction of proliferation in healthy cells should not be surprising, since cytoprotective mechanisms in healthy cells differ considerably from cancer cells.
The significant extent of liver repopulation (>10%) achieved after transplantation of BOS-treated LSEC should hold considerable value for therapeutic applications. For instance, several disorders of hemostasis will be amenable to either short-term or permanent support from transplanted healthy cells. Although gene therapy is advancing for hemophilia, cell transplantation will be significant for complex coagulation factor deficiencies, repair of vascular and tissue damage, as well as tissue engineering and neo-organogenesis. As multiple cell types may be transplanted simultaneously to reconstitute the liver, including hepatocytes, LSEC, KC, and HSC, 12,55 cell interactions may be studied in vivo. These studies of cell interactions will be advanced by efficient engraftment and proliferation of co-transplanted cells with the mechanism of ETA/B antagonism. This will facilitate translational applications.
The support of proliferation in transplanted LSEC despite only short-term BOS treatment was consistent with its prolonged effects on ETA/B receptors. 19 Recently, another attractive ETA/B receptor antagonist has become available (i.e., macitentan), which is an order of magnitude more potent than BOS due to greater binding affinity. Additionally, macitentan has no hepatic side effects compared to BOS, which inhibits the bile salt export pump and may cause liver injury due to retention of toxic bile salts. However, hepatic toxicity from BOS is uncommon, requiring systemic administration over prolonged periods. This will not be relevant if BOS is applied only to donor cells in vitro.
Footnotes
Acknowledgments
Supported in part by NIH grants R01-DK071111 and P30-DK41296. N.Y. received the Raman Fellowship Award from University Grants Commission, Government of India. Bosentan was kindly supplied by Actelion Pharmaceuticals Ltd., a unit of Johnson & Johnson, New Brunswick, NJ.
Author Disclosure
The authors declare no competing financial interests exist.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.