
Abstract
Traditional gene therapy (gene replacement) has made a breakthrough in treating inherited diseases. Adeno-associated virus (AAV) has emerged as a highly promising vector with innate ability, boosting the development of gene replacement and gene targeting. With the recent advance of engineered nucleases that work efficiently in human cells, AAV mediated-genome editing with nucleases has raised hopes for in situ gene therapy of inherited and non-inherited diseases. Here, the applications of AAV-mediated genome editing are highlighted, and the prospect of AAV and nucleases that will render extension of such success in clinical gene therapy is discussed.
Introduction
Due to the detailed exploration of the human genome in recent decades, hundreds of causative genes of human genetic disorders have now been identified. This has been greatly facilitated by the rapid progress in next-generation sequencing technology, which has allowed the identification of disease-associated genes for many inherited diseases, especially monogenic diseases.
1
Traditional clinical therapeutic approaches such as surgery or small molecule-based therapeutics cannot effectively treat these diseases. Consequently, gene therapy has emerged as a revolutionary concept, as it can potentially directly alter and repair the pathogenic gene. To date, a series of adeno-associated virus (AAV)-based gene therapy clinical trials have been performed for monogenic conditions, including lipoprotein lipase deficiency,
2
hemophilia,
3
and choroideremia.
4
In addition, AAV-mediated gene therapy holds promise for the treatment of complex disorders, such as heart disease
5
and age-related macular degeneration (AMD).
6
More details of clinical trials involving AAV can be found online (
AAV is considered a very promising gene-delivery vector, and has been tested in >100 clinical trials carried out over the last 15 years. Notably, with the ability to engineer AAV vectors and the development of genome editing enzymes, including zinc-finger nucleases (ZFNs), transcription activator-like effector nucleases (TALENs), and clustered regulatory interspaced short palindromic repeat (CRISPR)-Cas9, innovative genome editing methods are being developed to treat inherited and non-inherited diseases. AAV can serve as a donor template for homologous recombination (HR) with remarkably high efficiency. Double-strand breaks (DSBs) triggered by nucleases is also an important factor that greatly boosts the efficiency of HR-mediated genome editing. 7
AAV's replication relies on a helper virus such as adenovirus or herpesvirus. Therefore, it is considered to be a safe vector in gene therapy. Compared to other viral or non-viral vectors, recombinant AAV (rAAV) genomes are maintained predominantly as episomal circular forms and possess sustained expression, low pathogenicity (immune response), versatile serotypes (tissue specificity), and highly infectious efficiency, resulting in widespread potential applications for both basic and clinical research (Table 1). In a clinical trial for the treatment of hemophilia B (NCT00979238), AAV8 vectors with an F9 transgene were well expressed and tolerated in patients without the detection of toxicity after 3 years of vector administration. 3 Consequently, by using AAV for the delivery of genome editing nucleases, new genome editing strategies will be developed with great potential to generate disease models and correct mutations underlying genetic disorders.
Comparison of different viral and non-viral vectors
AAV Structure and Serotypes
AAV is a non-enveloped, replication-defective, single-stranded DNA virus, which only replicates in the presence of a helper virus. It was discovered in 1965 as a contaminant during the preparation of a simian adenovirus, and classified into the Parvoviridae family due to its small diameter of approximately 22 nm (Fig. 1). 8 AAV is composed of three structural proteins (VP1, VP2, and VP3) and a linear, single-stranded DNA (ssDNA) genome of approximately 4.7 kb, which contains 145 nucleotides inverted terminal repeats (ITRs). The first 125 nucleotides of the ITRs are palindromic and of great importance for viral replication. Because of alternative splicing and the presence of multiple start codons, AAV cassettes have two open reading frames (ORFs): rep and cap. Rep encodes four overlapping proteins (Rep78/Rep68 and Rep50/Rep42), which are responsible for replication, virion assembly, and site-specific DNA integration, as well as translational regulation. Cap encodes three structural proteins (VP1, VP2, and VP3) and a viral assembly factor (assembly-activating protein [AAP]), which contribute to the icosahedral capsid with the coordination of AAP (Fig. 1). 9
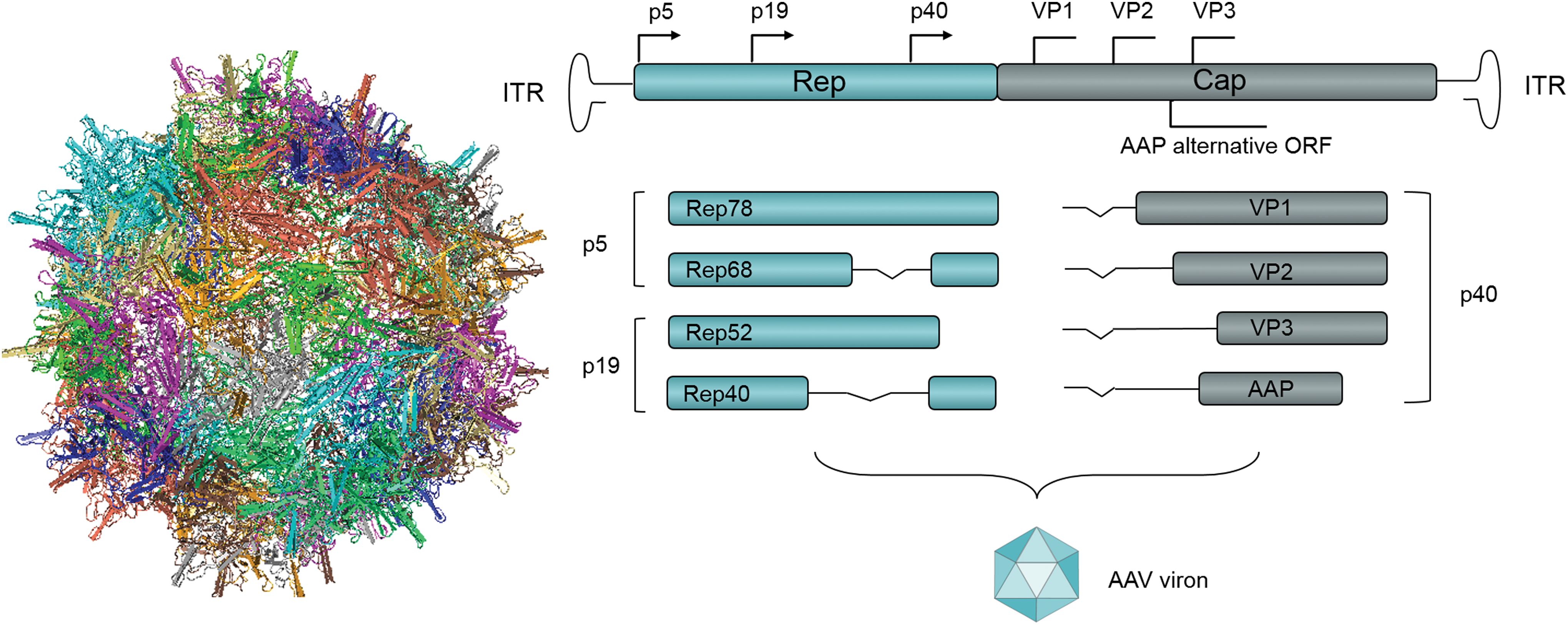
The structure of wild-type adeno-associated virus. Color images are available online.
Because of the diversity of its serotypes, which target various cell-surface receptors, AAV can infect different cell types with variable tissue tropism. Up to now, 12 different serotypes have been well characterized (AAV1–AAV12), and >100 new serotypes have been discovered more recently. The discovery of tissue tropism provides the basis for the selection of tissue-selective AAV vectors for targeted gene therapy (Table 2).
Tissue tropism of various AAV serotypes
AAV, adeno-associated virus.
AAV-Mediated Non-Homologous Integration and HR
Previous studies have demonstrated that wild-type AAV can integrate into a site at chromosome 19q13.4, termed the AAVS1 locus, in the absence of a helper virus. 10 More than 70% of integration events preferentially occur at this site via DSB and non-homologous end joining (NHEJ) pathways, which can induce insertions, deletions, mutations, or chromosomal translocations. Rep78/Rep68 play a major role in DSB, which can mediate the complex formation between AAV ITRs and the chromosome 19 integration site. 11 AAVS1 is considered a safe harbor for the integration of a foreign gene. Therefore, antibiotic resistance genes (e.g., neo) or fluorescent genes (e.g., eGFP) can be integrated into the AAVS1 locus for further studies, which can provide convenient markers for the specific selection of transduced cells. 12 AAV-mediated non-homologous integration enables the precise engineering of new cell lines. Moreover, rAAVs do not cause genomic breaks themselves, which is a benefit for their clinical use. 13 But rAAVs can still integrate at existing DSBs sites at the same time. 13 However, to reach more precise gene targeting or gene therapy, HR may be the better choice rather than AAV-mediated non-homologous integration.
HR is an accurate genome modification method, although it generally occurs at a low efficiency of 10–8–10–6. Compared to traditional methods based on transfection or electroporation of plasmid constructs, AAV-mediated site-specific integration frequency is 1,000-fold higher, reaching approximately 1% at the single-copy hypoxanthine phosphoribosyl transferase (HPRT) locus in normal human fibroblasts. 14 The HR frequency also varies in mammalian cells, depending on the genetic locus. 15 Additionally, HR frequency is sensitive to single-nucleotide polymorphisms (SNPs), and even a single mismatch can alter HR frequency. Hence, SNPs can be harnessed for directing allele-specific targeting in certain situations. 16
By altering the DNA sequence between homology arms, site-specific HR can be achieved. AAV-mediated HR has been shown to be able to introduce DNA substitutions, insertions, and deletion mutations with high fidelity at multiple chromosomal positions. 14,17 However, there is a preference for HR to occur at active transcriptional units because of the exposed single DNA strand. 18 To deliver an additional gene to correct mutations or to produce disease models for basic research or preclinical gene therapy, AAV methodology is of great value. Hirata et al. used AAV to introduce a 1.5 kb transgene cassette into HPRT and COL1A1 (type I collagen) loci in normal human fibroblasts with an efficiency of 1%. 19 Miller et al. showed that AAV can accurately correct chromosomal mutations in vivo at frequencies of 10–4–10–5 in hepatocytes. 20 Precise gene correction was observed by injecting AAV vectors carrying a lacZ fragment using the ROSA26 knock-in mouse with a missense mutation in lacZ. 20 Furthermore, a Duchenne muscular dystrophy (DMD) mouse model (mdx) was created by HR following intravascular co-delivery with two mini dystrophin-encoding rAAV6 vectors. 21 In order to reach the standard required for clinical therapeutic applications, the efficiency of HR is a crucial problem that must be overcome. As a result, new rAAV-rDNA vectors were created whose expression cassette is flanked by 28S ribosomal DNA (rDNA) sequences capable of HR into genomic rDNA. 22,23 Surprisingly, incorporation of a ribosome-skipping 2A peptide coding sequence into AAV-based targeting vectors for promoter trapping can dramatically improve gene-targeting efficiencies. 24 Blocking of undesired repair pathways such as NHEJ by inhibiting DNA-dependent protein kinase (DNA-PK) or Ku70 can also elevate HR frequency more than AAV alone. 25,26 Recently, a new AAV serotype, AAVHSC, is of remarkably higher infective and recombinant efficiency in CD34+ stem cells and some post-mitotic cells, which can enable the therapeutic manipulation of gene-based diseases. 27,28
Based on the above results, some pioneering studies addressing genetic diseases have been performed. Chamberlain et al. used AAV contained an IRES-neo cassette to disrupt dominant mutant COL1A1 collagen genes in mesenchymal stem cells from osteogenesis imperfecta sufferings. 29 Mucopolysaccharidosis type VII or Sly syndrome is an inherited disease caused by the mutation of GusB gene, and this was corrected at a frequency of up to 10–4 through AAV-mediated HR. 29 Also, a hereditary tyrosinemia type I mouse model (Fah5981SB ) was used to evaluate gene repair by delivering an AAV containing the Fah sequence (AAV-Fah5981SB ), and a correction frequency of 10–3 was achieved. 30 Epidermolysis bullosa (EB) is an inherited skin disease that is linked to dominantly inherited mutations in KRT5 or KRT14. Petek et al. used an AAV-mediated gene-targeting approach to correct KRT14 mutations in epidermal stem cells. 31 In addition, Li et al. introduced a TKNEO cassette into the APP gene in trisomy 21 induced pluripotent stem cells through AAV-mediated HR and removed the extra copy of chromosome 21. 32 Moreover, an AAV containing the gene for human coagulation factor IX (hF9) was used to infect mice with type B hemophilia, achieving on-target integration into ∼0.5% of the albumin alleles in hepatocytes and displaying therapeutic effects. 33 The outcomes described in these studies undoubtedly push AAV-mediated HR clearly into the clinical research arena, despite the ongoing challenges of HR efficiency.
AAV-Mediated Genome editing with Nucleases
To enhance the frequency of AAV-mediated HR, nucleases that can trigger DSB can be combined with the AAV to enhance HR activity. Previous studies confirmed that DSB played an important stimulatory role in genome editing and could increase AAV-mediated gene-targeting frequencies drastically by many thousand-fold. Thus, AAV-mediated genome editing with additional nucleases has developed into an efficient genetic tool, which enables the generation disease models and gene therapy agents.
So far, site-specific nucleases, including ZFN, TALEN, and CRISPR-Cas9, have led to breakthroughs in genome editing. CRISPR-Cas9 is the most popular of these because of its simplicity, efficiency, and low cost. Site-specific genome editing allows target genes to be knocked in, knocked out, or knocked down, and mutations can be restored effectively through AAV-mediated delivery.
Previous studies have demonstrated that systemic co-delivery of ZFN (or AAV-ZFN, ZFN mRNA) and AAV donor template works effectively in many situations. For reporter system construction, ZFN mRNA and AAV6 donor template were delivered into human hematopoietic stem cells (HSCs) and progenitor cells, resulting in the site-specific insertion of a promoter-GFP cassette.
34
In the preclinical gene therapy setting, ZFN and a G6PC donor transgene, both delivered with AAV, were targeted to the ROSA26 safe-harbor locus, markedly improving the survival of G6Pase knockout (G6Pase-KO) mice.
35
In a mouse model of hemophilia B, injection of an AAV encoding ZFN (AAV-ZFN) targeting the human F9 gene and AAV donor template (AAV-hF9) led to in situ correction of a defective hF9 gene and measurable circulating factor IX levels.
36,37
Moreover, AAV-mediated delivery of ZFNs individually targeted to hepatitis B virus (HBV) genes has an impact on HBV DNA replication and reduces the production of infectious HBV virions, extending this approach to new strategies to resolve infectious diseases.
38
Hitherto, ZFNs packed within AAV2/6 were applied in clinical trials for hemophilia B (NCT02695160), mucopolysaccharidosis I (NCT02702115), and mucopolysaccharidosis II (NCT03041324;
Although ZFN technology is the only genome-editing tool applied in the clinical gene therapy arena so far, off-target effects are measurable in preclinical studies, and this may be a concern. 39,40 TALEN, the second-generation genome-editing nuclease, is of lower toxicity than ZFN. 41 Chapdelaine et al. developed an AAV9 coding for one TALE coupled with a transcription activation domain to increase gene expression. 42 However, the large size (∼6.0 kb) and complex design of this construct render it comparatively difficult to package into one AAV, which restricts its application. In recent years, CRISPR-Cas9 has emerged as a more promising genome-editing tool with higher efficiency, and has been harnessed for genome editing in various species from microorganisms to plants and mammals. CRISPR-Cas9 has been applied in various fields, including disease model generation, dissection of disease mechanisms, and the development of preclinical or clinical therapies. Delivery to the appropriate target sites is essential for the practical application of this approach. However, traditional plasmid transfection is not effective in many cell types. In order to deliver a therapeutic gene editing construct to a wide variety cells in vivo or in vitro, AAV-based CRISPR-Cas9 delivery has been developed. 43 This takes advantage of the low immunogenicity of AAV, and since the CRISPR-Cas9 system is composed of two segments, Cas9 and sgRNA, delivery can be flexible according to different requirements.
Using Streptococcus pyogenes Cas9 (SpCas9), Swiech et al. designed a dual-vector system that packages SpCas9 (AAV-SpCas9) and sgRNA expression cassettes (AAV-SpGuide) in two separate viral vectors. They showed conspicuous disruption of multiple genes in post-mitotic neurons in adult mice. 44 Nishiyama et al. achieved highly efficient HR-based genome editing in developing and adult brains and in various brain areas of mouse in vivo through AAV-SpCas9 and AAV-donor-sgRNA. 45 Nevertheless, the size of the SpCas9 coding sequence (∼4.1 kb) limits its basic and clinical application as a result of the limited transgene capacity of AAV (∼4.7 kb). 46 Therefore, the subsequent discovery of the smaller-sized Staphylococcus aureus Cas 9 (SaCas9, ∼3.2 kb) has allowed its application in genome engineering. By packaging SaCas9 and its sgRNA expression cassette into a single AAV, a ∼95% decrease in serum proprotein convertase subtilisin/kexin type 9 (Pcsk9) and a ∼40% decrease in total cholesterol was observed 1 week after administration in mice. 47 Friedland et al. made use of two sgRNAs and SaCas9 packaging in a single AAV, which opened the door to multiple gene editing approaches using “all-in-one” AAV vectors. 48 In addition to AAV-CRISPR-Cas9-mediated knockout, knockdown and knock-in can also be realized by CRISPR-Cas9 and AAV donor. NRL, the gene for a key transcription factor in rod photoreceptor cells was knocked in by CRISPR-Cas9 and an AAV donor template in human embryonic stem cells (hESCs). 49 This established a platform for human genome manipulation in hESCs. Also, a large transgene cassette (∼6.5 kb) can be integrated into the genome by consecutive HR events using two separate AAV donors and CRISPR-Cas9. 50 To enhance the efficiency of CRISPR-Cas9-mediated gene disruption or HR, Gwiazda et al. utilized mRNA expressing Cas9 and mutant adenoviral E4orf6 and E1b55k helper proteins in association with AAV vectors expressing guide RNAs and recombination templates. This was named an mRNA/AAV delivery system, and worked effectively in primary human T cells. 51 Notably, homology-independent targeted integration (HITI) through AAV-SpCas9 and AAV-HITI (donor) is a new strategy for foreign gene knock-in. Using this approach, the retinal degeneration condition retinitis pigmentosa was treated in a rat model with a higher efficiency than traditional HR of 30–50%. 52 Furthermore, an AAV-SpCas9 and AAV-donor-sgRNA strategy was devised to perform homology-mediated end joining (HMEJ)-based targeted gene integration, with an efficiency much greater than strategies based on HR, NHEJ, and microhomology-mediated end joining. 53 Both the HITI and HMEJ methods function efficiently in dividing and nondividing cells in vitro and in vivo.
With the favorable progress in AAV-mediated CRISPR-Cas9 genome editing, preclinical studies of various diseases have become increasingly common. In a DMD mouse model, a frameshift mutation in Dmd exon 23 was targeted through AAV-SpCas9 coupled with guide RNAs packed in one AAV, which led to the excision of exon 23 and restored the normal dystrophin reading frame. 54 –56 Therefore, by using a similar approach, Dmd exons 52 and 53 in dystrophic mdx4cv mice were targeted, which made a new reading frame by deleting two exons and improved muscle function. 57 The smallest Cas9 ortholog, Campylobacter jejuni Cas9 (CjCas9, ∼3.0 kb) carrying sgRNA targeted to exon 23 was packed into a single AAV9 vector to treat a Dmd knockout mouse model with restoration of the reading frame through NHEJ. 58 Through two independent AAV9-SaCas9-sgRNAs targeting Lama2 in a mouse model of congenital muscular dystrophy type 1A (MDC1A), an intronic region containing the mutation was excised and a functional donor splice site was created through NHEJ. 59 Moreover, by utilizing the dual AAV system (AAV-Cas9, AAV-[donor]-sgRNA), an OTC gene mutation was corrected in newborn mice with hyperammonemia, a metabolic liver disease, at a frequency of approximately 10% 60 ; huntingtin (HTT) gene expression was reduced effectively in fibroblast cell lines derived from Huntington's disease patients 61 ; an intronic fragment of the Cep290 gene related to LCA10 in the mouse retina was deleted 62 ; the Nrl gene in rods in three different mouse models of retinal degeneration was disrupted 63 ; the genomic VEGFR2 locus related to angiogenesis-associated diseases was edited to abrogate angiogenesis in mouse models of neovascularization 64 ; the mutant amyloid precursor protein (APP) gene causes dominantly inherited Alzheimer's disease was targeted to decrease amyloid-β (Aβ) levels after the injection to hippocampus of adult mice 65 ; human-derived alpha-1 antitrypsin mutations (PiZ) were corrected by AAV-Cas9 and AAV-donor-sgRNA in a PiZ mouse model 66 ; GUCY2D mutations of LCA were disrupted in mouse and macaque photoreceptor without Cas9-specific T-cell responses, demonstrating the viability of treating inherited retinal diseases. 67 The achievements outlined above manifested appreciable therapeutic benefits relevant to human genetic disease, and reveal that the dual AAV system (AAV-Cas9, AAV-[donor]-sgRNA) is a practicably applicable method to mediate genome editing. Due to the off-target effects of CRISPR-Cas9, Cas9 ribonucleoprotein (RNP) and rAAV6 (donor) were combined to achieve HR at the HBB gene in HSCs at a frequency of 22.5%, with lower off-target effects. 68 AAV-mediated genome editing with CRISPR-Cas9 can also be applied to non-inherited diseases as well as inherited diseases using an “all-in-one” AAV. For instance, CjCas9 targeted Vegfa was to treat AMD in the mouse retina by injecting AAV-CjCas9-sgRNA in one AAV. 69 By using SaCas9 and multiple sgRNAs in one AAV, the HIV-1 provirus was excised in three different animal models, providing a potential approach for AIDS therapy. 70 To inhibit HBV replication, AAV8-SaCas9-sgRNA was used to decrease the levels of serum HBsAg and HBeAg in HBV transgenic mice. 71 Notably, the small effective capacity of AAV is a huge obstacle that limits the application of some genome-editing tools (Table 3).
Naturally occurring genome-editing nucleases
ORF, open reading frame; PAM, protospacer adjacent motif.
AAV-mediated genome editing with nucleases, especially CRISPR-Cas9, has led to rapid developments in the gene therapy field. However, how efficiency could be enhanced up to the standards needed for effective clinical treatment in terms of both AAV delivery and nuclease activity should be addressed. Furthermore, the off-target effects induced by nucleases are challenging. With the identification of high-fidelity Cas9 variants and base-editing enzymes such as cytidine deaminases or adenine deaminase, off-target effects may be minimized. 72,73
Conclusions and Outlooks
AAV is considered a promising gene delivery vector for genome editing and is widely used in a variety of studies. Compared to other viral and non-viral vectors, AAV is safe and efficient for human gene therapy applications. There are two major approaches to AAV-mediated genome editing: HR and combination with nucleases. HR is accurate, while its efficiency is pretty low. However, by combining the two methods, further efficient genome editing can be carried out with the donor template and nucleases to repair mutations. For certain diseases caused by overexpressed genes, only nucleases would be needed for disruption. Moreover, with the engineering of nucleases, base editing (C to T or A to G) has now been realized, and these types of alterations contribute a lot to single-base mutation diseases. 72,73 AAV-mediated base editing could be realized and single-base mutations could be corrected with the modulation of its package size.
Owing to the variety of AAV serotypes, AAV vectors have been used to infect various tissues successfully, including the retina, liver, heart, skeletal muscle, central nervous system, and so on through different modes of injection. As a result, rAAV is commonly used in gene therapy. Although many preclinical studies and clinical trials of AAV-mediated gene therapy have been performed, problems and obstacles (capacity, efficiency, presence of neutralizing antibodies, and off-target effects) still exist. Notably, the risk of hepatocellular carcinoma of AAV2 is reported, possibly due to the presence of a liver-specific enhancer-promoter element in the wild-type AAV2 genome, 74 which raises more concerns for the selection of the promoter in the constructs.
The capacity of AAV vector is up to 4.7 kb with workable efficiency,
46
which limits the application of some genome-editing tools. In order to enlarge the capacity of AAV vector, a set of efficient trans-splicing vectors for the transfection of large therapeutic genes were generated.
75
An intein-split system for the generation of the full-length protein after protein trans-splicing was applied into AAV base editors (AAV8.N-int-BE3 and
The optimization of AAV vectors described above enables potential and extensive applications in gene therapy (Table 4). More importantly, with the combination of nucleases, AAV-mediated genome editing has shown increasing promise in basic and clinical applications. Unlike AAV carrying therapeutic genes present as episomes, AAV-mediated genome editing with nucleases can realize in situ genetic correction, even at the resolution of single base correction. Although the presence of neutralizing antibodies to AAV vectors and nucleases is an existing problem that is yet to be solved 82 and elimination of off-target effects is also needed as much as possible, AAV vectors and nucleases can be fully exploited for in vivo and in situ gene correction for the clinical treatment of human diseases.
Problems and possible strategies of AAV-mediated genome editing
sgRNA, single guide RNA; rAAV, recombinant AAV; NHEJ, non-homologous end joining; RNP, ribonucleoprotein.
Footnotes
Acknowledgments
This work was supported by grants from the Natural Science Foundation of China (81201181; F.G.), Science Technology project of Zhejiang Province (2017C37176; F.G.).
Author Disclosure
All authors declare that they have no competing interests.