
Abstract
Microvascular dysfunction and resulting tissue hypoxia is a major contributor to the pathogenesis and evolution of cardiovascular diseases (CVD). Diverse gene and cell therapies have been proposed to preserve the microvasculature or boost angiogenesis in CVD, with moderate benefit. This study tested in vivo the impact of sequential delivery by bone-marrow (BM) cells of the pro-angiogenic factors vascular endothelial growth factor (VEGFA) and sphingosine-1-phosphate (S1P) in a myocardial infarction model. For that, mouse BM cells were transduced with lentiviral vectors coding for VEGFA or sphingosine kinase (SPHK1), which catalyzes S1P production, and injected them intravenously 4 and 7 days after cardiac ischemia–reperfusion in mice. Sequential delivery by transduced BM cells of VEGFA and S1P led to increased endothelial cell numbers and shorter extravascular distances in the infarct zone, which support better oxygen diffusion 28 days post myocardial infarction, as shown by automated 3D image analysis of the microvasculature. Milder effects were observed in the remote zone, together with increased proportion of capillaries. BM cells delivering VEGFA and S1P also decreased myofibroblast abundance and restricted adverse cardiac remodeling without major impact on cardiac contractility. The results indicate that BM cells engineered to deliver VEGFA/S1P angiogenic factors sequentially may constitute a promising strategy to improve micro-vascularization and oxygen diffusion, thus limiting the adverse consequences of cardiac ischemia.
Introduction
Cardiovascular disease (CVD) has been the leading cause of death in the United States since the beginning of the 20th century, with almost half of those deaths caused by coronary heart disease. 1,2 The most common long-term consequence of coronary heart disease is heart failure (HF), and its prevalence increased between 2009 and 2014, in part due to the improvement in acute treatment of CVD. 2 One of the clinical manifestations of coronary heart disease is myocardial infarction (MI), which is the sudden death of myocardial tissue from ischemia caused by a blockage of a coronary artery, usually from the rupturing of an atherosclerotic plaque. 3 MI causes cardiomyocyte death, which triggers acute inflammation, angiogenesis, and finally scar formation. 4 Fibrotic scar formation is the main healing mechanism after MI due to the limited ability of the adult mammalian heart to regenerate. 3 Fibrosis causes abnormalities in both relaxation and contractility of the heart muscle, which causes ventricular remodeling, a mechanism of heart adaptation to adverse stimuli. 5 Remodeling includes left ventricular wall thinning, ventricle dilation, and changes in its shape from ellipsoid to more spherical. 6 At the cellular level, cardiomyocytes in the viable myocardium undergo hypertrophy triggered by increased load. These malformations create increasing wall stress, which causes higher myocardial oxygen demand and could cause further enlargement of the area at risk for ischemia. 7 Pathological heart remodeling caused by MI and secondary hypoxia induces HF and high mortality in the long term. 8 Indeed, HF becomes the main cause of death of patients >65 years old within 6 years after MI. 9 HF can also derive from other CVD involving microvascular dysfunction and tissue hypoxia as hypertensive cardiac hypertrophy and diabetic coronary microangiopathy. 10 For that reason, prevention and ameliorating effects of HF are of great importance.
Over the past decades, a need for an effective therapy to prevent and limit hypoxia-related cardiac adverse remodeling and as a consequence HF, particularly after MI, remains unmet. Since the introduction of fibrinolysis and angioplasty in the late 1970s, acute treatment of MI has improved dramatically, although the mortality rate caused by CVD remains high. 11 Trying to overcome the inability of adult cardiac muscle to regenerate, many therapies were implemented, including cell and gene therapies and delivery of cytokines and soluble factors. One of the aims of these therapies has been modulating angiogenesis, 12 –18 since by preserving the microvasculature or boosting the formation of new vessels, tissue oxygenation would improve, alleviating adverse consequences of acute or chronic ischemia. In recent years, importance has also been given to modulating immune response after MI for its beneficial effect on heart repair. 19 –22
One of the main unsolved questions in these studies was the optimal delivery route for the factors with strategies including delivery of the recombinant protein, 23,24 gene delivery using plasmids 16,17 and modified RNA, 25 and virus-based approaches, mostly using adeno-associated virus (AAV). 14,26,27 On the other hand, cell therapies, whose main effect is believed to be based on secreting factors such as vascular endothelial growth factor A (VEGFA) and PDGF showed limited long-term improvement in cardiac vascularization. 28 –30 It was hypothesized that combining two approaches by delivering cells but also transducing them to overexpress proangiogenic factors may bring an improvement in cardiac vascularization after ischemia. In the field of proangiogenic therapies, the strong proangiogenic factor VEGFA generated promising results in animal models but showed no sustained improvement in myocardial perfusion measured in the NORTHERN clinical trial. 13 However, co-delivery of VEGFA and sphingosine-1-phosphate (S1P) by AAV promoted angiogenesis and reduced cell apoptosis in a pig model of MI. 14 Of note, sequential instead of simultaneous exposure to VEGFA and S1P led to the formation of more stable vessels in scaffolds in vitro, but this approach has never been tested in vivo. 31
This study investigated the effect of bone-marrow (BM) cells engineered to produce VEGFA or S1P and injected sequentially in improving vascularization in a preclinical mouse model of acute ischemic heart disease. Indeed, it is demonstrated that lentiviral vector (LV)-transduced BM cells injected intravenously (i.v.) in mice to deliver the pro-angiogenic factors VEGFA and S1P sequentially resulted in sustained increased vascularization 28 days after MI assessed by a newly implemented pipeline for automatized 3D image analysis of the microvasculature 32 and in contrast to the transient effect reported for non-transduced BM cells. As a consequence, the strategy also limited fibrosis and adverse cardiac remodeling. This study may offer new therapeutic possibilities for patients suffering cardiac ischemic disease.
Methods
All experimental procedures are explained in detail in the Supplementary Methods.
LV construction and evaluation
Human VEGFA and SPHK1 and GFP cDNAs were amplified by polymerase chain reaction and inserted downstream to the SFFV promoter of the
Flow cytometry
BM cells were stained with antibodies against CD11b-AF647, F4/80-PE-Cy7, Ly6C-FITC, CCR2-PE, and CX3CR1-Pacific Blue and 7-AAD live/death dye. Sample acquisition was performed using a BD LSRFortessa flow cytometer (BD Biosciences) and analyzed using FlowJo software (FlowJo LLC). Detailed staining procedure and a list of antibodies is available in the Supplementary Methods.
Aortic ring assay
Aortic ring assay was performed, as described in Baker et al. 34 Mouse aortic rings were cultured in the presence of various combinations of soluble factors (rhVEGFA 30 ng/mL, S1P 1 μM) or mouse BM cells previously transduced with LVs (BMhVEGF, BMhSPHK1). On day 8, rings were fixed in 1.6% paraformaldehyde, and incubated overnight at 4°C with the following antibodies/reagents: IB4 or anti-CD31 (1:150) for endothelial cells and anti-smooth-muscle actin (SMA; 1:400) for mural cells. Images were acquired with a Nikon A1R confocal system coupled to a Nikon Ti-Eclipse microscope using a 10 × objective and capturing 400 μm in depth with a z-stack every 3 μm. Analysis of aortic ring images was performed using Imaris software and a Matlab-based method for 3D-microvasculature developed in the lab. Reagents, antibodies, and detailed procedures are described in the Supplementary Methods.
In vivo mouse ischemia–reperfusion experiment protocol
Mice were kept at the animal facility of the Centro Nacional de Investigaciones Cardiovasculares under specific pathogen-free conditions and in accordance with the institutional guidelines. Animal procedures were approved by the corresponding legal authority of the local government of Madrid (reference number of approval PROEX/ 34/13). Ischemia–reperfusion (I/R) was performed in 10- to 12-week-old male BL6 mice by ligation of the left anterior descendent coronary artery for 45 min of ischemia followed by reperfusion. Echocardiography was performed 3 days after to confirm MI, and mice with a left ventricular ejection fraction (LVEF) between 20% and 40% were randomly assigned to control and treated groups. Four days post I/R, mice were injected i.v. with 5 × 106 BMhVEGF in 100 μL of phosphate-buffered saline (PBS) or with PBS only as a control. Seven days post I/R, a second injection was performed with 5 × 106 BMhSPHK1 or PBS. On day 28, after endpoint echocardiography, hearts were weighed and collected. Echocardiography was performed using Vevo 2100 ultra high frequency ultrasound to assess cardiac parameters (end-diastolic and end-systolic volumes, stroke volume (SV), LVEF and left ventricular posterior wall thickness). Analysis of heart roundness (i.e., sphericity) was performed on macroscopic images of the dissected hearts using ImageJ 35 software. Fibrosis was assessed based on Masson's trichrome staining of a series of seven levels of tissue cuts, starting from the apex to the base of the heart and visualized using NanoZoomer-2.ORS® (Hamamatsu); scar area is represented as the percentage of left-ventricular area using ImageJ software. To quantify cardiomyocyte size, an ImageJ-based semi-automated macro was designed where cardiomyocytes were recognized by laminin staining and their area and perimeter quantified. Detailed experimental procedures are described in the Supplementary Methods.
Microvasculature analysis
Sections (15 μm thick) were stained with anti-CD31, anti-PDGFRβ, and anti-SMA antibodies and with Hoechst for nuclear staining. Images were acquired with a Leica SP5 confocal microscope using a 40 × objective (NA 1.25) with oil immersion with z-stacks captured every 1 μm. Analysis of the microvasculature was based on the fully automatic image analysis pipeline developed in the authors' laboratory to analyze microvascular data. The pipeline permits the calculation of a pool of parameters that quantify all major features of the vascular network. It has been described in detail in Gkontra et al., 32 while a brief overview of necessary adaptations is provided in the Supplementary Methods.
Statistics
Statistical analysis was performed using GraphPad Prism v6/7. Distribution of the data sets was checked with D'Agostino–Pearson and Shapiro–Wilk normality tests, and then parametric or nonparametric tests were performed as indicated in the legends. Outliers were identified using the ROUT method (Q = 1.000%).
Results
Experimental design for in vivo sequential VEGF/S1P gene–cell angiotherapy in a MI mouse model
Sequential delivery of VEGFA (VEGF hereafter) and S1P has been shown to increase the formation of mature vessels in in vitro scaffolds compared to the effect of single or combined factors. 31 It was confirmed in the mouse aortic ring assay that sequential delivery of S1P after VEGF or combined delivery of the two factors induced a significantly higher proportion of sprouts, covered by SMA+ cells and a more complex vascular network than VEGF alone (Supplementary Fig. S1A and B). A decision was therefore made to test the effects of a combined gene–cell therapy in which BM cells were boosted to produce VEGF and S1P by LV transduction and to deliver these factors sequentially. For that, LVs were generated encoding for the human VEGF, or SPHK1 (sphingosine-kinase 1), the enzyme that catalyzes the conversion of the precursor sphingolipid sphingosine to the active form S1P (herein LV_ hVEGF and LV_hSPHK1, respectively). Then, the efficacy of the LV was validated in HEK and mouse BM cells transduced with LV_hVEGF and LV_hSPHK1, which indeed overexpressed VEGF and produced S1P (Supplementary Fig. S2). The proposed strategy was tested in the mouse model of acute cardiac I/R, which is closer to the human MI clinical practice. Before proceeding with the in vivo approach, the ability of i.v. injected BM cells to migrate to the infarcted myocardium was confirmed. Histological analysis performed 24 h after Tomato+ BM cell i.v. injection confirmed the presence of red round cells in the infarct zone of hearts 4 days after I/R. Only an unspecific non-membrane-associated red signal was detected in the hearts from PBS-injected mice (Supplementary Fig. S3A).
Prior to their i.v. injection, the study also analyzed whether LV transduction and culture in the presence of stem-cell factor (SCF) and macrophage colony-stimulating factor in vitro impacted on the phenotype of BM cells. Following the flow cytometry gating strategy detailed in the Methods (Supplementary Fig. S3B and C), it was observed that after LV transduction with LV_hVEGF or LV_hSPHK1, the amount of neutrophils increased, macrophages decreased, and the scarce myeloid progenitors remained unchanged (Supplementary Fig. S3D–F). Notably, although no differences in the percentage of monocytes (∼60%) were observed, after LV transduction, there was an increase in their CX3CR1 cell surface expression, particularly after LV_hSPHK1 transduction with milder changes in CCR2 levels (Supplementary Fig. S3G).
BM cell-mediated delivery of VEGF and S1P improves vascularization and reduces inter-capillary distances in the post-infarcted heart
Next, the study tested the in vivo impact of boosting BM cell sequential secretion of VEGF and S1P in a mouse model of cardiac I/R (Supplementary Fig. S4A). MI was confirmed by echocardiography 3 days post I/R, and mice with a LVEF of 20–40% were included in the study. Mice were afterwards randomly assigned to control and treated groups and then injected i.v. on day 4 post I/R with PBS or with 5 × 106 of BMhVEGF followed on day 7 by PBS or 5 × 106 of BMhSPHK1 similar to the time points tested in the in vitro aortic ring assay (Supplementary Fig. S1).
Sequential VEGF/S1P angiotherapy by BM cells was primarily designed to enhance angiogenesis and increase the functional microvasculature after I/R, which may secondarily reduce post-MI adverse effects. In an effort to quantify the effect of the proposed treatment to the microvasculature in an unbiased and reproducible manner, images of transverse slices of cardiac tissue from treated and untreated subjects were analyzed by means of a 3D fully automated image analysis pipeline developed at the authors' lab. 32 The tissue was labeled for blood vessels, in particular for endothelial cells (CD31) and for SMA and PDGFRβ+ cells, and then imaged by confocal microscopy. The automated analysis approach allowed the post-MI changes of the microvasculature of the control and BM cell-delivered VEGF/S1P hearts to be compared in terms of all major microvascular characteristics: morphology, topology, angio-architecture, efficiency for oxygen diffusion, as well as its relation with SMA+ and PDGFRβ+ cells. A summary of all quantified parameters and their changes are provided in Table 1. Reference values for the parameters extracted from microvascular tissues of basal mice are also given.
Quantitative analysis of microvasculature parameters in the cardiac tissue from treated and non-treated mice after myocardial infarction
The mean ± standard deviation of all parameters for cardiac tissue from treated (BMhVEGF + BMhSPHK1) and non-treated (phosphate-buffered saline) mice along with pairwise comparisons among the different tissue conditions. Capillaries correspond to CD31+ and SMA– vessels of diameter <5 μm, enlarged capillaries to CD31+ and SMA– vessels of diameter ≥5 μm, and SMA+ vessels to CD31+ and SMA+ vessels regardless their diameter. Up and down arrows indicate increase and decrease, respectively, of the parameter value for (1) tissue from infarcted areas of subjects treated with BMhVEGF + BMhSPHK1 compared to tissue from infarcted areas of untreated subjects, and (2) tissue from remote areas of subjects treated with BMhVEGF + BMhSPHK1 compared to tissue from remote areas of untreated subjects.
Number per mm vascular length.
Number per mm3 vascular volume.
p < 0.05; ** p < 0.01. The p-values were calculated by means of a two-sample t-test.
BM, bone marrow; VEGF, vascular endothelial growth factor; SPHK1, sphingosine kinase; SMA, smooth-muscle actin.
Among fractal parameters, lacunarity, a parameter measuring the heterogeneity in the distribution of the size of the gaps in the tissue, significantly decreased in the infarcted area of the hearts in the treated group. This indicates a smaller dispersion of non-vascularized areas, implying a better-organized microvasculature (Table 1). Although no significant differences were observed in the overall vascular volume density 28 days after I/R, after decomposing the microvasculature into its components, that is, capillaries (CD31+SMA−, <5 μm), enlarged capillaries (CD31+SMA−, >5.1 μm), and arterioles (CD31+SMA+), a trend to increased capillary volume density and decreased enlarged capillaries was noticed in the infarcted zone of hearts from mice injected with BM cells delivering VEGF/S1P compared to controls (Fig. 1A–C and Table 1). There were, however, no changes in the volume density and the percentage of arterioles (Fig. 1B and C). Accordingly, treatment also resulted in significantly more abundant endothelial cells in the vasculature of the infarcted zone (Fig. 1A). Interestingly, in the remote zone, a significant increase in the volume and percentage of capillaries and a decrease in the percentage of enlarged capillaries were also observe in hearts from the VEGF/S1P-treated group, in spite of the lack of statistically significant between-group differences in overall vascular volume density. Concomitantly, endothelial cell abundance in the remote zone was slightly increased in the treated group (Fig. 1D–F). These findings indicate that combined delivery of VEGF/S1P impacted mostly on the capillaries, increasing their volume density and the endothelial cell content while decreasing their remodeling/enlargement in both the infarcted and the remote zones.

Sequential bone marrow (BM) cell delivery of vascular endothelial growth factor (VEGF) and sphingosine-1-phosphate (S1P) increases capillary volume density and endothelial cell number on the infarcted heart post myocardial infarction (MI).
The main function of the microvasculature is to provide nutrients and oxygen to the heart, particularly after ischemia, to preserve cell viability and promote repair. Thus, toward obtaining insights into the efficiency of the network for oxygen diffusion, traditional indicators of oxygen diffusion (i.e., maximal extravascular distances) were studied by means of the automated pipeline. According to the larger proportion of capillaries and higher endothelial cell numbers, maximal extravascular distances were significantly reduced in the infarct zone of hearts from mice injected with BM cells producing VEGF and S1P 28 days after I/R compared to the controls, indicating better oxygen diffusion. A similar trend to reduced extravascular distances was observed in the remote zone of treated mice (Fig. 2).
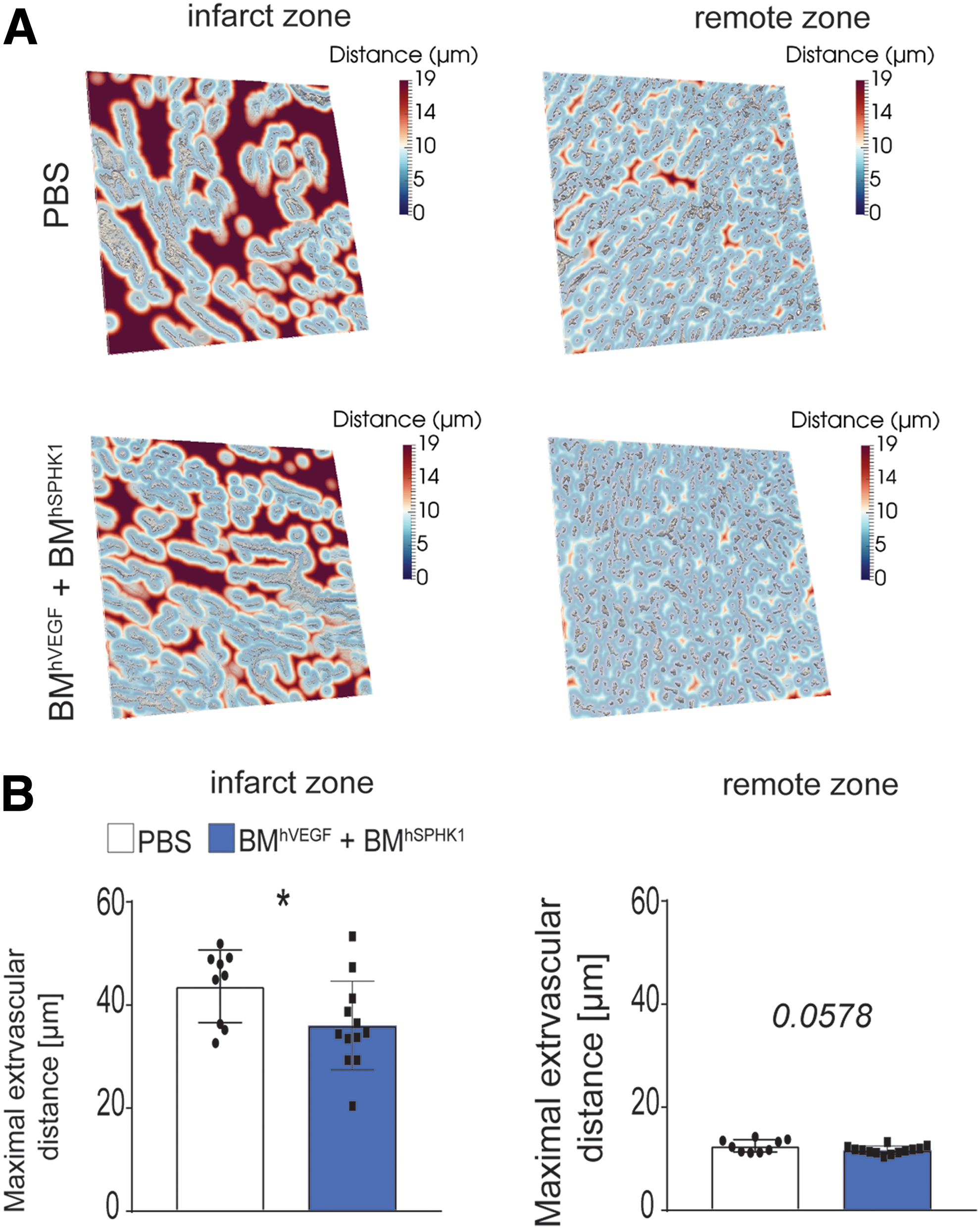
BM cell sequential delivery of VEGF and S1P reduces intercapillary distances in the post-infarcted heart.
Next, the study sought to decipher the contribution of VEGF and S1P overexpression versus endogenous secretion of factors by BM cells to the myocardial angiogenic phenotype, since PBS was used as in vivo control. Non-transduced BM cells did not increase vascular sprouting versus control medium in the aortic ring in contrast to sequential exposure to BM cells secreting VEGF and S1P, which induced a significant increase in vascular sprouts with a good proportion covered by SMA+ cells, indicative of stable vessels, and a more complex vascular network (Supplementary Fig. S4B). These findings argue in favor of overexpressed VEGF and S1P by LV-transduced BM cells as the factors responsible for boosting capillarization post MI.
Sequential BM cell delivery of VEGF and S1P diminishes post-infarction tissue fibrosis with no impact on cardiac contractility
After MI, dead cardiomyocytes are mostly replaced by fibrotic tissue, reducing cardiac pump function. The study analyzed whether improved capillarization and oxygen diffusion by BM cell sequential delivery of VEGF/S1P had any impact on cardiomyocyte survival and thus in cardiac performance. Despite the presence of isolated clusters of viable cardiomyocytes within the scar of infarcted hearts from VEGF/S1P-treated mice but not in untreated controls 28 days post-I/R (Supplementary Fig. S5A), echocardiography analysis showed no significant between-group differences in the reduced LVEF and the SV (Supplementary Fig. S5B and C).
Regardless the ejection fraction values, fibrosis extent and tissue stiffness are the major factors driving post-MI progression to HF. Therefore, although a major impact of the BM cell-VEGF/S1P therapy could not be observed on cardiac contractility, next fibrosis-related parameters were assessed, since cardiac injury and hypoxia are enhancers of the myofibroblast-driven fibrotic program. 36,37 Between-group differences could not be detected in the total scar volume of the left ventricular free wall calculated from serial Masson-stained sections spanning the whole heart 28 days post I/R (Fig. 3A and B). However, the cardiac section closest to the valves consistently showed a significantly reduced scar area in the BM cell VEFG/S1P-treated group (Fig. 3C). Complementarily, using the images of mouse cardiac tissues stained for PDGFRβ and CD31, the myofibroblast abundance were also automatically quantitated as the ratio of the volume of myofibroblasts over the volume of all PDGFRβ+ cells. In this approach, myofibroblasts are considered PDGFRβ+ areas that are not in touch with the vessels. 32,38 Following this approach, significantly reduced myofibroblast abundance was noticed in the infarct zone of hearts from BM cell VEGF/S1P-treated mice compared to untreated controls. Myofibroblasts were barely detected, but there were fewer in the remote zone of the treated group (Fig. 3D and E and Table 1). Since myofibroblasts are in charge of secreting and remodeling extracellular matrix proteins, particularly collagens, to form the scar, myofibroblast reduction prompted collagen fiber organization and condensation to be analyzed, in spite of no significant differences observed in overall scar area. Analysis of first-order features of collagen fibrils in second harmonic generation microscopy images revealed increased skewness and kurtosis in the collagen fibers of the scar from infarcted hearts of the BM cell VEGF/S1P-treated group compared to controls (Fig. 3F and G). These values would indicate thinner and underdeveloped collagen fibers and thus lower tissue stiffness. 39

Sequential BM cell-secreted VEGF and S1P angiotherapy reduces myofibroblast abundance and collagen fiber compaction in the infarcted heart post I/R.
VEGF and S1P sequential delivery by BM cells limits adverse cardiac remodeling post MI
One of the consequences of MI is cardiac remodeling defined as the post-damage alterations that result in acute and chronic changes of heart size, mass, geometry, and function, which may finally lead to cardiac dysfunction and HF. 40 First, ventricle cavity size and wall thickness were quantitated, and a trend was observed of reduced dilatation and thickening of the left ventricle in the VEGF/S1P-treated mice 28 days post I/R (Fig. 4A and B). Since volume and thickness changes are not fully developed 4 weeks after I/R in mice, 41 global cardiac geometry was also analyzed, which is modified more acutely. 40 It was found that hearts from BM cell VEGF/S1P-treated mice kept their ellipsoid-like shape in contrast to those from untreated control mice that became significantly rounder at 28 days post I/R, as expected (Fig. 4C). Moreover, analysis of cardiomyocytes in the remote zone, which usually become hypertrophic in response to post-MI left ventricular stiffness and volume overload, 40 showed that cardiomyocytes in the remote zone of hearts from BM cell VEGF/S1P-treated mice became less hypertrophic, with significantly smaller area and perimeter compared to control mice 28 days post I/R (Fig. 4D and E). Finally, cardiac remodeling also results in changes at the molecular level, with proteins increasing (collagen type I) or decreasing (Atpa2a/Serca2a) during the process. 40 Notably, VEGF/S1P treatment led to decreased collagen type I and increased Atp2a2 (Serca2) protein levels in heart extracts compared to untreated mice 28 days post I/R (Fig. 4F and G), confirming that the angiotherapy ameliorated adverse cardiac remodeling. 40
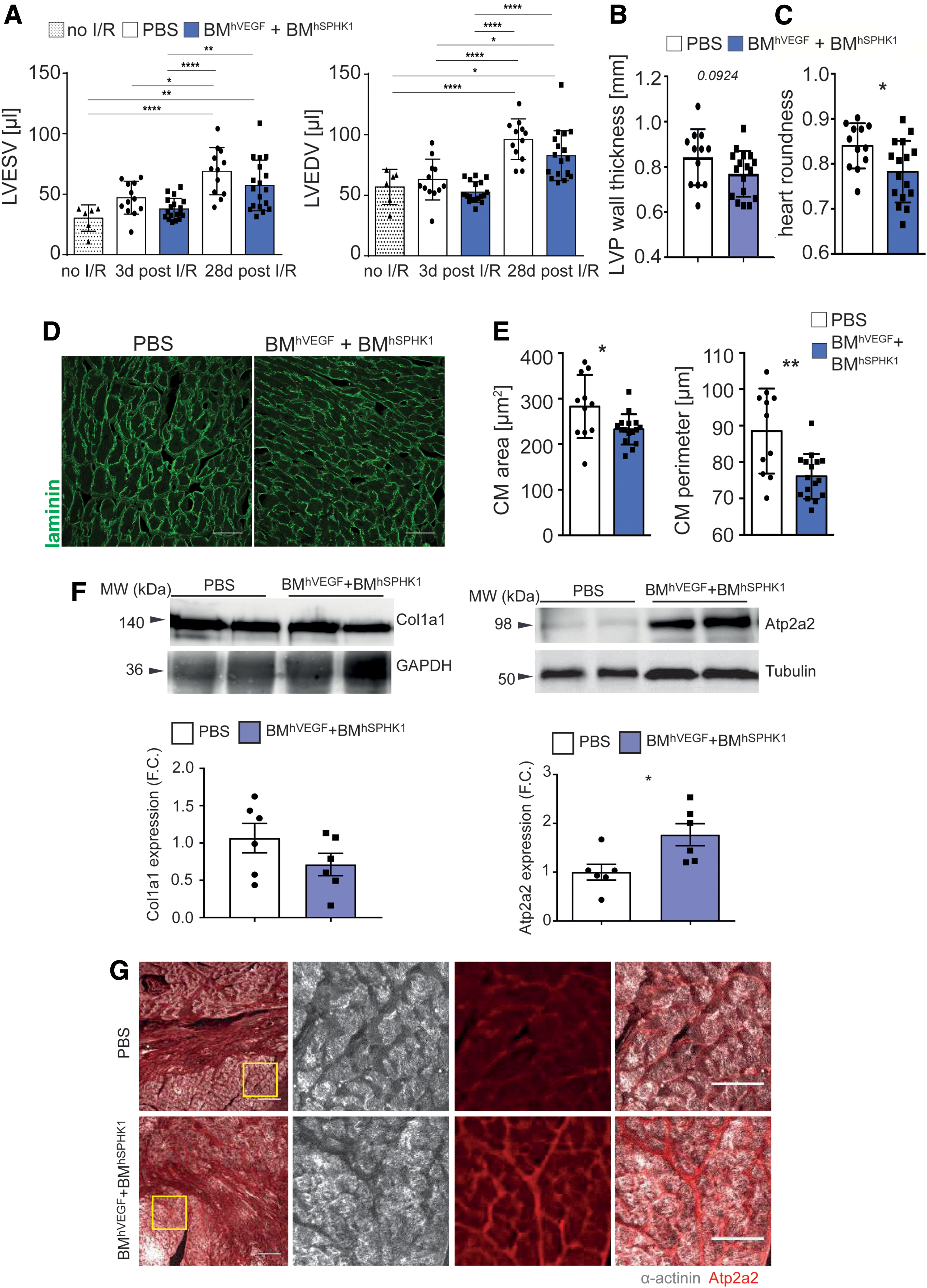
Sequential delivery of VEGF and S1P by lentiviral vector (LV)-transduced BM cells limits adverse cardiac remodeling post MI.
Collectively, these findings indicate that sequential VEGF/S1P gene–BM cell therapy limits post-MI adverse cardiac remodeling consistent with the enhanced capillarization and oxygen diffusion effects and reduced myofibroblast abundance in the infarcted tissue.
Discussion
This study tested the effect of sequential delivery by BM cells of the proangiogenic factors VEGF and S1P on boosting angiogenesis and/or preserving the microvasculature after MI in vivo. The intervention succeeded in increasing endothelial cell numbers and capillaries and diminishing adverse angioadaptation, thus improving oxygen diffusion 4 weeks after the ischemic event. In spite of no major effect on cardiac contractility, BM cell sequential VEGF/S1P therapy also reduced myofibroblast-driven tissue fibrosis and limited adverse post-MI cardiac remodeling.
In terms of safety and possible translation of this therapy, LV transduction is performed ex vivo, and the injected cells are mostly differentiated. This would limit the possibility of transferring the LV to other cell types in the organism and reduce risks of genetic alterations in the hematopoietic progenitors and their progeny by LV insertion. Previous analysis in the pig infarcted heart showed that on day 3 post MI, the 3D structure of the microvasculature is mostly preserved, but its function starts to decline, as manifested by the larger extravascular distances causing disturbed oxygen diffusion. 32 Endogenous angiogenesis after MI occurs during the first 2 weeks after the ischemic event. Thus, detailed kinetics of the endothelial cell marker PECAM1/CD31 in a mouse model of permanent coronary artery ligation showed an increase in endothelial cells at 2 days with a maximum at 4 days and then a decline and return to baseline levels 7 days after MI. 4,36,42,43 These findings support the vascular boosting intervention 4 days post MI.
This study analyzed the impact of the BM cell VEGF/S1P therapy at the 28-day endpoint post I/R. Notably, persistent benefits were observed from this strategy in contrast to previously reported therapeutic approaches in which effects on the vasculature were often transient. For example, the intramyocardial injection of mesenchymal stem cells showed benefits in perfusion on the infarcted area at the first week but not after 4 weeks. 44 This newly implemented BM cell VEGF/S1P therapy mainly induced an increase in the abundance of endothelial cells and reduced extravascular distances in the infarcted area of hearts from treated mice. Concomitantly, trends toward decreased presence of enlarged capillaries were observed. These effects might indicate reduced angioadaptation, which normally occurs in response to capillary loss and flow redistribution in the remaining vessels. 32,45 Previous proangiogenic therapies, including AAV-mediated overexpression of VEGF and ANG1 in a pig model of MI, 14 plasmid-mediated overexpression of bFGF and PDGF in a rat model of MI, 46 and microcapsule release of FGF2 and HGF in a rat model of HF, 47 improved the cardiac vasculature by increasing both capillary and arteriole/large-vessel content. As it is becoming recently accepted, interventions in the cardiac stroma 48 as the microvasculature may secondarily lead to reduced CM death, tissue damage, and fibrosis, since this is often triggered by tissue hypoxia. 37
The intervention was based on previous reports showing that sequential VEGF and S1P actions were of benefit to induce stable vessels in scaffolds in vitro. 31 VEGF is a proangiogenic factor, which induces endothelial cell migration, proliferation, and survival, depending on the endothelial cell context, and at high persistent levels may induce vascular leakiness and regression. 49,50 The increase in endothelial cell number in the infarcted area of hearts from treated mice suggests an overall effect of VEGF in endothelial cell proliferation and/or survival. Moreover, stimulation of endothelial cells with VEGF causes overexpression of S1P1 receptors on these cells and, in isolated arteries, enhances S1P-mediated vasorelaxation and eNOS phosphorylation. 31 This effect may contribute further to pro-survival and proangiogenic programs in the capillary endothelium. It is worth mentioning that although VEGF and S1P are endogenously produced in the heart, cell-type transcriptomics analysis in the pig heart revealed decreased mRNA levels of VEGF and SPHK1 3 days post I/R (unpublished data), supporting the rationale of boosting VEGF/S1P tissue levels by LV-BM delivery. S1P itself is considered a master regulator of signaling pathways involved in cardiac improvement after MI, and it has lately attracted attention as a possible target. 51 It cannot be ruled out that S1P secreted locally by the transduced BM cells in the infarcted area could have, in addition to enhanced capillarization, some direct impact on cardiomyocyte survival and thus on reducing cardiac damage. Nonetheless, the lack of major differences in the infarct size argues against a major role of this protection mechanism. Furthermore, in contrast to the reported S1P effect on myofibroblasts, 51 reduced myofibroblast abundance is observed, suggesting that S1P paracrine effects on cell types other than endothelial cells seem unlikely. Autocrine effects, however, may still be possible, since transduction with LV_hVEGF or LV_hSPHK1 resulted in BM-derived monocytes with higher CX3CR1 levels, which may enhance their early traffic to the damaged heart. 52 In this line, higher expression of the CX3CR1 receptor on circulating Ly6Clow monocytes has been shown to induce a positive effect on their recruitment and the promotion of angiogenesis in the injury site in hind-limb ischemia 53 and carotid injury 54 models. Moreover, S1P can promote an anti-inflammatory phenotype in macrophages through activation of their S1P1 receptor after damage and contribute to macrophage-mediated cardiac repair. 55 This possibility merits further analysis with macrophage specific markers. Of note, significantly increased capillaries and a trend to diminished extravascular distances were also observed in the remote zone of BM cell VEGF/S1P-treated infarcted mice, suggesting that LV-BM cells can also induce vascular effects in that area. 56
VEGF/S1P gene–cell therapy did not improve post-MI cardiac contractility in accordance with other previously reported strategies. Nevertheless, in spite of no major impact on contractility, VEGF/S1P combined angiotherapy limited post-MI adverse cardiac remodeling, as demonstrated by the preserved cardiac geometry and cardiomyocyte morphology in the remote zone, as well as the higher Atp2a2 protein levels. The reduced cardiac sphericity induced by the angiotherapy supports its clinical relevance, since it has been related to post-MI long-term survival. 57 This improvement may be related to the reduced collagen stiffness and myofibroblast abundance in the infarcted area but also in the remote zone of hearts from the treated mice. 58 Whether remote changes are directly related to the therapy or indirectly as a consequence of the effects on the scar properties (better perfusion, lower stiffness) in the infarcted area remains to be elucidated. Moreover, since cardiac macrophages proliferate in response to mechanical stretching caused by stiffer failing myocardium, 20 the reduced collagen stiffness in treated mice may also explain the trend to lower amount of macrophages in the remote zone observed in these mice (unpublished data).
There were fewer functional beneficial effects of the angiotherapy than expected, despite a consistent biological effect on histology endpoints. This may be related to insufficient factor concentration in the in vivo experiments, and more dramatic effect needs to be induced to elicit relevant functional changes. In this regard, future studies with higher transgene overexpression, or more effective gene–cell delivery systems (intracoronary, trans-endocardial) may overcome the limitations observed in the murine model. 59
This work presents some limitations, which require future studies. First, in this study, gene–cell therapy was compared to a placebo to acquire a clear determination of its efficacy. Although the absence of angiogenic effects induced by non-transduced BM cells on the mouse aortic rings argue in favor of specific effects of sequential delivery of VEGF plus S1P on the phenotype observed in the microvasculature 28 days post MI, comparison with injection of wild-type or LV mock-transduced BM cells would help establish the precise contribution of paracrine signaling by the BM cells versus the delivered VEGF/S1P in vivo. Second, deciphering the actual impact of the combined gene–cell VEGF and S1P therapy on the microvasculature, cardiomyocyte survival, and/or monocyte/macrophage subsets will require deeper analysis at earlier time points after ischemia. Third, the benefits of this strategy have only been tested in the I/R model by its proximity to clinical practice. It will also be of interest to evaluate the impact of combined VEGF/S1P gene–cell therapy in the extensively used permanent left anterior descending coronary artery ligation in which arterialization is relevant for revascularization, as well as in CVD involving chronic cardiac ischemia. Forth, in terms of global cardiac function, clinical trials using intracoronary injection of whole BM showed no long-term improvement in LVEF after MI, 60,61 despite promising short-term effects. 62,63 Therefore, analysis of the impact of BM cell VEGF/S1P therapy at longer time points after cardiac ischemia is guaranteed to elucidate whether the beneficial effects on cardiac remodeling persist in time.
In sum, this study offers a novel therapeutic strategy based on sequential BM cell delivery of angiogenic factors to enhance micro-vascularization and oxygen diffusion, thus limiting the adverse consequences of cardiac ischemic diseases.
Footnotes
Acknowledgments
We thank Mónica Gómez for her help with I/R surgery, Lorena Flores and Ana Vanessa Alonso López for performing and analyzing echocardiography, Dr. Manuel Lobo for ultrasound image analysis, Drs. Borreguero and Maria Villalba for their help in interpreting the results, and Verónica Labrador and CNIC's Microcopy Unit for their expertise and help with image analysis.
This study was supported by grants from the Spanish Ministerio de Ciencia, Innovación y Universidades SAF2014-52050-R and SAF2017-83229-R to A.G.A. and BIO2015-67580-P to J.V. and the Carlos III Institute of Health-Fondo de Investigación Sanitaria (PRB2, IPT13/0001-ISCIII-SGEFI/FEDER, ProteoRed). The research leading to these results received funding from the People Programme (Marie Curie Action) of the European Union's Seventh Framework Programme (FP7/2007-2013) under REA grant Agreement 608027. The CNIC is supported by the Spanish Ministerio de Ciencia, Innovación y Universidades and the Pro-CNIC Foundation, and is a Severo Ochoa Center of Excellence (award SEV-2015-0505).
Author Disclosure
No competing financial interests exist.
Supplementary Material
Supplementary Methods
Supplementary Fig. S1
Supplementary Fig. S2
Supplementary Fig. S3
Supplementary Fig. S4
Supplementary Fig. S5
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.