
Abstract
Alport syndrome is a genetic disease caused by mutations in type IV collagen and is characterized by progressive kidney disease. The Col4α3 −/− mouse model recapitulates the main features of human Alport syndrome. Previously, it was reported that kidney microRNA-21 (miR-21) expression is significantly increased in Col4α3 −/− mice, and administration of anti-miR-21 oligonucleotides (anti-miR-21) attenuates kidney disease progression in Col4α3 −/− mice, indicating that miR-21 is a viable therapeutic target for Alport syndrome. However, the expression pattern of miR-21 in the kidneys of patients with human Alport syndrome has not been evaluated. Paraffin-embedded kidney specimens were obtained from 27 patients with Alport syndrome and from 10 normal controls. They were evaluated for miR-21 expression and for in situ hybridization and mRNA expression by quantitative polymerase chain reaction. In addition, anti-miR-21 was administrated to Col4α3 −/− mice at different stages of disease, and changes in proteinuria, kidney function, and survival were monitored. Transcriptomic analysis of mouse kidney was conducted using RNA sequencing. miR-21 expression was significantly elevated in kidney specimens from patients with Alport syndrome compared to normal controls. Elevated renal miR-21 expression positively correlated with 24 h urine protein, serum blood urea nitrogen, serum creatinine, and severity of kidney pathology. On histological evaluation, high levels of miR-21 were localized to damaged tubular epithelial cells and glomeruli. Kidney specimens from both humans and mice with Alport syndrome exhibited abnormal expression of genes involved in kidney injury, fibrosis, inflammation, mitochondrial function, and lipid metabolism. Administration of anti-miR-21 to Alport mice resulted in slowing of kidney function decline, partial reversal of abnormal gene expression associated with disease pathology, and improved survival. Increased levels of miR-21 in human Alport kidney samples showed a correlation with kidney disease severity measured by proteinuria, biomarkers of kidney function, and kidney histopathology scores. These human data, combined with the finding that a reduction of miR-21 in Col4α3 −/− mice improves kidney phenotype and survival, support miR-21 as a viable therapeutic target for the treatment of Alport syndrome.
Introduction
Alport syndrome is a rare inherited disorder with renal, cochlear, and ocular involvement caused by mutations in the COL4A3, COL4A4, or COL4A5 genes. The X-linked form of Alport syndrome results from mutations in COL4A5, which encodes the collagen IV α5 chain. Autosomal forms of Alport syndrome are caused by mutations in COL4A3 and/or COL4A4 encoding the collagen IV α3 and α4 chains, respectively. 1,2 Without treatment, most patients with Alport syndrome will progress to end-stage renal disease. Currently, there is no disease-modifying therapy available for patients with this disorder. Therefore, developing a therapy that effectively slows kidney disease progression in patients with Alport syndrome is of critical importance.
The impaired production and/or assembly of collagen IV results in structural defects of the glomerular basement membrane (GBM), which leads to hematuria, proteinuria, and progressive renal failure. 3 The Col4α3 knockout mouse model is a well-characterized murine model of Alport syndrome that, similar to humans with this disorder, exhibits progressive glomerulonephritis characterized by proteinuria, increase in serum blood urea nitrogen (BUN), and typical thickening and splitting of the GBM. 4 Col4α3 knockout mice have been widely utilized for both Alport syndrome and general chronic kidney disease (CKD) research. 5 –7
MicroRNAs (miRNAs) are small noncoding RNAs, approximately 20–25 nucleotides in length, which can regulate gene expression by inhibiting the translation or increasing the degradation of their target messenger RNAs. 8 miRNAs play important roles in multiple biological processes, including immune modulation, metabolic control, neuronal development, cell cycle, muscle differentiation, and stem-cell differentiation. Dysregulation of miRNAs has been observed in many diseases such as cancer, hepatitis C infection, inflammatory diseases, myocardial infarction, metabolic disease, and kidney diseases. 8 The ability of a single miRNA to regulate multiple downstream target mRNAs altered in disease conditions makes miRNAs attractive therapeutic targets with the potential to impact a variety of molecular pathways. 9 Among the previously characterized miRNAs, miR-21 has attracted the most attention in the nephrology field, as it was found to be dysregulated in a multitude of kidney disorders, including X-linked hereditary nephropathy in dogs, polycystic kidney disease, diabetic nephropathy, and kidney cancers. 10 –15 Recently, it was shown that renal miR-21 is upregulated in Col4α3 −/− mice on the 129/SvJ background, and administration of anti-miRNA-21 oligonucleotides, a chemically modified single stranded RNA molecule with full sequence complementarity to miR-21, significantly slows kidney disease progression and improves survival in Alport mice. 16 Currently, anti-miR-21 (RG-012) is in clinical development for Alport syndrome in a Phase II clinical trial. 17
Though miR-21 expression has been studied in multiple animal models of kidney disease, including Alport syndrome, miR-21 expression patterns in kidneys from patients with Alport syndrome have not been reported. The goals of the current study were to investigate miR-21 expression in kidneys from patients with Alport syndrome and to correlate its expression with measurements of kidney function, kidney histopathological scores, and CKD stages. In addition, some gene markers of key molecular pathways implicated in the pathophysiology of Alport syndrome were quantified in both human and mouse kidney samples. Furthermore, the study explored how neutralization of miR-21 altered the expression patterns of these pathways in Alport mice.
Methods
Selection and collection of human kidney samples
Paraffin-embedded kidney specimens from 27 patients with Alport syndrome (15 CKD stage 1 and 12 CKD stages 2–4; stage diagnosis was based on KDIGO 2012 Clinical Practice Guidelines 18 ) were randomly selected from samples collected between 2002 and 2015 at the Department of Pediatrics in Peking University First Hospital, Beijing, P.R. China. Aside from a positive family history and characteristic symptoms, Alport syndrome was confirmed by at least two of the following methods: kidney biopsy, skin biopsy, or genetic testing. Tissue samples of the unaffected portion of kidney from patients undergoing nephrectomy for renal carcinoma were used as normal controls. The study included 10 normal controls (7 male) aged between 35 and 49 years (median 45 years). All control samples were collected in 2015. A routine urinary test showed protein was negative for all normal controls. All control samples were reviewed by renal pathologists, and only samples demonstrating normal kidney morphology without carcinoma cells, fibrosis, or inflammation were included in the current study as controls.
All kidney biopsies were collected by fine-needle aspiration before being fixed in 10% formalin and subsequently embedded in paraffin blocks prior to sectioning. Patient information including age, sex, height, weight, and laboratory parameters of serum and urine at the time of biopsy was obtained from the database of the Department of Pediatrics in Peking University First Hospital. The study was approved by the Ethics Committee of Peking University First Hospital, and all patients signed a document confirming their informed consent for study participation.
RNA isolation and quantitative polymerase chain reaction for human kidney samples
Total RNA, including small and miRNA, was isolated from human kidney samples embedded in paraffin blocks with a miRNeasy FFPE Kit (Qiagen; cat. no. 217504) following the manufacturer's protocol. One 5 μm section was freshly cut from each paraffin block and used for isolation. RNA concentration was measured with a NanoDrop spectrophotometer.
miRNA-21 relative expression was measured by TaqMan miRNA assays (Life Technologies; cat. no. 4427975) following the manufacturer's protocol. Briefly, 5 ng of total RNA was reverse transcribed with a TaqMan MicroRNA Reverse Transcription Kit (Life Technologies; cat. no. 4366596) with specific reverse transcription primer for each miRNA. Reverse transcription product was assayed in a TaqMan miRNA reaction (Life Technologies; miR-21 assay ID 000397; miR-16 assay ID 000391). miRNA-16 served as the endogenous control.
Gene expression was measured using TaqMan gene expression assays (Life Technologies; cat. no. 4331182) following the manufacturer's protocol. Briefly, total RNA was reverse transcribed to cDNA by SuperScript™ IV VILO™ Master Mix (Life Technologies; cat. no. 11766050). Then, the reverse transcription product was assayed in a TaqMan gene expression reaction. Genes measured were collagen type I alpha 1 chain (COL1A1; Life Technologies; assay ID: Hs00164004_m1), epidermal growth factor (EGF; Life Technologies; assay ID: Hs01099999_m1), superoxide dismutase 1 (SOD1; Life Technologies; assay ID: Hs00533490_m1), and importin 8 (IPO8; Life Technologies; assay ID: Hs00183533_m1). IPO8 served as the endogenous control.
All TaqMan reactions were performed in triplicate on 384-well plates on an ABI 7900HT Sequence Detection System (Life Technologies). Temperature cycles were as follows: 50°C for 2 min, 95°C for 10 min, followed by 40 cycles of 95°C for 10 s and 60°C for 1 min. Expression levels were calculated by using the comparative CT (ΔΔCT) method. Results were normalized to endogenous control, and expression levels are shown as the fold change relative to normal control group.
miR-21 in situ hybridization for human kidney samples
Two 5 μm serial sections were cut from each paraffin block and used for in situ hybridization (ISH), one for hybridization with DIG-labeled locked nucleic acid (LNA) probe for miR-21 and the other one for hybridization with scramble control probe (miRCURY LNA miRNA ISH Optimization Kits; Exiqon). ISH was performed using an automated procedure on a Discovery Ultra instrument (Ventana Medical Systems, Inc.). The hybridization temperature was 58°C, and the hybridization time was 2 h. Only samples with clean or very low signal of scramble control staining were included in the result analysis.
Histopathology analysis of human kidney samples
Histopathology scoring was performed by two independent nephropathologists who were blinded to the slide information. The pathology scoring criteria were modified from previously published Remuzzi criteria. 19 Scores were defined by the percentage of tissue exhibiting pathological changes out of the total reviewed area. Multiple views were evaluated for each sample, and the mean value was used as the final score. The detailed pathological scoring criteria were: 0 = no pathological change is observed; 0.5 = pathological region is <10%; 1 = pathological region is between 11% and 20%; 1.5 = pathological region is between 21% and 35%; 2 = pathological region is between 36% and 50%; and 3 = pathological region is >51%
Animals
All animal experiments were conducted under protocols approved by the Institutional Animal Care and Use Committee of Sanofi. The Col4α3 knockout mice on the 129/SvJ background, 129-Col4α3 tm1Dec/J, were obtained from The Jackson Laboratory. The Col4α3 knockout mice on the C57BL/6J background were congenic and generated by Sanofi by backcrossing the 129-Col4α3 tm1Dec/J to the C57BL/6J background for more than 10 generations. Female Col4α3 heterozygotes on the C57BL/6J background were bred with male Col4α3 heterozygotes on the 129/SvJ background to produce the B6;129-Col4α3 tm1Dec (Col4α3 −/−) mice and their wild-type (WT; B6;129-Col4α3 +/+) littermate controls. Only the mice from the first generation of each cross were used in the studies.
Animal studies with anti-miR-21 treatment
Starting at 5 weeks (early-stage disease) or 9 weeks (late-stage disease) of age, Col4α3 −/− mice were treated with anti-miR-21 (RG-012) by subcutaneous injection twice per week at 25 mg/kg. Anti-miR-21 was provided by Regulus Therapeutics. Phosphate-buffered saline (PBS)-treated Col4α3 −/− and untreated WT were included as controls.
Serum BUN measurement in mice
Blood samples were collected at different ages or at various time points during the studies. Whole blood was collected by orbital bleeding in serum separator tubes (BD Microtainer; cat. no. 365967). Then, serum was separated by centrifugation. Serum samples were stored at −80°C until analysis. Serum BUN was measured on a Cobas Integra 400 plus analyzer (Roche Diagnostics) following the manufacturer's protocol.
Urine collection and urinary albumin measurement in mice
Twenty-four-hour urine samples were collected for each individual mouse by using murine metabolic cages. Urine samples with fecal contamination were excluded. Urine samples were centrifuged at 1500 g for 15 min at 4°C, and debris-free urine samples were stored at −80°C until analysis. Urinary albumin concentration was determined by enzyme-linked immunosorbent assay (Albuwell/M kit; cat. no. 1011; Exocell, Inc.). Urinary creatinine concentration was measure on a Cobas Integra 400 plus analyzer (Roche Diagnostics). Urinary albumin levels are presented as the albumin-to-creatinine ratio (ACR; mg/g), which was calculated by dividing albumin concentration in milligrams by creatinine concentration in grams. 20
RNA isolation, miR-21 relative expression, and transcriptomic analysis of mouse kidney samples
Mouse kidney samples were collected at different ages or at various time points during the animal studies. After the animals were euthanized, the kidneys were quickly removed and snap frozen in liquid nitrogen. All samples were stored at −80°C until RNA isolation. Total RNA, including small and miRNA, was isolated with a miRNeasy Mini Kit (Qiagen; cat. no. 217004) according to the manufacturer's instructions. Renal miR-21 levels were quantified with TaqMan miRNA assays as described above for human samples and expressed as fold change relative to littermate WT controls. Transcriptomic analysis was conducted following the Illumina TruSeq mRNA-Seq protocol, with at least 15 million reads for each sample. All FASTQ files from the RNA-Seq experiment were processed with Omicsoft Array Studio v6.2. Sequence reads were mapped to the reference mouse genome (Genome Reference Consortium Build 37) by the Omicsoft Aligner. Alignments were then quantified into FPKM values 21 at the gene level using the gene model from “RefGene20121217.” For differential expression analysis, log2 (FPKM +1) values were used for Student's two-tailed t-test on Col4α3 −/− versus WT at different ages over the disease progression. Similarly, log2 (FPKM +1) values were used for one-way analysis of variance (ANOVA) to compare PBS-treated Col4α3 −/− versus WT and anti-miR-21 versus PBS-treated Col4α3 −/−. Hierarchical clustering was conducted for differentially expressed genes after applying a statistical threshold (p < 0.01 and absolute fold change >1.5).
miR-21 ISH for mouse kidney samples
Mouse kidney samples were harvested at different ages. After the animals were euthanized, the kidneys were quickly removed and fixed in 10% neutral-buffered formalin for a minimum of 24 h before being embedded in paraffin. ISH of miR-21 for mouse kidney was performed by following the same protocols as described above for human kidney samples.
Statistical analysis
Statistical analysis was performed using GraphPad Prism v7 (GraphPad Software, Inc.). To compare human clinical characteristic parameters, human pathological scores, human renal miR-21 relative expression, human gene expression levels, mouse renal miR-21 relative expression, serum BUN, and ACR in Col4α3 −/− mice with anti-miR-21 treatment, the Kruskal–Wallis test (with multiple comparisons corrected by Dunn's test when more than two groups were presented) was applied. Pearson's and Spearman's correlation were used to analyze the correlation between the miR-21 relative expression and the kidney parameters of the Alport patients and Col4α3 −/− mice. To compare mouse data showing changes over time, two-way ANOVA with multiple comparisons was applied, corrected by Tukey's test. The survival distributions of mice between anti-miR-21 and PBS-treated groups were compared by log-rank test with a two-sided significance level of 0.05.
Results
Patient demographics for human kidney samples
Demographic and clinical data for the patients with Alport syndrome are summarized in Table 1. In total, 27 pediatric patients with Alport syndrome were categorized into CKD stage 1 (n = 15) and CKD stages 2–4 (n = 12) based on their estimated glomerular filtration rate (eGFR) at the time of biopsy. A total of 10 morphologically normal kidney specimens were included as normal control samples; these samples were obtained from adult patients, since nephrectomy surgery in a pediatric population due to renal carcinoma is exceedingly rare.
Clinical characteristics of AS samples enrolled in the current study
Values are presented as median (interquartile range) or number of samples.
eGFR was calculated by modified Schwartz estimation: eGFR = 0.413 × [height (cm)/serum creatinine (mg/dL)].
p < 0.05 vs. AS CKD stage 1 group.
AS, Alport syndrome; CKD, chronic kidney disease; eGFR, estimated glomerular filtration rate.
Due to the chronic progressive nature of Alport syndrome, the median age, height, and body weight for the CKD stages 2–4 group were greater than those of the CKD stage 1 group. Likewise, the Alport CKD stages 2–4 group had increased levels of serum creatinine (median 1.38 mg/dL), which was significantly higher than the Alport CKD stage 1 group (median 0.44 mg/dL). The Alport CKD stages 2–4 group had decreased eGFR (median 51 mL/min ·1.73 m2), which was significantly lower than the Alport CKD stage 1 group (median 103 mL/min ·1.73 m2).
All Alport patients had proteinuria, while all normal controls had undetectable levels of urinary protein. The Alport CKD stages 2–4 group had more severe proteinuria (median 2.29 g/24 h) than the Alport CKD stage 1 group (median 0.32 g/24 h).
Renal miR-21 is upregulated in patients with Alport syndrome
To evaluate renal miR-21 expression in human Alport kidneys, relative miR-21 expression was quantified using RNA extracted from paraffin-embedded kidney tissue from either kidney biopsy or nephrectomy specimens. qPCR measurement showed Alport kidney specimens had significantly higher levels of miR-21 expression compared to normal controls, with a median fold change of 2.3 (interquartile range [IQR] = 1.7–3.5) in the CKD stage 1 group and a median fold change of 3.6 (IQR = 2.5–4.8) in the CKD stages 2–4 group compared to normal controls (p < 0.05 for both Alport groups compared to normal control group). Furthermore, the renal miR-21 expression level in patients at CKD stages 2–4 was significantly higher than in patients at CKD stage 1 (p < 0.05; Fig. 1A).

Renal microRNA-21 (miR-21) expression in patients with Alport syndrome (AS) compared to normal controls.
To determine the cellular localization of miR-21 expression in Alport kidneys, ISH was performed with a miR-21 probe and scramble control probe on paraffin-embedded kidney sections. Due to the technical challenges of miR-21 ISH assay, especially on archived paraffin-embedded kidney biopsy specimens, 6/15 specimens from the CKD 1 group and 5/12 specimens from the CKD stages 2–4 group passed the quality control with clean or very low signal of scramble control staining. On the other hand, 9/10 specimens from normal controls passed the quality control. Consistent with the qPCR results, ISH showed increased miR-21 expression in Alport specimens while being virtually undetectable in normal control samples (Fig. 1B). The miR-21 staining was more pronounced in samples from CKD stages 2–4 compared to those from CKD stage 1 (Fig. 1B). The increase in miR-21 expression was limited to tubular epithelial cells in the areas where interstitial fibrosis was demonstrated by Masson's trichrome staining on serial sections (Fig. 1C). Increased miR-21 signals were also detected in glomeruli, along the capillaries, and in the renal interstitium around Bowman's capsule in Alport CKD stages 2–4 patients (Fig. 1D).
Renal miR-21 expression correlates with clinical biochemical measurements and kidney pathology scores in patients with Alport syndrome
To evaluate the relationship between renal miR-21 expression and disease progression, correlation analyses were done between relative miR-21 expression and clinical biochemical parameters in patients with Alport syndrome. Relative kidney miR-21 expression positively correlated with 24 h urinary protein (r = 0.71, p < 0.001) and serum creatinine concentration (r = 0.41, p < 0.05). More importantly, relative kidney miR-21 expression negatively correlated with eGFR (r = −0.44, p < 0.05; Fig. 2A–C).
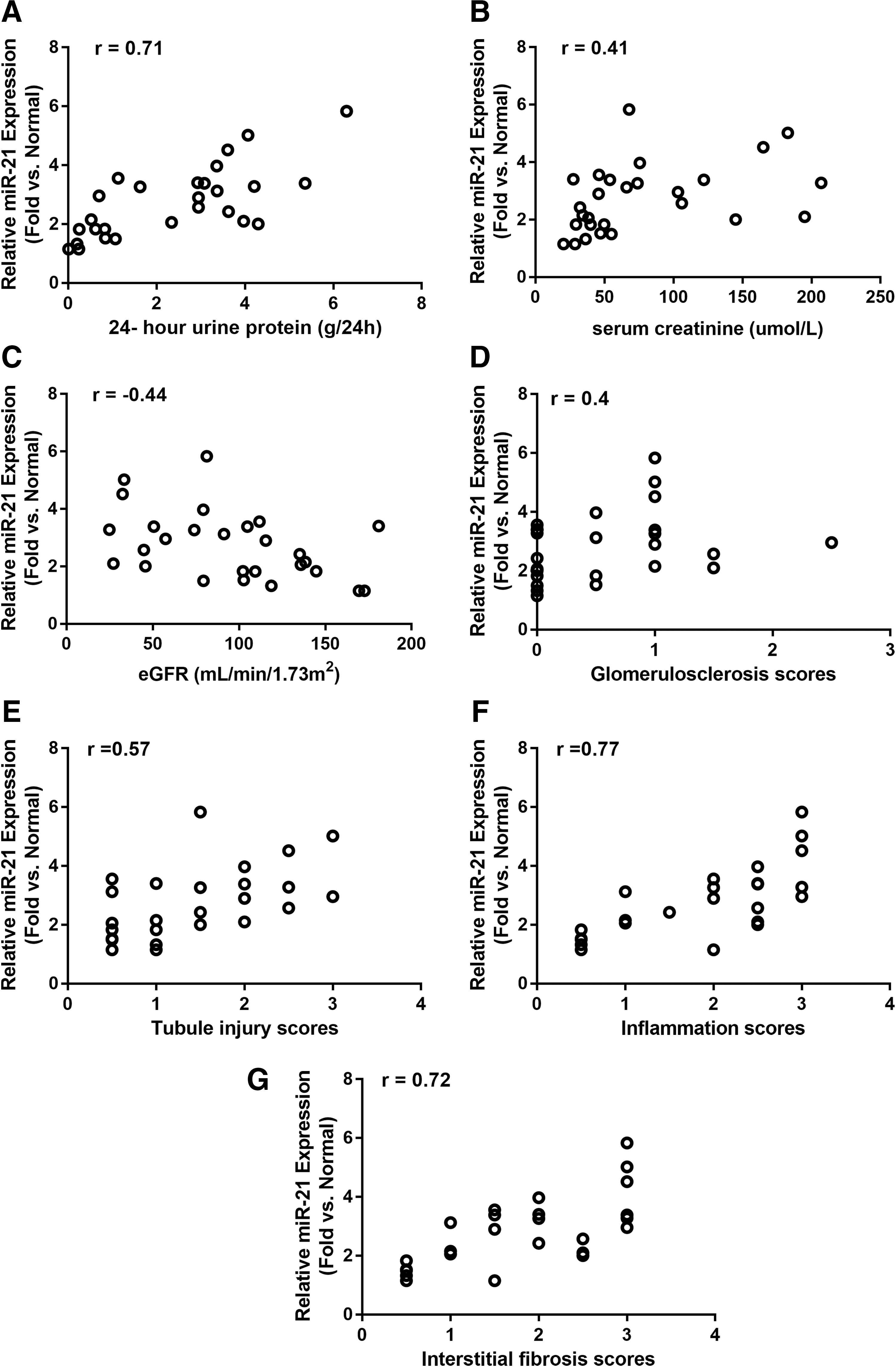
Pearson's correlation between human AS patients' relative renal miR-21 expression and laboratory parameters at the time of renal biopsy:
To study the relationship between miR-21 expression and disease stages, the kidney samples were scored using Remuzzi criteria, including scores for glomerulosclerosis, tubular atrophy, interstitial inflammation, and interstitial fibrosis. The pathology scoring for each group is summarized in Table 2. The kidney samples from patients with Alport syndrome had significantly higher pathology scores, including higher rates of glomerulosclerosis, tubular atrophy, interstitial inflammation, and interstitial fibrosis, compared to the normal control group. All of these scores further increased in samples from patients with CKD stages 2–4 compared to samples from patients with CKD stage 1 (Table 2). Relative renal miR-21 expression in Alport samples positively correlated with all four histopathological scores studied, including glomerulosclerosis (r = 0.40, p < 0.05), tubular atrophy (r = 0.57, p < 0.01), interstitial inflammation (r = 0.77, p < 0.001), and interstitial fibrosis (r = 0.72, p < 0.001; Fig 2D–G).
Renal pathological characteristics of normal control tissues and renal biopsy samples from AS patients
Values are presented as median (interquartile range).
p < 0.05 vs. normal control group; # p < 0.05 vs. Alport CKD stage 1.
B6;129-Col4α3 tm1Dec homozygous mice in mixed C57Bl6/J and 129X1/SvJ genetic background exhibit kidney phenotypes similar to human Alport syndrome
To address the pathological role of miR-21 in the development of kidney disease in Alport mice, B6;129-Col4α3 tm1Dec homozygous mice (Col4α3 −/−) were generated by crossing heterozygous Col4α3 tm1Dec females (Col4α3 –/+) on C57Bl6/J background with heterozygous Col4α3tm1Dec males (Col4α3 –/+) on 129X1/SvJ background. Only B6;129-Col4α3 tm1Dec homozygous mice (Col4α3 −/−) and their WT littermates (B6;129-Col4α3 +/+) from the first generation of each cross (F1 hybrid of C57Bl6/J and 129X1/SvJ) were used in the studies. The B6;129-Col4α3 tm1Dec homozygous mice developed slower kidney disease progression compared to Col4α3tm1Dec mice on the 129X1/SvJ background (data not shown). In addition to albuminuria, Col4α3 −/− mice exhibited a significant increase in serum BUN at 9 weeks of age, which progressively increased and reached end-stage kidney disease by 15 weeks of age, suggesting they closely recapitulate the CKD stages in patients with Alport syndrome (Fig. 3A). Relative renal miR-21 expression measured by qPCR progressively increased in Col4α3 −/− mice compared to WT mice starting at 9 weeks of age (Fig. 3B) and demonstrated a positive correlation with serum BUN (Fig. 3C).

Renal function and miR21 expression in B6;129-Col4α3
tm1Dec (Col4α3−/−) mice compared to littermate wild-type (WT) controls.
Consistent with qPCR results, ISH of miR-21 in the kidneys of Col4α3 −/− mice showed a progressive increase of miR-21 expression correlating with disease severity (Fig. 3D). Higher magnification images showed increased miR-21 signals in cortical renal tubules where tubular necrosis and regeneration were observed and in Bowman's capsule where parietal cell hypertrophy and hyperplasia in glomeruli were detected (Fig. 3E).
Anti-miR-21 treatment improves the survival of Col4α3 −/− mice by slowing kidney disease progression at both early and late stages
Anti-miR-21 was administered by subcutaneous injection twice per week at 25 mg/kg to Col4α3 −/− mice; untreated littermate WT and PBS-injected littermate Col4α3 −/− mice were included as controls. Treatment started at either 5 or 9 weeks of age, as indicated in Fig. 4A.

Anti-miR-21 protects Col4α3
−/− mice from kidney disease progression and improves survival. Col4α3
−/− mice were subcutaneously injected with anti-miR-21 at 25 mg/kg or phosphate-buffered saline (PBS) twice per week.
Anti-miR-21-treated Col4α3 −/− mice had significantly lower renal miR-21 expression than PBS-treated Col4α3 −/− mice at 15 weeks of age (Fig. 4B). Anti-miR-21 treatment significantly delayed the onset of kidney disease, as demonstrated by a slower rise in BUN, compared to PBS-treated Col4α3 −/− mice when treatment started at 5 weeks of age (Fig. 4C). Similarly, in comparison to PBS-treated Col4α3 −/− mice, anti-miR-21-treated Col4α3 −/− mice showed a significant reduction in albuminuria beginning at 6 weeks of age (Fig. 4D). The attenuation of albuminuria could be detected within 1 week of initiation of anti-miR-21 therapy. More importantly, anti-miR-21 treatment significantly improved the survival of Col4α3 −/− mice compared to PBS treatment (log-rank p < 0.0001), increasing the median survival time from 17.5 to 22 weeks in Col4α3 −/− mice (Fig. 4E).
To test whether anti-miR-21 therapy is effective at a later stage of kidney disease in Alport mice, anti-miR-21 treatment was initiated at 9 weeks of age. It was found that anti-miR-21 treatment significantly attenuated kidney disease progression characterized by significantly reduced BUN and a trend of reduction of urine ACR measured at 15 weeks of age in Col4α3 −/− mice. Anti-miR-21 also improved overall survival compared to PBS-treated controls (log-rank p < 0.0001), increasing the median survival time from 17.8 to 18.8 weeks in Col4α3 −/− mice (Fig. 4F–H).
Genes mediating kidney injury, fibrosis, and mitochondria function are dysregulated in the kidneys of Alport patients
To understand the pathogenic role of abnormal kidney miR-21 expression in humans with Alport syndrome further, relative expression of genes involved in kidney injury, fibrosis, and mitochondria function were measured in human kidney samples. It was observed that the kidney injury marker EGF was significantly decreased in kidneys of patients with Alport syndrome at CKD stage 1 and further reduced in later disease stages. Interestingly, renal miR-21 and EGF expression levels demonstrated a strong negative correlation (r = −0.63, p < 0.001). The fibrosis marker COL1A1 was significantly increased in the kidneys of patients with Alport syndrome at CKD stage 1 and further increased in later disease stages. The mitochondria function marker SOD1 was significantly decreased in Alport patients (Fig. 5A–D).
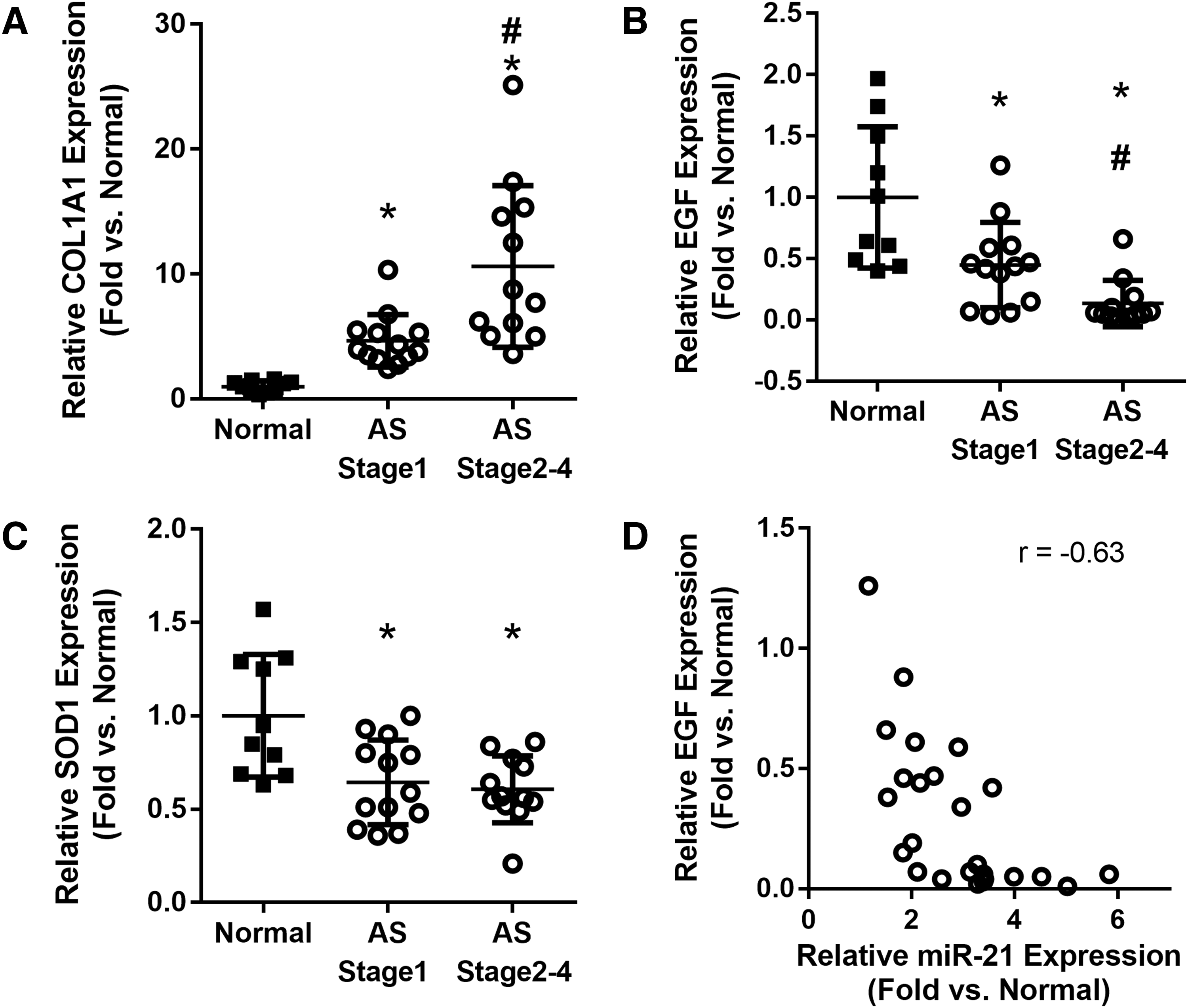
Renal gene expression in AS patients.
Dysregulated renal transcriptomic profiles were partially reversed by anti-miR-21 treatment in Col4α3 −/− mice
For a molecular understanding of disease progression in Alport syndrome, RNA-Seq profiling of kidney samples from Col4α3 −/− mice and littermate WT mice was conducted at 6, 9, 12, and 15 weeks of age. At 15 weeks of age, 6,435 genes were identified as differentially expressed between Col4α3 −/− and WT mice after applying a statistical threshold (p < 0.01 and absolute fold change >1.5), which were hierarchically clustered in Fig. 6 showing Col4α3 −/− versus WT at different ages. Specific genes representing different pathways are listed in Table 3. There are clear temporal progression patterns for both up- and downregulated genes in Col4α3 −/− mice, with the magnitude of fold changes increasing with animal age (Fig. 6 and Table 3).

RNA-Seq analysis of kidneys from Col4α3 −/− and littermate WT mice. Hierarchical clustering of renal gene expression profiles comparing Col4α3 −/− vs. littermate WT mice over disease progression. A total of 6,435 differentially expressed genes were selected based on week 15 data after applying a statistical threshold (p < 0.01 and absolute fold change >1.5). Kidney samples represented by columns of the clustering heat map are sorted by the grouping of corresponding animal ages and genotypes. Only a subset of gene symbols is labeled in the clustering dendrogram. n = 5–6 for each group at each time point.
Renal gene expression changes in Col4α3−/− mice compared to littermate wild-type mice
Average fold changes and t-test p-values are shown. For each age group, t-tests were conducted on log-transformed FPKM values of representative genes in top-ranked differentially expressed pathways between Col4α3 −/− and wild-type mice, n = 5–6 in each group at each time point.
The study also examined the kidney transcriptomic profiles of Col4α3 −/− mice receiving anti-miR-21 or PBS treatment from 5 to 15 weeks of age, as well as their WT controls. Comparing anti-miR-21 versus PBS treatment in Col4α3 −/− mice, 2,932 genes were identified as differentially expressed after applying a statistical threshold (p < 0.01 and absolute fold change >1.5), which were hierarchically clustered and compared to the WT group (Fig. 7). This data set not only confirmed abnormal gene expression profiles in the kidneys of Col4α3 −/− mice compared to WT shown in Fig. 6 and Table 3, but also demonstrated that anti-miR-21 treatment partially reversed the transcriptomic dysregulation in Col4α3 −/− mice. Anti-miR-21 treatment significantly reversed the abnormal expression of representing markers of kidney injury (e.g., Egf and Kim-1), fibrosis (e.g., Acta2, Ctgf, Fn1, Col1a1, Col3a1, Mmp2, Mmp7, Mmp9, and Tgfβ1), inflammation (e.g., Il1b, Il6, Ccl2, and Tnfrsf1b), mitochondrial function (e.g., Sod1, Sod2, and Sod3), and lipid metabolism (e.g., Ppara) in Col4α3 −/− mice (Fig. 7).
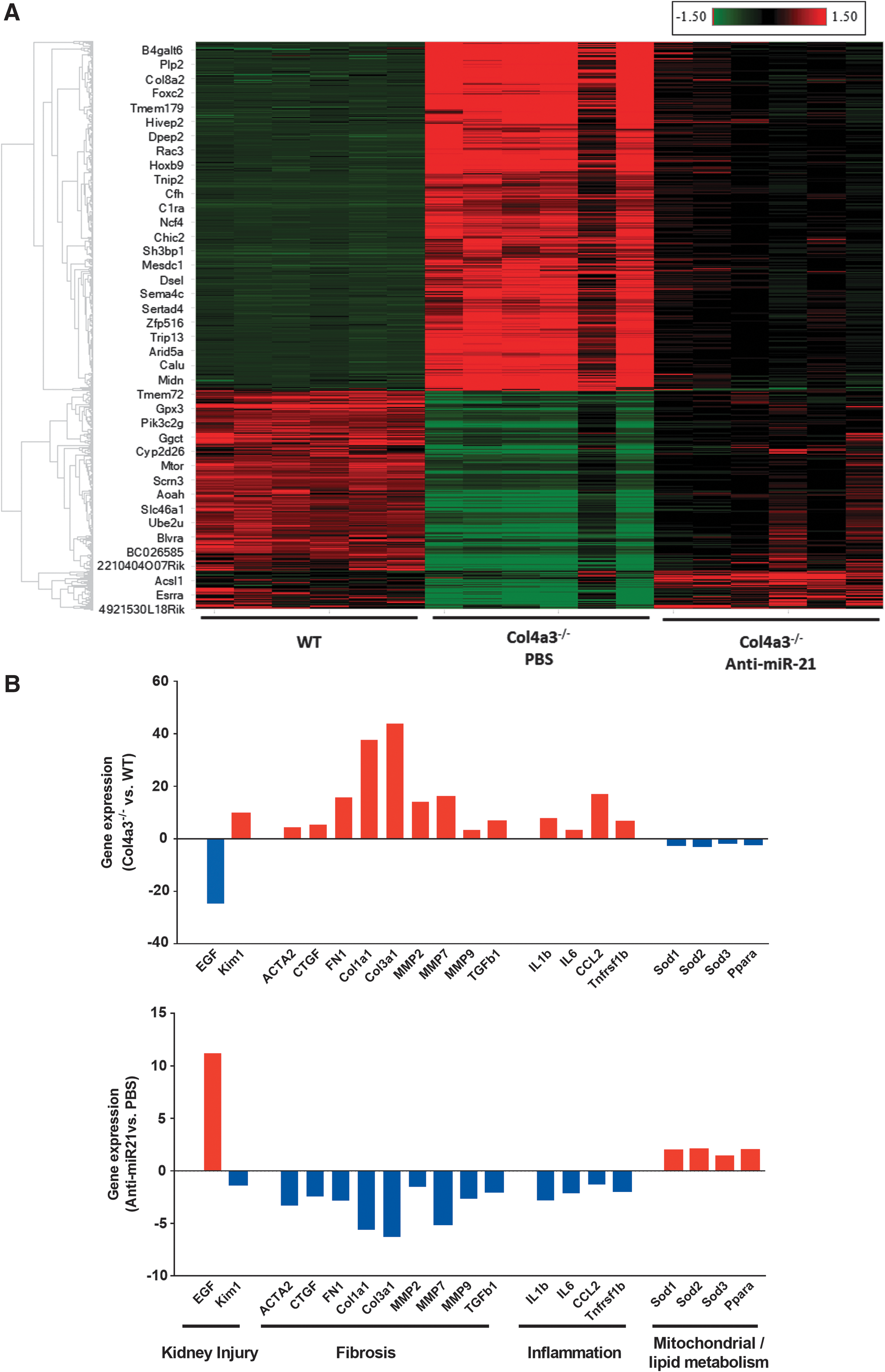
RNA-Seq analysis of kidneys from Col4αα3
−/− mice receiving anti-miR-21 or PBS treatment from 5 to 15 weeks of age.
Discussion
miRNAs are potent regulators of mRNA stability and have become of interest as possible therapeutics or therapeutic targets. Among the miRNAs of interest, miR-21 has been shown to play a very crucial role in many biological processes, including development, cancer, cardiovascular diseases, apoptosis, and inflammation. 22 Recent reports have suggested that anti-miR-21 might be a potential therapeutic target for the treatment of kidney diseases, including Alport syndrome. 16,23 –25 The importance of miR-21 in the development of kidney disease in Alport syndrome and other kidney diseases has been demonstrated by genetic deletion of miR-21 or blocking miR-21 using anti-miR-21 oligonucleotide in animal models. 11,16,23 A previous study in Alport mice demonstrated that pharmacological silencing of miR-21 leads to dramatic disease attenuation by stimulating multiple metabolic pathways. 16
More recently, it was demonstrated that expression of miR-21 is also increased in kidney tissue of dogs with X-linked Alport kidney disease. 10 The present study reports that similar to murine and canine Alport kidneys, miR-21 expression is increased in human kidneys from patients with Alport syndrome using both qPCR and miR-21 ISH. More importantly, kidney miR-21 expression correlated with disease severity measured by 24 h total urine protein content, serum creatinine, eGFR, and kidney pathology scores. These data provide fundamental information for developing anti-miR-21 to treat human patients with Alport syndrome.
The results support miR-21 as a key and early pathogenic factor that plays an important role in kidney disease progression in Alport syndrome. In support of this notion, elevation of renal miR-21 expression was observed in the early stages of the disease before measurable kidney function decline, and miR-21 expression was further increased when the kidney disorder progressed to later stages evidenced by declined eGFR and other biochemistry parameters. This finding is in agreement with the observation that miR-21 upregulation precedes histological changes in the kidneys of Col4α3 −/− mice. 16 Moreover, expression of miR-21 had a negative correlation with renal function as measured by eGFR in Alport patients. This trend was more profound in the Col4α3 −/− mouse model, where renal miR-21 expression was strongly correlated with BUN (r = 0.85, p < 0.0001; Fig. 3C). ISH confirmed the same trend in human patients and Col4α3 −/− mice. More importantly, blockade of miR-21 by anti-miR-21 oligonucleotides was renoprotective and improved survival in Col4α3 −/− mouse models, even when treatment started at later disease stages.
Proteinuria is an important hallmark of glomerular diseases and a prognostic predictor of kidney disease progression. 26,27 The increase in urinary protein in glomerular disorders is not only a result of glomerular injury, but itself may elicit further kidney damage. 28 It was observed that renal miR-21 levels strongly correlated with proteinuria in patients with Alport syndrome. Similarly, glomerular miR-21 expression has previously been reported to be positively associated with the ACR in a diabetic nephropathy population. 29 Though the exact mechanism of upregulation of miR-21 and proteinuria in patients with Alport syndrome remains unclear, it is clear that anti-miR-21 oligonucleotides attenuated proteinuria in Alport mouse models, suggesting this therapy could potentially benefit human patients with Alport syndrome and possibly other kidney diseases with glomerular pathology.
To identify the cell population in which the dysregulated miR-21 expression may play a role, ISH was performed to localize miR-21 in Alport kidneys. The data clearly showed that the increased miR-21 signals were located in both glomeruli and tubules. More importantly, miR-21 staining co-localized to the areas with pathological changes in the kidneys of both human and mice with Alport syndrome, particularly in areas where interstitial fibrosis was most abundant, indicating that increase in miR-21 expression may play a critical pathological role in those areas. Moreover, the data demonstrated a strong correlation between renal miR-21 and pathological scores of interstitial fibrosis and inflammation. Gene expression confirmed that COL1A1, a renal fibrosis marker, was dramatically and progressively upregulated in human Alport kidneys and Col4α3 −/− mouse kidneys alike. Importantly, blocking miR-21 could effectively prevent the increase in Col1α1 expression as well as other representing fibrosis markers in Col4α3 −/− mice.
Regardless of the primary causes, fibrosis is a common downstream pathological feature of almost all CKDs and results from increased synthesis and deposition of extracellular matrix within the kidney, which is often accompanied by increased apoptosis, epithelial to mesenchymal transition, and inflammatory cell infiltration. 30 Multiple studies have reported the association of upregulated miR-21 with renal fibrosis. 25,31 –33 In addition to the findings in Alport syndrome, miR-21 has been reported to be strongly associated with renal fibrosis in immunoglobulin A (IgA) nephropathy where miR-21 expression is most prominent in areas demonstrating abnormal tissue architecture. 34 In agreement with this theory, global transcriptome sequencing of kidneys from Col4α3 −/− mice showed that multiple fibrosis genes were significantly and progressively upregulated during disease progression. Fibrosis signaling was ranked as one of the top dysregulated pathways, and upregulated fibrosis genes were partially reversed upon anti-miR-21 treatment in the current study, which is consistent with what has been published in a mouse model on a different genetic background. 16 Collectively, these data support miR-21 as a critical profibrotic regulator in Alport kidney disease, and targeting miR-21 may provide therapeutic benefit to the kidney disease of Alport syndrome and possibly renal fibrosis in general.
EGF is a secreted protein mainly expressed in the kidneys, especially in the renal tubules. 35 –37 EGF binds to EGF receptor as a part of the ErbB signaling pathway, which regulates growth, survival, proliferation, and differentiation in mammalian cells. 38 Intrarenal EGF mRNA levels correlate with the rate of eGFR loss in CKD. 35 Urinary EGF is strongly correlated with intrarenal EGF mRNA levels and interstitial fibrosis, 35,39 and may serve as a prognostic biomarker in CKDs, including children with Alport syndrome. 40,41 In this study, renal EGF expression level was significantly, dramatically, and progressively downregulated in Alport patients, which is in agreement with prior reports in other CKD conditions, including diabetic nephropathy, IgA nephropathy, focal segmental glomerulosclerosis, minimal change disease, membranous nephropathy, acute kidney injury, and lupus nephritis. 39,42 –45 The data showed a negative correlation between miR-21 and EGF in Alport patients. Furthermore, blocking miR-21 trended to normalize EGF expression in Alport mice. Those results support a close relationship between miR-21 and EGF, even though the mechanism of this relationship remains to be defined.
Superoxide excess has been implicated in the pathogenesis of several systemic diseases, including kidney diseases. 46 Oxidative stress is closely related to mitochondrial dysfunction, and mitochondrial genes are among the predicted targets of miR-21. 47 In the transcriptome analyses, mitochondrial dysfunction was ranked as one of the top dysregulated pathways in kidneys from Col4α3 −/− mice, and anti-miR-21 treatment partially reversed many of the genes in this pathway. 16 Sod1, Sod2, and Sod3 are the three isoforms of the superoxide dismutase, which are the major defense systems against the superoxide anion. 48 Consistent with previous reports that showed mitochondrial dysfunction in an Alport mouse model, 16 the current study showed significant downregulation of SOD1 in human Alport kidneys and Sod1–3 in Col4α3 −/− mouse kidneys. Anti-miR-21 treatment ameliorated their reduction in Col4α3 −/− mice. Expression of miR-21 has been shown to correlate SOD activity negatively in tissues from human gastric cancers, 49 and upregulation of miR-21 was observed when vascular smooth-muscle cells were treated with hydrogen peroxide. 50 Overexpressing miR-21 in vitro has been shown to suppress SOD2 and SOD3. 51 In addition, Ppara it is a miR-21 target gene and a principal regulator modulating energy and lipid homeostasis. 16,52, 53 Its changes and response to anti-miR-21 were previously reported in Alport mice in 129/SvJ background. 16 Consistent with what has been published already, in the current study, Ppara is downregulated in Col4α3 −/− mice, and anti-miR-21 tends to normalize its dysregulation. Though details of the exact roles miR-21 plays in oxidative stress and mitochondrial dysfunction are not established, mounting evidence supports a key link between them. This study is the first to explore the relationship between these changes in human patients with Alport syndrome.
Inflammation is an important aspect in CKD as well as Alport syndrome. 54,55 Inflammation is closely connected with multiple pathways, including fibrosis and mitochondrial dysfunction. 56,57 Therefore, it was not surprising to see that inflammation is among the most dysregulated pathways in Alport mouse kidneys. Similar to the other pathways discussed already, multiple inflammation genes were significantly and progressively upregulated in Alport mice and partially reversed by anti-miR-21 treatment.
Kim1 is an important kidney injury marker in both acute and chronic kidney diseases. 58 –60 Urinary Kim1 is widely studied as a renal disease biomarker and is believed to be a good predictor for kidney disease progression. 60 In the current study, renal Kim1 expression significantly and progressively increased in Alport mice, and anti-miR-21 significantly attenuated its dysregulation. This provides strong evidence of the renoprotective effects of blocking abnormally increased miR-21.
By analyzing the gene expression profiles, multiple genes/pathways that are dysregulated in the kidneys of Col4α3 −/− mice were identified, which may be critical to renal disease progression (Fig. 7B). The similarity of gene expression changes between humans and mice with Alport disease (Fig. 5 and Table 3) indicates that the dysregulated pathways associated with abnormal expression of miR-21 might be similar between mice and humans, which supports Col4α3 −/− as a good mouse model of Alport kidney disease at both pathological and molecular levels. Therefore, attenuation of the dysregulated genes/pathways by anti-miR-21 treatment in Alport mice (Fig. 7) may provide reliable information for treating human Alport disease. In addition, the current results from B6;129Col4α3 −/− mice are consistent with previously published data from Col4α3 knockout mice on the 129/SvJ background, which further confirms that the present observation represents a disease pathology associated the disease not genetic background. 16
One important limitation of the current study is that age-matched controls were not used for the human study. Since it is not possible to obtain kidney biopsy samples from children without disease, results were compared with adult samples from the normal portions of nephrectomy kidneys that were removed due to renal carcinoma. All slides from the controls samples were reviewed by pathologists to make sure that no renal pathological changes (e.g., inflammation or fibrosis) were present in these samples and that all control specimens were histologically normal.
To the best of the authors' knowledge, this is the first study to demonstrate upregulation of renal miR-21 in human patients with Alport syndrome and the correlation of elevated miR-21 with disease severity and disease stage. ISH showed that abnormally increased miR-21 expression is localized to the damaged tubular epithelia, glomeruli, and areas exhibiting interstitial fibrosis. Similar distribution of abnormally expressed miR-21 expression was observed in the kidneys of mice and humans with Alport kidney disease. Anti-miR-21 treatment is renoprotective and significantly improves survival in Col4α3 −/− mice at both early and late disease stages. Expressions of COL1A1, EGF, and SOD1 are dysregulated in the kidneys of human patients and in a mouse model of Alport syndrome. RNA-Seq revealed multiple pathways are dysregulated in Alport mice, and anti-miR-21 treatment partially corrects these changes. Taken together, the data support the role of miR-21 in kidney disease pathogenesis in Alport syndrome through fibrosis, inflammation, mitochondria, and lipid metabolism pathways, and suggest miR-21 as a potential therapeutic target for kidney disorder of Alport syndrome.
Footnotes
Acknowledgments
Funding for this research was provided by the following organizations: the National Key Research and Development Program of China (no. 2016YFC0901505), Beijing key laboratory of molecular diagnosis and study on pediatric genetic diseases (no. BZ0317), and Sanofi (Framingham, MA). We would like to thank Vijay Modur and Yong Xue for their insightful discussion.
Author Disclosure
W.S., J.B., E.Y.X., C.P., L.P., E.L., O.I.-B., and S.L. were employed by Sanofi. D.McK. was employed by Regulus Therapeutics and has a patent US20170369879A1 issued. The other authors declared no competing interests.