
Abstract
After the discovery and characterization of the adenovirus in the 1950s, this prevalent cause of the common cold and other usually mild diseases has been modified and utilized in biomedicine in several ways. To date, adenoviruses are the most frequently used vectors and therapeutic (e.g., oncolytic) agents with a number of beneficial features. They infect both dividing and nondividing cells, enable high-level, transient protein expression, and are easy to amplify to high concentrations. As an important and versatile research tool, it is of essence to understand the limits and advantages that genetic modification of adenovirus vectors may entail. Therefore, a retrospective analysis was performed of adenoviral gene therapy constructs produced in the same laboratory with similar methods. The aim was to assess the impact of various modifications on the physical and functional titer of the virus. It was found that genome size (designed within “the 105% golden rule”) did not significantly affect the physical titer of the adenovirus preparations, regardless of the type of transgene (e.g., immunostimulatory vs. other), number of engineered changes, and size of the mutated virus genome. One statistically significant exception was noted, however. Chimeric adenoviruses (5/3) had a slightly lower physical titer compared to Ad5-based viruses, although a trend for the opposite was true for functional titers. Thus, 5/3 chimeric viruses may in fact be appealing from a safety versus efficacy viewpoint. Armed viruses had lower functional and physical titers than unarmed viruses, while five genomic modifications started to decrease functional titer. Importantly, even highly modified armed viruses generally had good titers compatible with clinical testing. In summary, this paper shows the plasticity of adenovirus for various vector, oncolytic, and armed oncolytic uses. These results inform future generations of adenovirus-based drugs for human use. This information is directly transferable to academic laboratories and the biomedical industry involved in vector design and production optimization.
Introduction
Genetic engineering methods have enabled the modification of wild-type viruses for use as safe and effective gene therapy vectors for multiple diseases. 1 Among the most thoroughly characterized gene therapy vectors is the adenovirus, which also continues to be the most popular. 2 Wild-type adenoviruses commonly infect the upper respiratory, gastrointestinal, and urinary tracts, or the conjunctiva, causing mild and self-limiting disease. Occasionally, pneumonia or even encephalitis can occur in immunosuppressed individuals or children. Of note, rare cases of systemic dissemination of adenovirus demonstrate the ability of the virus to travel through blood into distant tissues, which has implications for the therapeutic use of tumor-selective viruses. 3 In this regard, it is not surprising that transduction of non-injected metastases has been seen in cancer patients. 3 –5
Adenovirus is a non-enveloped double-stranded DNA virus with a protein capsid. 6 The capsid structure encloses a genome consisting of roughly 36 kb pairs. 7 Conveniently, the virion structure allows insertion of complementary DNA (cDNA) elements for expression of foreign transgenes. In addition to immunostimulatory transgenes (e.g., cytokines and chemokines), non-immunostimulatory transgenes (e.g., tagging proteins such as luciferase, green fluorescent protein [GFP], or prodrug converting enzymes, and ion channels), and cDNA coding for full-length antibodies (e.g., anti-CTLA4 and anti-HER2) can be inserted into the adenovirus genome to generate recombinant adenovirus vectors. 8,9
For therapeutic gene transfer purposes, replication-deficient adenovirus vectors can be engineered. In this approach, adenoviral early genes (E1–E4, often just E1 and E3) are deleted and replaced with a transgene of interest. 6 The inserted genes are typically placed under a constitutive promoter element, such as the cytomegalovirus (CMV) or Rous sarcoma virus (RSV) promoter, which allow transgene expression in any transduced cell. For example, Advexin is an E1/E3-deleted serotype 5 adenovirus expressing wild-type p53 under CMV. 10 A similar virus, Gendicine, has been approved for routine use in China.
In another embodiment of the vector concept, adenoviruses coding for immunostimulatory cytokines have been constructed and used clinically for various cancer types; AdCAIL-2 and TG1024 are both interleukin 2–encoding non-replicating vectors explored in the treatment of melanoma. 11,12 Several other cytokines have also been inserted, including tumor necrosis factor alpha, 13 interferon beta, 14 and interferon gamma. 15 Prodrug converting enzymes have also been a popular approach, such as rAd/CPG2(Q3)-Thy1 coding for GPI anchored, prodrug converting carboxypeptidase G2. 16 Several vector approaches have been promising enough to make it to randomized Phase III testing, but adenoviral virotherapy agents for cancer treatment have not yet been approved outside of China. 13
In contrast to replication-deficient viruses, replication-competent oncolytic viruses are able to go through their entire life cycle in a tumor but not in normal cells, culminating in cell lysis and release of new virions. Tumor specificity can be achieved on the level of replication with tumor-specific promoters or deletions transcomplemented in tumor cells (transcriptional targeting). In addition, the virus capsid can be modified to increase infection of tumor cells, or to overcome lack of receptors that are downregulated in advanced cancers (transductional targeting). 6,17,18
In the former approach, tumor tissue–active promoters, such as human telomerase reverse transcriptase, carcinoembryonic antigen, cyclooxygenase, and E2F, have been used in oncolytic adenovirus constructs. 19 –22 Deletions of E1b55k or E1A constant region 2 have been employed to restrict the virus replication to cells defective in the p14ARF/p53 or p16/Rb pathways, respectively. Oncorine, an oncolytic adenovirus approved for the treatment of head and neck cancer in China, is an example of the former type of deletion mutant. Of note, it is logical to combine the benefits of vectored transgene delivery with the oncolytic platform capable of local amplification and systemic dissemination to metastases. 4
In addition to transcriptional targeting, the adenovirus fiber protein can be modified to feature either synthetic elements such as RGD-4C or structures from different serotypes. Often the goal is to avoid reliance on the Ad5 receptor (coxsackie adenovirus receptor [CAR]), which is known to be almost universally downregulated in advanced metastatic tumors due to its role in cellular adhesion. 23 To solve this issue, several adenovirus variations have been explored, such as Ad5/7, 24 Ad5/11, 25 Ad5/35, 26 and Ad3/Ad11p. 27 The clinically most studied chimera, however, is the 5/3 structure, which features the knob of serotype 3 in an otherwise Ad5 virion. 22,23,28,29 Again, there is no reason to avoid combining the benefits of transductional targeting with the armed oncolytic platform. 30
Heretofore, a systematic analysis of the effects of adenovirus modifications (i.e., capsid structure, transgene type, and overall number of changes) on viral vector yield has not been performed. Physical and functional titer data of circa 50 different adenoviral gene therapy constructs produced in the Cancer Gene Therapy Group (CGTG) at the University of Helsinki between 2002 and 2018 were gathered in order to assess the impact of various modifications on virus titers. Additionally, since it has been suggested that the amount of DNA that can be packaged into adenovirus virions is limited, 31,32 correlation analysis was performed to evaluate the effect of genome size on virus titers.
Methods
Adenoviruses
Adenovirus construction and production is described in detail in the sources found in Table 1. Typically, non-replicative viruses were constructed by insertion of a transgene into a cloning plasmid, which was then co-transfected with a “rescue plasmid” (containing the rest of the virus genome) into bacteria featuring the requisite eukaryotic recombination enzymes. Linearized plasmid was then propagated in HEK293 cells, for example, which provide the E1 region in trans. 23 Many replication competent (oncolytic) chimeric adenoviruses were constructed by an adapted BAC- recombineering method, 33 –35 and propagated in non-transcomplementing A549 cells to avoid the risk of back-recombination generating a modified but non-tumor selective virus. Produced virions were collected from cells showing cytopathic effect by trypsin detaching or scraping, centrifugation, and repeated freeze–thaw cycles. Downstream purification employed two consecutive cesium chloride density centrifugations followed by dialysis to remove cesium chloride.
List of adenoviruses
Serotype 5 fiber protein except for the knob, which is serotype 3.
Selectively replication competent in tumor cells: oncolytic virus.
Physical titer (virus particles [VP]) was assessed spectrophotometrically (OD260 reading). Functional titering was done with a 50% tissue culture infective dose (TCID50) assay in HEK293 or A549 cells, depending on virus type. TCID50 titers were converted into a more conventional plaque-forming units (PFU) titer using a formula. 18,36 VP/PFU ratios (physical to functional titer) were calculated by dividing VP titer by PFU titer.
Statistical analyses
All virus constructs included in the statistical analyses are listed in Table 1. All statistical analyses, Kruskal–Wallis tests, and Spearman's correlations were performed using GraphPad Prism v7 (GraphPad Software, Inc., San Diego, CA).
Results
Chimeric 5/3 adenoviruses have moderately lower physical but not functional titers compared to fully serotype 5 viruses.
Viruses with fully serotype 5 capsid structure had the highest VP titers, with an arithmetic mean of 4.2 × 1012 VP/mL and a median of 2.3 × 1012 VP/mL (n = 30 virus preparations; Fig. 1A and C). Chimeric 5/3 viruses had lower titers, with a mean of 2.4 × 1012 VP/mL and a median of 1.1 × 1012 (n = 88), and the difference to serotype 5 viruses was statistically significant (p = 0.0412). Viruses with other capsid modifications (RGD, pk7, pk21, RGD + pk7) had a similar trend for moderately lower VP titers (M = 2.4 × 1012 VP/mL, median = 1.9 × 1012, n = 6, not statistically significant; note smaller n) versus fully Ad5.
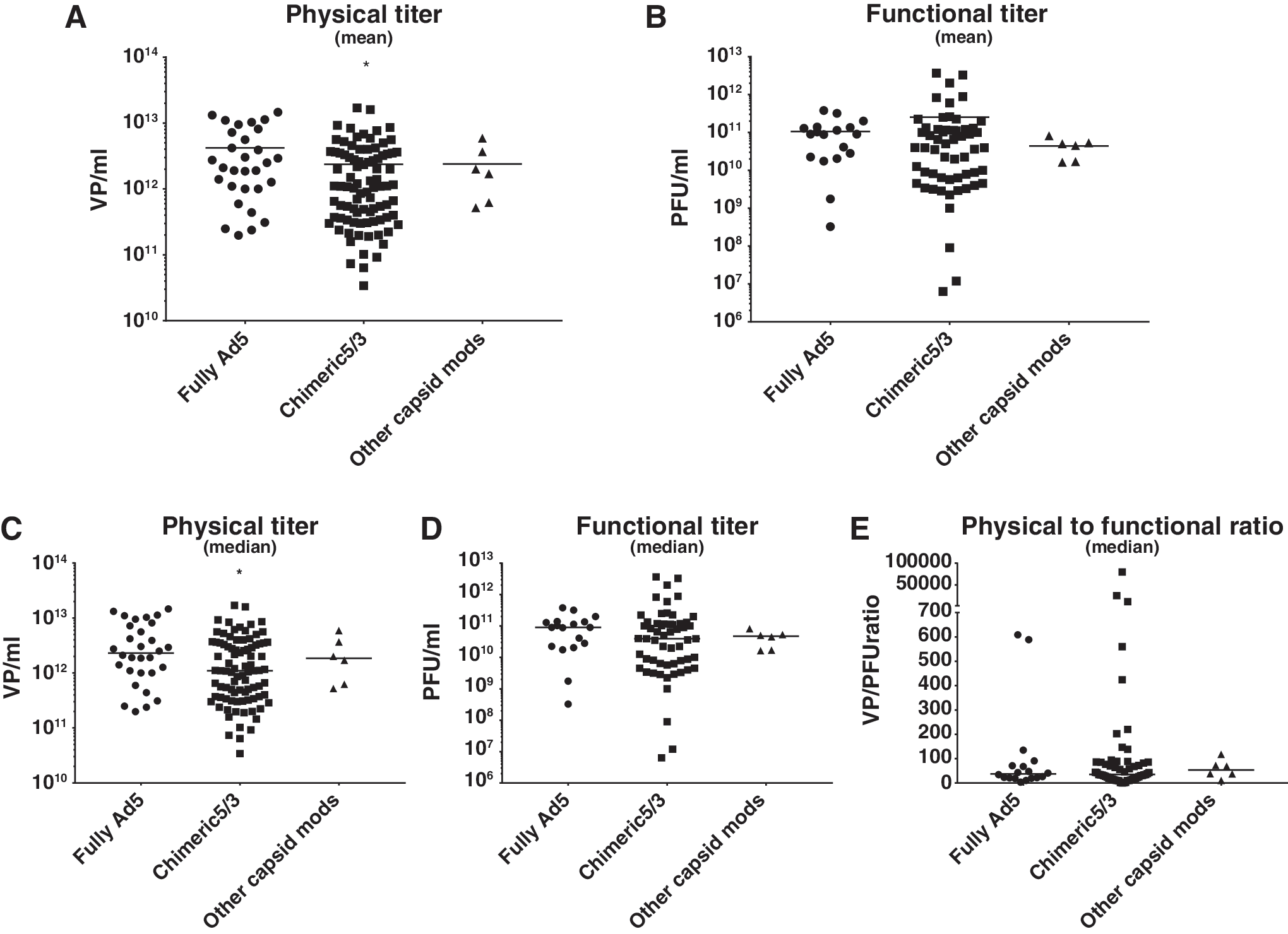
The effects of capsid structure modifications grouped according to the type of modification indicated on the graph.
Functional titers were not significantly different between fully Ad5, chimeric 5/3, and other capsid-modified adenoviruses. Ad5 mean PFU titer was 1.1 × 1011 PFU/mL, 5/3 chimeric virus mean was more than twice as high (2.5 × 1011 PFU/mL), while the mean of the other capsid modification group was lower (4.4 × 1010 PFU/mL). The respective medians were: serotype 5 = 9.0 × 1010 PFU/mL, chimeric 5/3 = 4.0 × 1010 PFU/mL, other modifications = 4.8 × 1010 PFU/mL (Fig. 1B and D).
Ratios of physical to functional titer were not significantly different between groups (Fig. 1E). The ratio was 37.6 for Ad5 and 35.2 for Ad5/3. Because some clear outliers were identified, medians were used for ratio calculations. Regarding outliers, Ad5/3lucS* 37 with shaft mutations had ratios ranging from 11,483 to 79,556, suggesting that this capsid configuration (or other properties of the virion) did interfere with virus production and function (Fig. 1E).
Transgene type significantly affects functional virus titers
Comparison of physical VP titers of viruses with different transgene types did not reveal significant differences (Fig. 2A). It is noteworthy, however, that the functional titer of virus groups with immunostimulatory (p = 0.0054), non-immunostimulatory transgenes (p = 0.0403), and antibody transgenes (p = 0.0131) were significantly lower compared to the control group of viruses with no added transgenes (Fig. 2B). Comparison of transgene groups revealed significantly lower ratios for the immunostimulatory group (p = 0.0016) and antibody groups (p = 0.027) compared to the group with no transgenes (Fig. 2C).

The effect of transgene type on adenovirus titers. The transgenes were classified as immunostimulatory, non-immunostimulatory, or antibodies. The effect of the produced transgene was evaluated by comparing the titers with the viruses not carrying any transgenes. Analysis of the effect of the above-mentioned groups on:
Number of modifications affects virus functionality
Viruses were grouped according to the number of functionally relevant, engineered, modifications. Kruskal–Wallis analysis did not indicate statistically significant differences between the number of modifications and physical/functional titers, or the ratios between physical and functional titers when fewer than five modifications were done. However, five genetic modifications caused a significant drop in the physical and functional titers (p = 0.139 and p = 0.002, respectively; Fig. 3).

Genome size does not correlate with physical or functional virus titer
Spearman correlation analysis was performed to compare functional titers with genome sizes. Wild-type adenovirus serotype 5 (Ad300wt strain purchased from ATCC, Manassas, VA) served as 100% reference value. When genome size was plotted against physical titer, no correlation was observed between the two factors (r = 0.04; Fig. 4A). Similar results, indicating a lack of correlation, were observed with functional titer (r = −0.06; Fig. 4B) and VP/PFU ratio (r = −0.03; Fig. 4C). Of note, there was no attempt to make viruses whose genome size would be <98% or >105% of the wild-type genome, and it seems unwise to extrapolate the findings beyond these limits.
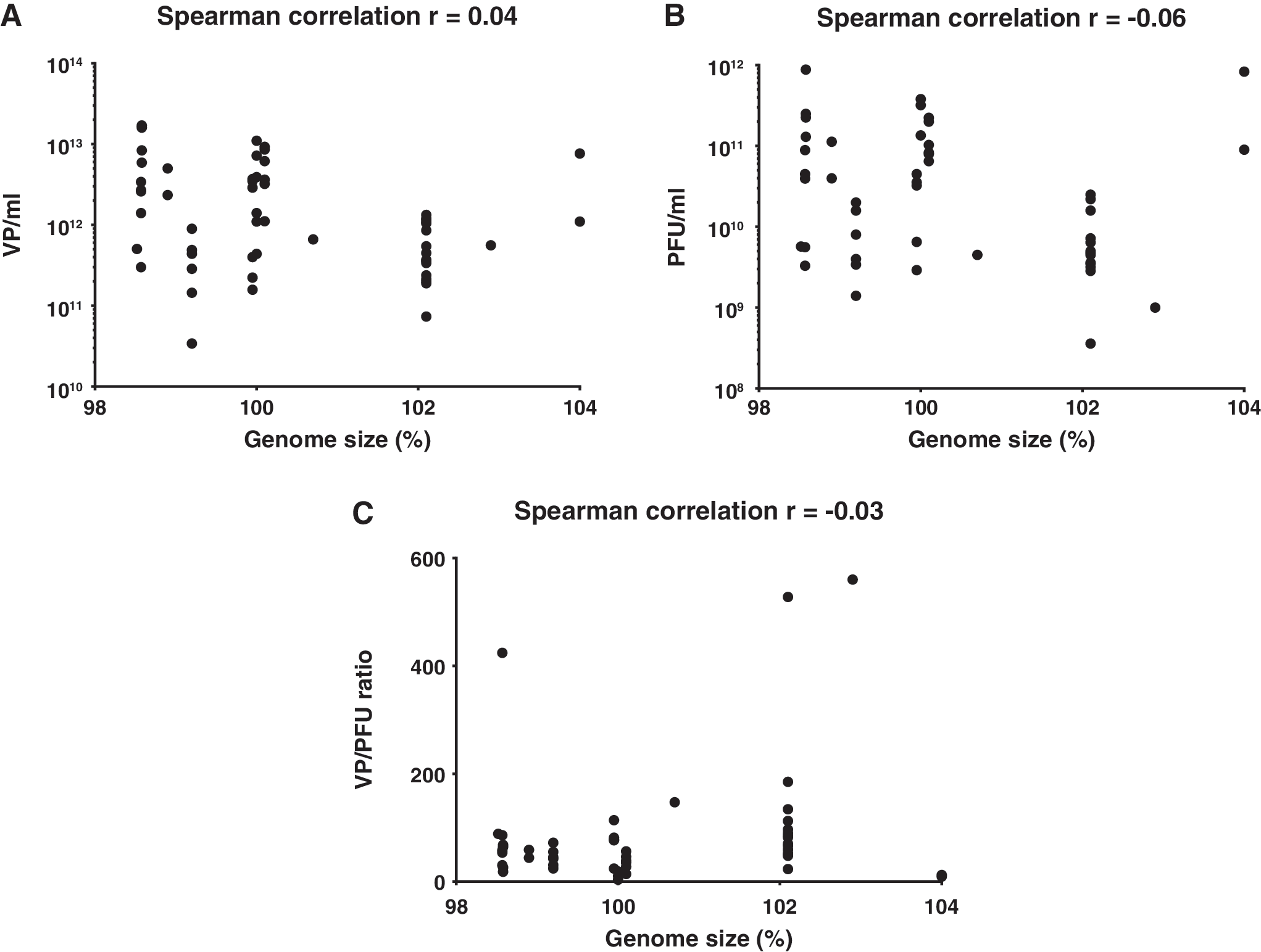
Correlation of virus genome size with physical and functional titer. Spearman's correlation analysis of physical and functional virus data
Discussion
The construction and production of biological therapeutic agents for human use is a heavily regulated field with a high demand of accuracy in order to provide safe and reliable drugs. Importantly, as adenoviruses are being utilized for several clinical and preclinical purposes, it is of essence to understand the effects of man-made modifications to the packaging ability of the virus. One of the main reasons for conducting modifications to adenovirus vectors relates to the concern regarding pre-existing immunity against the vector and subsequent vector-depleting immune response and toxicity. 38
The adenovirus capsid consists mainly of three components—fiber, penton, and hexon proteins—which are available for neutralizing antibodies. Of these, the fiber proteins alone do not seem to function as major targets for neutralizing antibodies, since the hexon proteins have been shown to be important immunoneutralizing targets as well. 39 Therefore, it is expected that while chimerism and other modifications to the fiber allow a degree of escape from pre-existing antibodies, 40 they do not dramatically affect the immunogenicity of the viruses. Thus, over time, antibodies will also develop against modified capsids. Of note, with increased understanding of selecting the right vector for each purpose, adenovirus has been very safe in the context of cancer therapy. 39,41 –43 Immunogenicity is an important tool when inducing antitumor immunity, while it might be a caveat when treating hereditary disease, for example.
In cancer therapy, the antiviral immune response is harnessed as a positive, immune system–activating, tumor-depleting force in the battle against tumors that are difficult to treat. In this regard, some early studies identified key issues regarding vector design, such as the association of genome size and virion stability. 7,31,32 They concluded that adenovirus vectors are unstable at sizes >39 kb, which is roughly 105% of the wild-type adenovirus genome size. This result has subsequently been applied as the gold standard when adenovirus vectors have been developed and designed.
Interestingly, however, even though many types of oncolytic and vector adenoviruses have been constructed, it is not known how modification type, the number of engineered modifications, capsid type, or replication competence affect stability and packaging of the virus. Lack of understanding of these issues is a major caveat in the field, since, on one hand, it is easy to attempt to construct almost any type of virus but, on the other hand, many of the modifications that can be envisioned might easily affect virus assembly.
For example, capsid is one of the key determinants in virion stability, so any changes to it could reduce titers. Moreover, transgenes added to the virus genome could potentially affect producer cells. Subsequently, it would not be surprising if the number of modification would eventually decrease the ability of the virus to replicate. In the course of evolution, each of the 57 adenovirus serotypes discovered to date have been optimized for stability and replication. Thus, man-made, engineered changes could result in decrease in these attributes. The adenovirus genome is read in both directions, and most of the DNA is coding sequence, indicating that any change can have unexpected changes in other adenovirus proteins.
Therefore, the effect of modifications was analyzed in vectors used and/or made in the CGTG at the University of Helsinki. Several modifications to the adenovirus fiber and/or knob have been employed, altering the specificity and efficacy of infection. This study found that virions with chimeric 5/3 fiber modifications produced a slightly but statistically significantly lower physical titer compared to virus preparations produced with unmodified Ad5 knob and shaft domains. In contrast, mean functional titers seemed higher with chimeric viruses. This is in accordance with previous findings indicating that chimeric viruses result in a higher rate of oncolysis and virus DNA to nucleus transportation, and more efficient replication efficacy compared to control viruses. 44
Efficacy, however, is a multifactorial endpoint influenced not only by virus replication and lysis of cells, but also by infection dynamics related to expression of the relevant receptors. There are many data indicating that the receptors for Ad5/3, such as desmoglein 2, are expressed to higher degree on tumor cells than the Ad5 receptor CAR. 17,44,45 Additionally, studies such as those performed by Kawakami et al. indicate that 5/3 chimerism also enhances replication efficiency, 44 thus other than chimeric modifications made to the analyzed virus preparations seem to hamper the previously noted enhanced virion production.
Importantly, a lower physical to functional particle ratio may be advantageous for human use. Adverse events are known to be associated with physical titer, while efficacy is associated with functional titer. For these reasons, physical titering is recommended in clinical trials, as safety is even more important than efficacy. 46
Of note, the titers reported here are not the result of optimization of production. Instead, in most academic laboratories, students, technicians, and other researchers produce viruses themselves, with limited optimization of the process. At CGTG, a panel of quality control assays is performed on each virus batch produced, and batches not fulfilling these criteria are discarded. However, process development has not been the most important issue in preclinical stages and academic processes, since standard methods generally give good adenovirus yields. In contrast, when moving on to clinical trials, it is important to minimize the proportion of inactive particles (which influence adverse events but not efficacy), emphasizing the importance of process development in virus production. Testing the multiplicity of infection, cellular confluency, the cell line used, infection medium, harvest time, and other details can increase yield dramatically. 47
The observed production efficacy was similar when the group of “other capsid modifications” was compared to wild-type-capsid virion preparations. The other capsid modifications group contains several different knob protein modified virions. Thus, these modifications might have altered the stability of the virions in the production system. Although fiber protein modifications of Ad2 have been shown to impact the assembly of the virion, 48 a negative effect of knob protein modifications on virion stability could be less obvious because the knob proteins are located on the outermost parts of the virion. Additionally, several studies have noted a higher infectivity of knob modified Ad5, which would not have been seen if the knob modified virions were very unstable. 17,49
It was hypothesized that when transgenes are inserted into the adenovirus genome, titers could be influenced. Transgene products are produced during virus replication, which might reduce the ability of the cell to produce viral proteins, assuming an overall limited production capacity. Alternatively, the transgene product itself might affect the host's signaling cascades, eventually either hampering or—less likely—enhancing the virion production efficacy. Therefore, the transgenes in this study were grouped and analyzed with regard to virus titers. It was found that the type of insert did not impact the number of produced total virions. However, the production of functional virions was significantly lowered (e.g., immunostimulatory vs. no transgene). Therefore, this indicates that transgenes can affect host-cell production, thus impeding the optimal production and/or folding of virion peptides, the correct assembly of the virion, or some other yet unidentified part of proper virus production.
The effect of the number of modifications was examined in detail. No significant differences were observed in physical and functional titers between preparations of viruses with zero to four modifications. However, the titer was significantly lowered when five modifications were made to virus genomes. This might be caused by a variety of reasons. For example, several signaling sequences (also known as localization peptides) are imbedded into the genome, and the sequence in itself can function as a stabilizing entity. 50 Alternatively or in addition, some other yet unknown functions of the sequence might have been disturbed unknowingly while modifying the genome.
As previous studies by Kennedy and Parks showed that a genome size >105% is undesirable, 7 all virus preparations made in this study were constructed within this rule. Additionally, it has been suggested that smaller genome size correlates with poorer heat stability of adenovirus virions, and if crucial virion proteins are deleted, helper viruses are needed. Thus, this indicates that deleting too much can be counterproductive. However, in the study conducted by Kennedy and Parks, the virions also contained additional modifications, which might have affected results. 7 However, it is maybe not so interesting to make smaller viruses because often the aim is to use the virus to deliver a payload, which requires adding something to the virus.
Thus, it was of interest to see how sensitive virion production was to size changes within the 105% “golden rule.” This analysis revealed that size of the studied virus genomes did not affect the physical or functional virus titers. When adenoviruses are assembled, the electrostatic interactions between DNA and the capsid proteins are neutralized by the basic adenoviral proteins VII, V, and μ, thus probably contributing to the noted flexibility of the adenovirus virions. 51
Since the titer for replication-incompetent and -competent cells are determined in different cell lines (HEK293 and A549 cells, respectively), the intrinsic and non-intrinsic properties of the cell lines might interfere with the results. Therefore, to address this issue, titering data gathered by infection of either A549 or HEK293 cell lines were compared (Supplementary Methods and Results 1, Supplementary Fig. S1). For most viruses, titers were similar, regardless of cell line. Of note, all biological production systems are artificial and thus not perfectly comparable. Specifically, replication-incompetent viruses cannot be analyzed with A549 cells. These might be things to keep in mind when designing quality assays for clinical products.
To assure further that the bias of replication-competent versus -incompetent titration data did not interfere with the results, the analysis was done excluding replication competent data from the data set. In summary, this test showed that when replication-incompetent titration data were deleted from the analysis, the previously noted results were the same or enhanced once when the data were reanalyzed (Supplementary Fig. S2).
With replication-competent viruses, the input titer is just one variable. With each tumor cell producing tens of thousands of new virions, titer may not be the critical determinant of overall efficacy. Instead, when designing viruses for cancer treatment, cellular, intratumoral, and antiviral issues may be more relevant. Also, intratumoral complexities such as extracellular matrix, hyperbaric, necrotic, hypoxic, and acidic areas may thwart spreading of the virus. 52 Of note, the production of big proteins such as antibodies and immunostimulatory proteins seems to be a burden to functional virion production, and thus needs attention from virologists in order to solve this issue. In conclusion, the data collected in this study reveal that adenovirus can be modified quite flexibly without clinically relevant loss of functional titer.
Footnotes
Acknowledgments
This study was supported by Jane and Aatos Erkko foundation, HUCH Research Funds (EVO), Sigrid Juselius Foundation, Finnish Cancer Organizations, the University of Helsinki, and TILT Biotherapeutics Ltd. The University of Helsinki Biostatistics consultation unit (Paula Bergman) is acknowledged for their support in biostatistical conundrums.
Author Disclosure
A.H. is a shareholder in Targovax ASA. A.H. is an employee and shareholder in TILT Biotherapeutics Ltd. R.H., E.H., S.S., M.S., and V.C.-C. are employees of TILT Biotherapeutics Ltd.
Supplementary Material
Supplementary Methods 1
Supplementary Results 1
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.