
Abstract
Timeless is a regulator of molecular clockwork in Drosophila and related to cancer development in mammals. This study aimed to investigate the effect of Timeless on cell proliferation and cisplatin sensitivity in cervical cancer. Timeless expression was determined by bioinformatics analysis, immunohistochemistry, and quantitative polymerase chain reaction (qPCR). Chromatin immunoprecipitation assays and reporter gene assays were applied to determine the transcriptional factor contributing to Timeless upregulation. The effects of Timeless depletion on cell proliferation and cisplatin sensitivity were determined through in vitro and in vivo experiments. Cell apoptosis and senescence were assessed by flow cytometry and β-galactosidase staining. DNA damage and DNA repair pathways were determined by comet assay, immunofluorescent staining, and Western blot analysis. Timeless is aberrantly expressed in ∼52.5% of cervical cancer tissues. E2F1 and E2F4 contribute to the transcriptional activation of Timeless. Timeless depletion inhibits cell proliferation and increases cisplatin sensitivity in vitro and in vivo. Knockdown of Timeless induces cell apoptosis and cell senescence. Mechanically, Timeless silencing leads to DNA damage and impairs the activation of the ATR/CHK1 pathway in response to cisplatin in cervical cancer. Timeless is overexpressed in cervical cancer and regulates cell proliferation and cisplatin sensitivity, presenting an attractive target for cisplatin sensitizer in cervical cancer.
Introduction
Cervical cancer is the fourth leading cause of cancer-related death in women worldwide, and nearly 130,000 new cases are annually diagnosed in China. 1 Although vaccines and screening programs for cervical cancers have been widely implemented, a higher prevalence rate is still found in developing countries. Platinum-based chemotherapies are one of the major strategies for treating advanced cervical cancer. 2 However, the response and survival rate in these patients are still unfavorable. Thus, effective therapeutic targets to improve platinum response in cervical cancer urgently need to be developed.
The Timeless gene is evolutionarily conserved and involved in circadian rhythmicity in Drosophila melanogaster. However, its role in mammals is currently largely elusive. 3 Accumulating reports have demonstrated the involvement of Timeless in cancers, such as colorectal cancer, small cell lung cancer, and breast cancer. 4 –7 These findings suggest that Timeless plays an important role in the development of human cancer. Timeless is also implicated in DNA replication, telomere length, integrity maintenance, 3 and ATM/ATR signaling pathway regulation. 8,9 Recent studies showed that Timeless interacted with poly(ADPribose) polymerase 1 and regulated homologous recombination and nonhomologous end-joining pathways, 10,11 both of which are required for the repair of DNA double-strand breaks (DSBs). 12 However, the role of Timeless in cervical cancer remains elusive.
This study showed that Timeless was aberrantly expressed in >50% of cervical cancer tissues. The depletion of Timeless suppressed tumor cell growth and sensitized cervical cancer cells to cisplatin in vitro and in vivo. Silencing Timeless induced DNA damage and impaired the activation of ATM/CHK2 and ATR/CHK1 signaling pathways in response to cisplatin treatment, suggesting that Timeless could represent a valuable therapeutic target in cervical cancer.
Materials and Methods
Ethics statement
The protocols for handling paraffin-embedded cervical cancer specimens, as well as analyzing patients' data were approved by the Ethics Committee of The First Affiliated Hospital of Soochow University (Suzhou, Jiangsu Province, China). Written informed consent forms were signed by each enrolled patient. All tissue samples were registered by a case number in the database with no patient names or personal information indicated.
Antibodies and reagents
The following primary antibodies were used for Western blot analysis. Timeless (ab109512) was purchased from Abcam (Cambridge, United Kingdom). RAD51 (8875), P21 (2947), cleaved caspase 3 (9661), γH2AX (9718), CHK2 (3440), ATM (2873), phospho-ATM, phospho-CHK2, and phospho-CHK1 were purchased from Cell Signaling Technology (MA). Phospho-ATR (GTX128145) was purchased from Gentex Pharma (WI). GAPDH (M20006) was purchased from Abmart (Shanghai, China). ATR (AF4717) and CHK1 (AF1630) were purchased from R&D Systems (MN). The secondary antibodies used were as follows: horseradish peroxidase-labeled goat anti-mouse IgG (H+L) (A0216), goat anti-abbit IgG (H+L) (A0208), and donkey anti-goat IgG (H+L) (A0181) were purchased from Beyotime Institute of Biotechnology (Shanghai, China). An ATR inhibitor (VE-821) was purchased from Selleck Chemicals (TX).
Cell proliferation and cisplatin sensitivity assay
A total of 4 × 103 cells were seeded in 96-well plates and incubated overnight for attachment. Then, they were transfected with indicated siRNA oligos using lipofectamine. The medium was replaced with a Cell Counting Kit-8 24, 48, 72, 96, and 120 h after transfection and incubated at 37°C for 60 min. Subsequently, the absorbance at 450 nm was measured. In drug sensitivity assay, the cells were treated with indicated doses of cisplatin 48 h after transfection and cultured at 37°C for 24 h; the next steps were the same as mentioned earlier. Triplicate wells were assayed for each condition.
Comet assay
An alkaline comet assay was performed according to the manufacturer's protocol of and OxiSelect™ Comet Assay Kit (STA350) purchased from CELL BIOLABS, Inc. (CA). A 10 μL cell suspension at a concentration of 2 × 105/mL was mixed with 100 μL low-melting agarose and the mixture was layered onto a slide. The slides were immediately placed at 4°C for solidification, transferred to a lysing buffer at 4°C for 1 h, and then transferred to an alkaline solution for 30 min. Next, alkaline electrophoresis was conducted for 15 min at 33 V and ∼300 mA. The slides were stained with Vista Green DNA Dye for 15 min. Finally, 50 comet images were captured for every sample using a fluorescence microscope (Leica Micosystems, Wetzlar, Germany). CASP software was used to analyze the comet images, and the tail moment and olive tail moment were considered the analysis parameters.
Chromatin immunoprecipitation
Chromatin immunoprecipitation (ChIP) assay was performed using an EZ-ChIP Kit (EMD Millipore, MA) according to the manufacturer's protocols. Protein DNA complexes were precipitated with normal IgG, anti-E2F1 antibody (1:100; Cell Signaling Technology), and anti-E2F4 antibody (1:50; Santa Cruz Biotechnology, TX) at 4°C overnight with rotation. Polymerase chain reaction (PCR) was performed with the following primers: E2F1-binding region in the Timeless promoter: sense, 5′-CAATTCTGTACTCTGCTCCT-3′; antisense, 5′-AGGTCTCAACTAGATGCTAAG-3′; E2F4-binding region in the Timeless promoter: sense, 5′-TGTACTCTGCTCCTTAGTCTC-3′; antisense, 5′-GGCACTTTCTGTTCTAATTCAGG-3′; the negative control primer sense, 5′-TGTAGGAGAAGGAGGTTACTA-3′; and upstream antisense, 5′-GAGATAGAAGGTGCTGAGAG-3′.
Tumor growth in nude mice
Female athymic nu/nu mice (aged 6–8 weeks) were used for all in vivo xenograft studies. The mice were quarantined for at least 1 week before experiments. All animal studies were conducted in compliance with the approved institutional Animal Use Protocol. For in vivo cisplatin sensitization assay, female mice were subcutaneously inoculated with Ca Ski cells (1 × 106) at the flank. After 1 week, siRNA was administered intratumorally at a dose of 0.5 OD per injection biweekly. Three days after the first injection of siRNA, cisplatin was administered intraperitoneally (i.p.) at a dose of 2.5 mg/kg every 2 days. For in vivo tumor growth assay, exponentially growing SiHa-shRNA-Ctrl (1 × 106) or SiHa-shTimeless (1 × 106) cells were implanted subcutaneously at the flank of nude mice and grown for 4 weeks. The tumor length and width were measured every 2 days using a digital caliper three times, and the average length and width were used for calculating the tumor volume using the formula (length/2) × (width 2 ).
Statistical analysis
All statistical analyses were carried out using SPSS 20.0 software (IBM, NY). The quantitative values were presented as mean ± standard deviation. The differences between two groups were analyzed using the Student's t-test. GraphPad Prism 7.0 was used for multiple comparison tests. A p-value <0.05 was considered statistically significant (*p < 0.05; **p < 0.01; ***p < 0.001).
More details of experiments are listed in Supplementary Materials and Methods in Supplementary Data.
Results
Timeless is aberrantly expressed in cervical cancer
Previous reports showed that the genes involved in DNA repair might regulate cisplatin sensitivity in several types of cancers.
13
To identify DNA repair genes as potential cisplatin sensitizer in cervical cancer, a web-based meta-analysis was performed using three open-access cervical cancer gene expression array datasets in Oncomine (
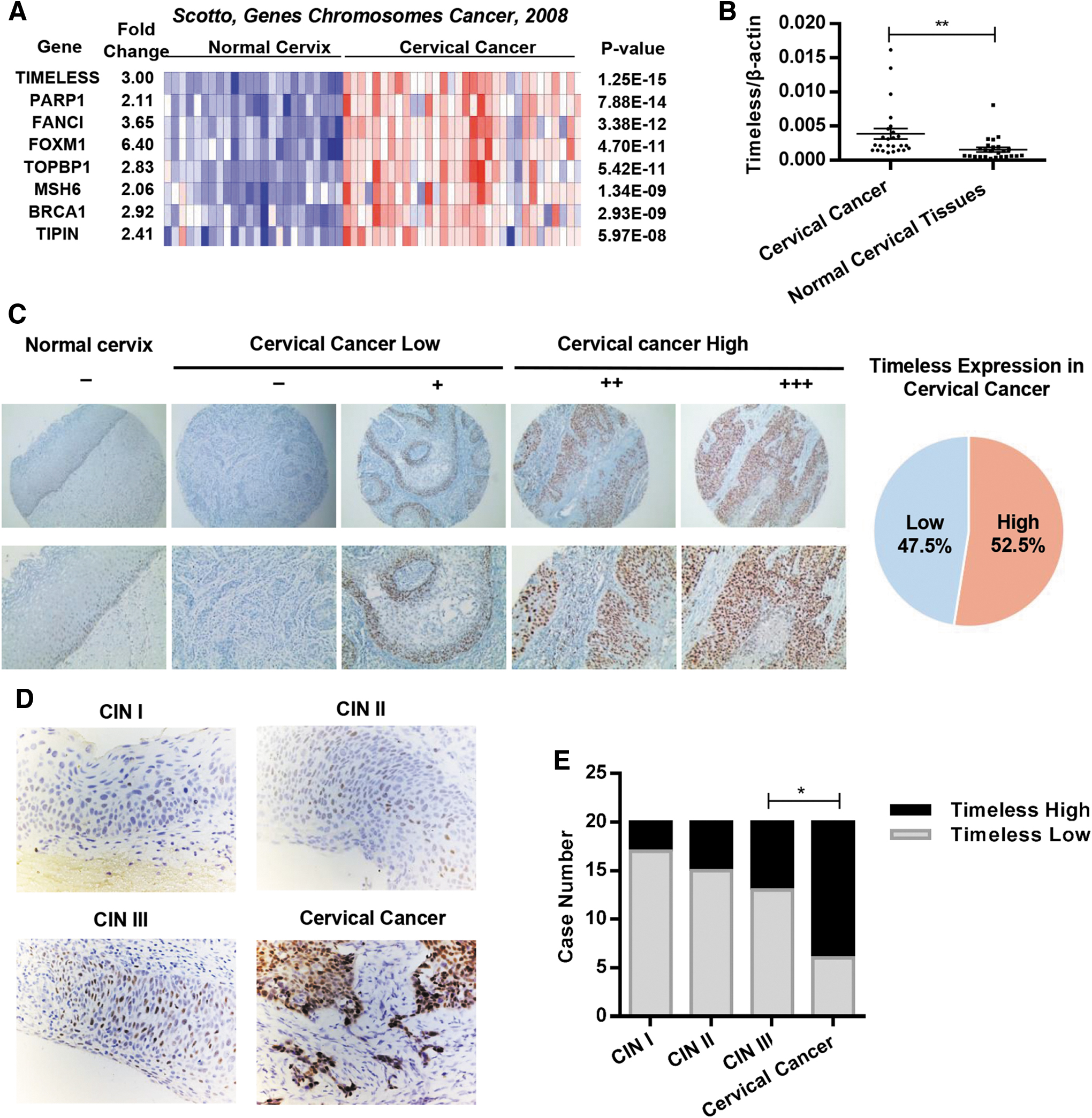
Timeless is aberrantly expressed in human cervical cancer.
E2F1 and E2F4 enhanced the expression of Timeless by directly binding to the Timeless promoter
The mechanism underlying the overexpression of Timeless in cervical cancer was determined by investigating whether any transcription factor is responsible for the increased expression of Timeless in cervical cancer, as no significant copy-number alterations were found on chromosome 12q13.3, in which the Timeless gene was located. 14,15 ChIP-sequencing (ChIP-Seq) data obtained from the University of California Santa Cruz (UCSC) database* suggested that E2F1 and E2F4 both interacted with the Timeless promoter in different regions (Fig. 2A), indicating that E2F1 and E2F4 might contribute to the transcriptional activation of Timeless. A previous study showed that free E2Fs activated gene transcription when the pRB-E2F complex was disrupted by the HPV E7 protein. 16 To assess whether E2F1 or E2F4 promotes Timeless transcription in cervical cancer cells, two pairs of siRNA oligos targeting human E2F1 and E2F4 were transfected into HeLa cells. Both mRNA and protein levels of Timeless significantly decreased in E2F1- or E2F4-depleted cells (Fig. 2B, C), indicating that both E2F1 and E2F4 might affect the expression of Timeless. In addition, ChIP-qPCR assay was performed to validate whether E2F1 and E2F4 bound to specific sites of the Timeless promoter. As expected, the anti-E2F1 and anti-E2F4 antibodies, but not control antibody (IgG), precipitated the Timeless promoter (Fig. 2D). Interestingly, the binding sites of E2F1 (+331 to +343 bp from the transcription initiation site) and E2F4 (+236 to +248 bp) were located in the first intron region of Timeless genome sequence. To further assess the functional role of E2F1- and E2F4-binding regions, site-specific mutagenesis was performed using a wild-type pGL3-Timeless promoter construct (−1500 to +500 bp) (Fig. 2E), in which the mutation of the E2F1- or E2F4-binding region inhibited luciferase activity of the reporter construct, and double mutation led to further inhibition of luciferase activity (Fig. 2F). Taken together, these results suggested that E2F1 and E2F4 bound directly to different sites of the promoter region and activated the transcription of the Timeless gene. Meanwhile, the fact that the reporter remained partially active suggested that other factors might also contribute to the transcriptional activation of Timeless.
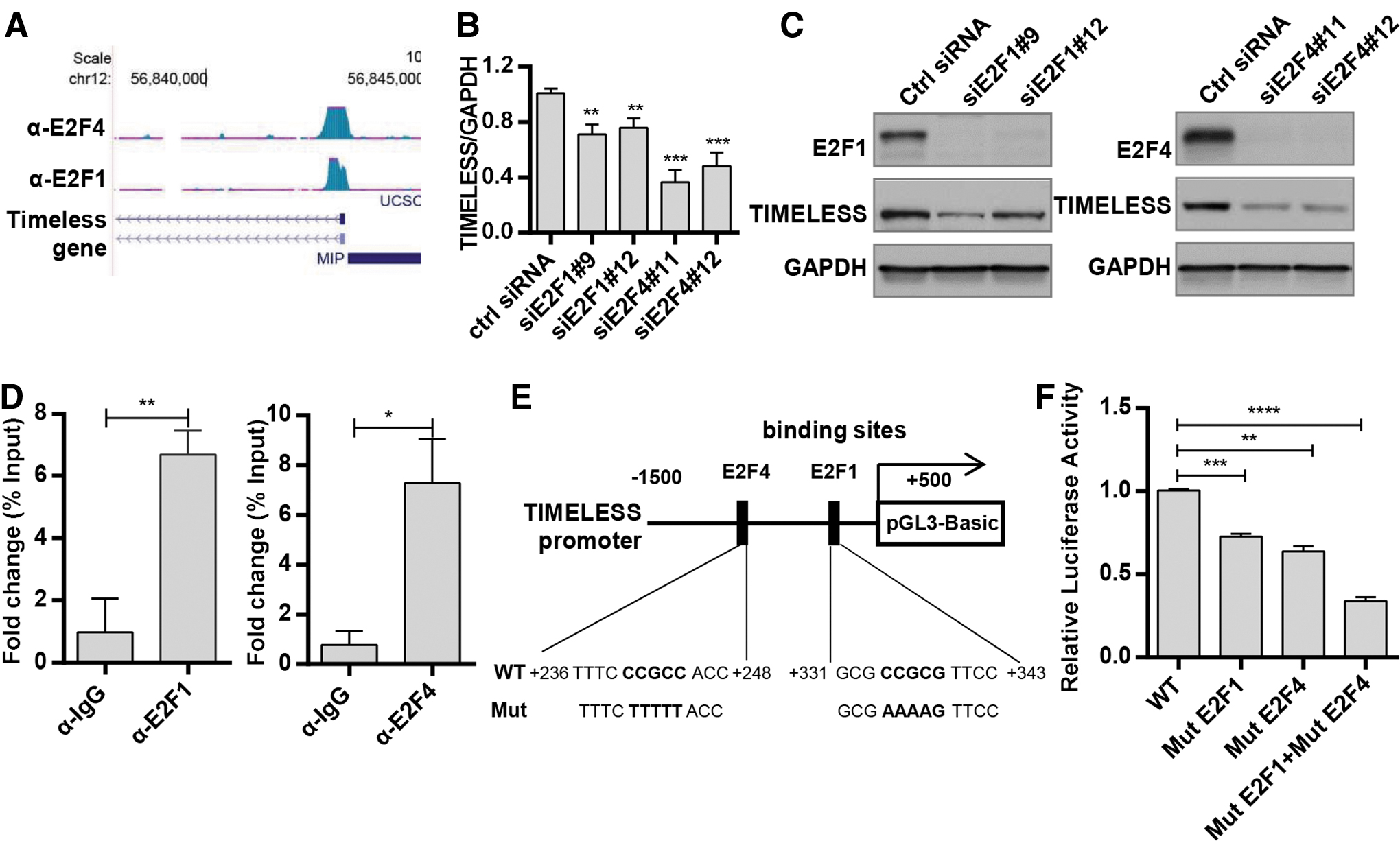
E2F1/E2F4 enhances Timeless expression by directly binding to Timeless promoter.
Loss of Timeless inhibited cervical cancer cell proliferation, and induced cell apoptosis and senescence
As Timeless is overexpressed in cervical cancer, this study investigated whether Timeless contributed to cervical cancer progression. It first verified that Timeless was expressed in four cervical cancer cell lines: SiHa, HeLa, Ca Ski, and C33A (Supplementary Fig. S2A). Then, three different small interfering RNAs (siRNAs) were applied to silence the expression of Timeless, and the knockdown efficiency was checked by Western blot analysis (Fig. 3A and Supplementary Fig. S2B). Tumor cell proliferation was significantly inhibited in SiHa, Ca Ski, and HeLa cells transfected with siRNAs targeting Timeless (Fig. 3B and Supplementary Fig. S2C). In addition, the knockdown of Timeless also impaired clone formation ability in three cervical cancer cell lines (Fig. 3C and Supplementary Fig. S2D). Next, the study investigated whether the knockdown of Timeless led to apoptosis or senescence in cervical cancer cells. Cell apoptosis was assessed at 72 h after siRNA transfection by flow cytometry (Fig. 3D and Supplementary Fig. S2E). The quantitative analysis revealed that the knockdown of Timeless promoted apoptosis in SiHa, Ca Ski, and HeLa cells (Fig. 3E and Supplementary Fig. S2F). Furthermore, Timeless depletion also led to a significant increase in β-galactosidase staining in SiHa cells (Fig. 3F, G), indicating increased cell senescence. In accordance with aforementioned findings, the knockdown of Timeless resulted in the elevated expression of cleaved caspase-3 and p21 (Fig. 3H–J and Supplementary Fig. S2G, H), indicating that the depletion of Timeless led to cell apoptosis and senescence in cervical cancer.

Loss of Timeless inhibited cervical cancer cell proliferation and induced cell apoptosis and senescence.
Timeless depletion sensitized cervical cancer cells to cisplatin in vitro
Previous studies showed that the genes involved in the DNA repair pathway, such as ERCC1, might regulate cisplatin sensitivity and affect patient prognosis. 17,18 They also demonstrated that Timeless was involved in replication fork stability and DNA damage repair. 10 This study assessed whether Timeless depletion could sensitize cervical cancer to cisplatin. The cervical cancer cells were treated with cisplatin after Timeless siRNA transfection, and the cellular viability was detected 24 h after the treatment. The loss of Timeless in SiHa, Ca Ski, and HeLa cells produced a >1.5-fold change in IC50 (Fig. 4A–C). The changes in the IC50 value in three cell lines were all significant (Supplementary Fig. S3A–C), indicating the cisplatin sensitization effect of Timeless depletion in cervical cancer cells.

Timeless knockdown sensitized cervical cancer to cisplatin in vitro. Cells were plated at a density of 5,000 cells in a 96-well plate and transfected with indicated siRNAs; 24 h later, cells were treated with increased concentration of cisplatin for another 48 h. Viability of cells was measured with a CCK8 assay. The dose–response curve of SiHa, CaSki, and HeLa cells was plotted in
Then, cell apoptosis and senescence were measured after the combined treatment of siRNA and cisplatin. As expected, more apoptotic cells were found following cisplatin treatment in Timeless downregulated clones versus control (Fig. 4D, E and Supplementary Fig. S3D, E). In addition, Timeless siRNA transfection also led to a significant increase in cell senescence after cisplatin treatment compared with control siRNA (Fig. 4F, G). These data indicated that Timeless depletion might enhance cisplatin efficacy by inducing cell apoptosis and senescence.
Silencing Timeless induced DNA damage and impaired ATR/CHK1 signaling in cervical cancer cells
Next, the study examined whether the sensitization effect of Timeless was associated with the DNA-damage-repair pathway alteration in cervical cancer. First, single-cell gel electrophoresis assay 19 was applied to detect the effect of Timeless on DNA damage in cervical cancer. As shown in Fig. 5A and B, Timeless-depleted cells showed increased tail formation, indicating a higher level of DNA damage. Immunofluorescence staining showed that both Timeless siRNAs resulted in significantly increased γH2AX foci (Fig. 5C). Immunoblotting also suggested that the knockdown of Timeless led to an increased level of γH2AX (Fig. 5D and Supplementary Fig. S4A). In addition, the expression of RAD51, a protein that plays a major role in the homologous recombination of DNA during DSB repair, 20 was also significantly inhibited in cervical cancer cells (Fig. 5D and Supplementary Fig. S4A). Then, the study determined whether exogenous Timeless expression rescued Timeless siRNA-induced DNA damage. To avoid the effect of siRNA on exogenous Timeless expression, co-transfection was performed using the pIRES2-EGFP-Timeless plasmid and siRNAs targeting the 3′-UTR region of the Timeless gene (siTIM-UTR-216/271), which were tested for silencing efficiency (Supplementary Fig. S4B, C). The downregulation of Timeless and increase in the expression of γH2AX by silencing Timeless were both reversed by exogenous Timeless expression (Supplementary Fig. S4C, D, E), indicating that Timeless silencing led to DNA damage in cervical cancer cells.

Targeting Timeless induced DNA damage and impaired ATR signaling pathway.
In response to cisplatin-induced DNA crosslinks and adducts, rapid DNA damage response (DDR) was observed, which was mediated by the ATM/ATR signaling pathway, as well as by two downstream kinases, checkpoint kinase 1 and 2 (CHK1 and CHK2). 21 Several inhibitors targeting the ATM/ATR pathway are under preclinical or clinical trial evaluating anticancer efficiency. 22 This study examined whether the activation of ATM or ATR pathway was impaired in Timeless-depleted cells treated with cisplatin. As illustrated in Fig. 5E, in cells with Timeless depletion, ATR/CHK1 phosphorylation was significantly attenuated after cisplatin treatment; however, ATM/CHK2 phosphorylation was almost not affected. These data indicated that the ATR/CHK1, but not ATM/CHK2 pathway, was impaired in cells with Timeless silencing after cisplatin treatment. Besides, Rad51 protein was also downregulated in Timeless-silenced cells after cisplatin treatment (Fig. 5E). These data suggested that Timeless played a key role in ATR/CHK1 signal activation, and Timeless deficiency impaired ATR-mediated signaling in response to cisplatin-induced DNA damage. To further determine whether the sensitization effect of Timeless depended on the attenuated ATR pathway, the ATR inhibitor VE-821 was applied to inhibit ATR activity, and the IC50 value was also assessed. As shown in Supplementary Fig. S4F, ATR inhibition mimicked the sensitization effect of Timeless silencing. More importantly, Timeless depletion did not further sensitize cells to cisplatin after ATR inhibition (Supplementary Fig. S4F), indicating that the sensitization effect of Timeless at least partially depended on ATR inhibition.
Targeting Timeless attenuated tumor growth and sensitized cervical cancer cells to cisplatin in a xenograft model
The cisplatin sensitization effect of Timeless depletion in cervical cancer was further confirmed using cell lines derived from a xenograft model. The flow diagram of the in vivo experiment is shown in Fig. 6A. The expression of Timeless was silenced by the intratumoral injection of siRNA, as evidenced by Western blot analysis of tumor lysates after the first injection of siRNA (Fig. 6B). In accordance with in vitro data, decreased Rad51 expression and increased H2AX expression were also detected in tumors after the first injection of Timeless siRNA (Fig. 6B). As shown in Fig. 6C–E, the knockdown of Timeless resulted in a significant decrease in tumor growth after cisplatin treatment compared with nontargeting control siRNA treated with cisplatin. In addition, the depletion of Timeless, independent of cisplatin, also resulted in a modest, but significant, decrease in tumor growth in Ca Ski xenografts (Fig. 6C–E). Knockdown of Timeless also inhibited the tumor growth in SiHa xenografts (Supplementary Fig. S5B). Taken together, these results confirmed that cervical cancer cells with Timeless depletion showed increased sensitivity to cisplatin in vivo.
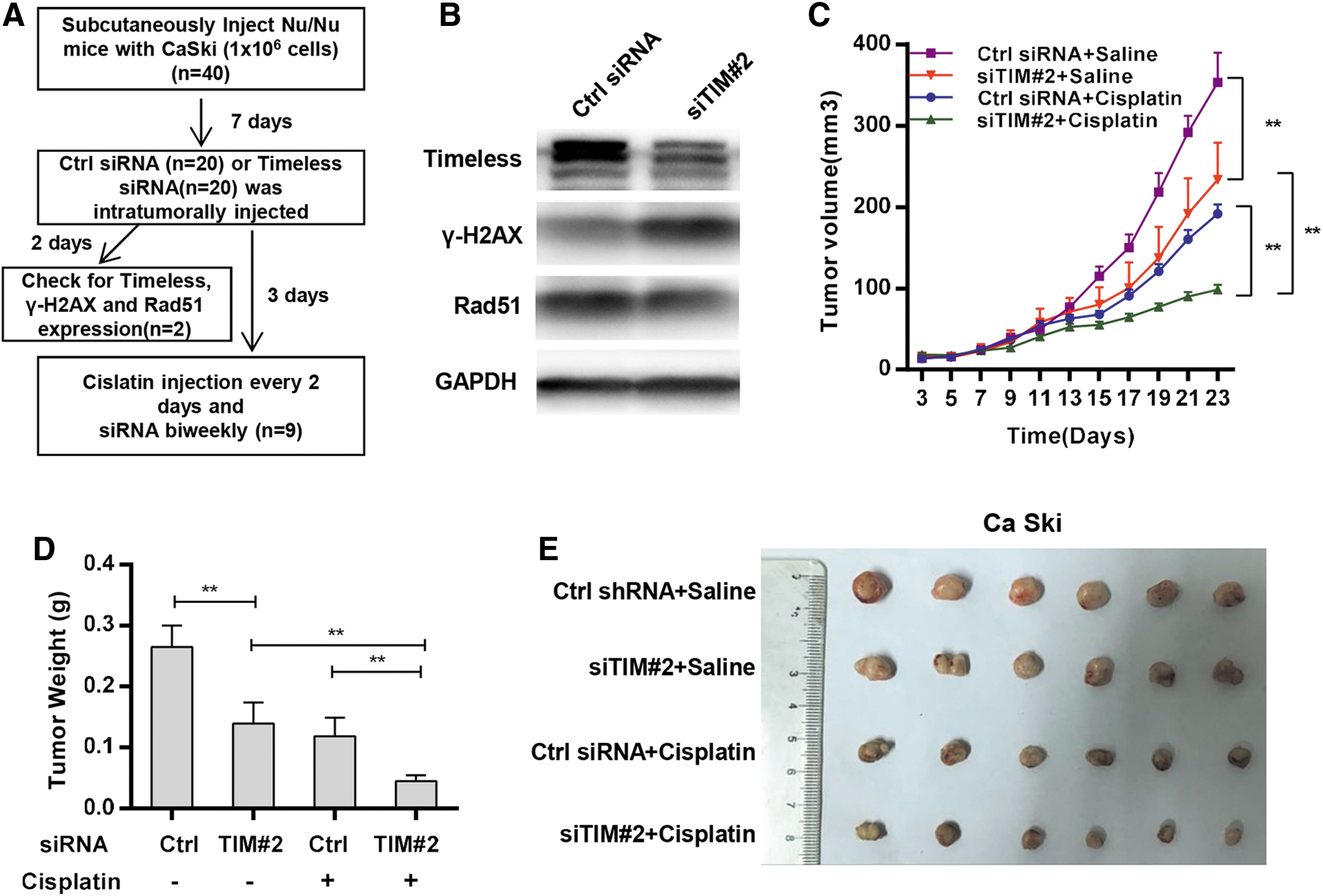
Targeting Timeless attenuated tumor growth and sensitized cervical cancer cells to cisplatin in xenograft model.
Discussion
Aberrant expression of Timeless has been reported in several types of cancers. 6,23 –25 However, the role of Timeless in cervical carcinoma has remained unclear. This study verified that Timeless was frequently overexpressed in cervical cancer tissues. In addition, it also showed that E2F1 and E2F4 contributed to Timeless overexpression by directly activating gene transcription in cervical cancer.
The phenotype analysis supported a protumorigenic function of Timeless because its depletion resulted in the deceased viability of cervical cancer cells. Three reasons may justify this phenotypic change. First, Timeless acts as a replisome-associated protein, 26 which is implicated in DNA synthesis, S-phase-dependent checkpoint activation, and chromosome cohesion. 27,28 Its depletion gave rise to a defective mitotic progression, 29 resulting in lower rates of cell proliferation. Second, it also might be partially attributable to the promotion of apoptosis. Timeless-deleted cells displayed an elevated level of apoptosis, along with the generation of cleaved caspase-3. Eventually, knocking down of Timeless might induce the aging of cervical cancer cells since Timeless-depleted cells expressed more senescence-associated β-galactosidase. This was also proved by the increased expression of p21, a canonical marker of cell aging. Together, the results indicated that Timeless depletion impaired the viability of cervical cancers.
Timeless silencing results in significant DNA damage accumulation in cervical cancer cells, as evidenced by increased H2AX phosphorylation (γH2AX) in immunoblotting assay and tail formation in comet assay. This damage accumulation may be ascribed to increased DNA damage or impaired DNA repair, or both. Oncogene-induced replication stress is one of the hallmarks of cancer cells, and leads to cell senescence under some circumstances. 30 Timeless is a member of replication fork complex that confers to fork stabilization and prevents the collapse of the stalled replication fork. Timeless dysfunction may result in the accumulation of stalled or collapsed replication fork and DNA damage. Timeless has been shown to recruit at the replication fork and interact with the checkpoint kinase CHEK1 and the ATR-ATRIP complex. It is involved in ATM-/CHEK2-mediated signaling, suggesting that Timeless might play an important role in DDR. 31,32
Platinum-based chemotherapy is one of the main methods for treating advanced cervical cancer, 33 by inducing DNA crosslinks and replication stress. The results showed that targeting Timeless sensitized cervical cancer cells to cisplatin in vitro and in vivo. Although cisplatin induces several types of DNA damage, whether it can directly produce double-strand DNA breaks is still unclear. 34 Cisplatin-induced obstructed replication forks can be converted into replication-associated DSBs, which are repaired by homologous recombination. 35 When DSBs occur, ATM/ATR phosphorylates the histone H2A variant H2AX, which can spread thousands of base pairs around DSB sites and provide docking sites to recruit DNA damage-responsive sensors. The recruitment of these sensor proteins further activates or maintains ATM kinase activity and amplifies ATM signaling. 36,37 In cisplatin-treated cervical cancer cells, Timeless depletion led to the impaired activation of the ATR/CHK1 pathway, indicating that Timeless might act upstream of the ATR/CHK1 pathway in the DNA damage repair process in cervical cancer.
The decreased RAD51 expression may also contribute to the cisplatin sensitization effect of Timeless depletion. Decreased RAD51 might induce noneffective homologous recombination repair and decreased γH2AX, thereby causing cells to undergo mitotic catastrophe or apoptosis, and sensitize cells to cisplatin treatment. The specific mechanism of the regulation of RAD51 by Timeless in cervical cancer needs further investigation. This mechanism of sensitivity to cisplatin is similar to that in previous studies, which reported that RAD51C-deficient irs3 cells were more sensitive to killing by cisplatin compared with the parental V79-4 cell line, and XPF-deficient UV41 cells were also more sensitive than their parental AA8 cell line to cisplatin. 34,38
In clinical settings, patients with the FIGO stage IB-IIA disease have a recurrence risk ranging from 10% to 20% despite co-current chemoradiation, while patients with stage IIB-IVA disease have a 50–70% chance of recurrence. 39 Both cisplatin and radiation interfere with the DNA damage repair process by disturbing the progression of replication forks. Therefore, targeting Timeless in conjunction with chemotherapy may make it possible to decrease the doses of chemotherapy agents and hence minimize side effects. Collectively, the results suggested that Timeless might play important roles in DNA damage repair and provide a candidate therapeutic target in cervical cancer.
The identification of novel therapeutic targets has still remained a critical issue for improving the survival of patients with cervical cancer. This study demonstrated that Timeless, a factor involved in DNA repair, was overexpressed in cervical cancer tissues and was essential for tumor cell proliferation. Its deficiency inhibited tumor cell proliferation and sensitized cervical cancer cells to cisplatin, promoted apoptosis and senescence, and impaired DNA repair pathway in cervical cancer. According to the renewal of interest in the targeting of DDR as a strategy for antitumor therapy, the findings suggested that the silencing of Timeless in conjunction with cisplatin could potentially be an effective treatment strategy for advanced cervical cancer.
Footnotes
Author Disclosure
No competing financial interests exist.
Funding Information
This work was supported by the National Natural Science Foundation of China (Grant Nos. 81772773, 81672560, and 81302275), The Jiangsu Provincial Medical Youth Talent (Grant No. QNRC2016753), the Project of Jiangsu Provincial Maternal and Child Health Association (Grant No. FYX201709), the Project from Jiangsu Key Discipline of Human Assisted Reproduction Medicine Foundation (FXK201749), and Jiangsu Provincial Medical Talent of the Project of Invigorating Healthcare through Science, Technology and Education (ZDRCA2016044).
Supplementary Material
Supplementary Table S1
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.