
Abstract
Chemotherapy causes inflammation, which promotes cancer development and results in complications such as hemorrhages and thrombosis. Development of new therapeutic strategies to limit inflammatory responses will potentially reduce these side effects in cancer patients. Gene therapy is an attractive cancer treatment because of its high specificity and limited side effects. A tumor suppressor gene associated with retinoid-interferon–induced mortality-19 (GRIM-19) was delivered by an amphiphilic copolymer poly(ɛ-caprolactone) and ethanolamine-functionalized poly (glycidyl methacrylate) (PCG). The transfection outcome of PCG/pGRIM-19 complexes was studied both in vitro and in vivo. The antitumor therapeutic effects were evaluated in a well-vascularized Neuro-2a neuroblastoma tumor mouse model in comparison with that of cisplatin. The PCG/pGRIM-19 nanocomplex showed high transfection efficiency and low toxicity. In vitro transfection of PCG/pGRIM-19 in Neuro-2a cells reduced the expression levels of Cyclin D1, BCL-2, and MMPs 2 and 9, and inhibited cell proliferation, migration, and stimulated apoptosis. In vivo experiments indicated that PCG/pGRIM-19 therapy downregulated signal transducer activator of transcription 3, nuclear factor-κB, and MMP9 expression in tumor tissues. Compared with cisplatin treatment, gene therapy with PCG/pGRIM-19 significantly inhibited local complications of intratumor hemorrhages, and systemic complications such as anemia and pulmonary embolism, thereby effectively prolonged mouse survival. Our results highlight the potential of PCG/pGRIM-19 gene therapy in reducing tumor burden and complications, providing novel strategies to enhance clinical cancer therapy.
Introduction
The complexity of cancer poses many challenges to treatment. 1 For example, inflammation, a fundamental innate immune response to perturbed tissue homeostasis, 2 is a recognized hallmark of cancer that affects all stages of cancer development and progression. 3 Besides promoting tumorigenesis and metastasis, 2,4 inflammation also causes endothelial dysfunction, which promotes intratumor hemorrhages 5 and systemic complications such as anemia and thrombosis. 6 These complications increase the difficulty of cancer treatment. Unfortunately, chemotherapeutic drugs injure the endothelium, which aggravate complications caused by endothelial dysfunction. 7,8 Development of new therapeutic strategies to limit inflammatory responses will potentially reduce these side effects in cancer patients. 3
Gene therapy has attracted great interest in cancer treatment because of its high efficacy and limited side effects. 9 It maximizes therapeutic advantages over conventional cytotoxic chemotherapy by modulating gene expression in cancer cells. 10 Gene associated with retinoid-interferon–induced mortality-19 (GRIM-19) is a cancer suppressor gene that inhibits signal transducer activator of transcription 3 (STAT3) and nuclear factor-κB (NF-κB) signaling pathways. 11,12 GRIM-19 restrains the malignant behavior of cancer cells by regulating proliferation- and invasion-associated genes, such as CCND1, BCL-2 family, and MMPs. 12 Overexpression of GRIM-19 in cancer cells has been reported to suppress STAT3-mediated signal transduction and cancer growth, 13 which makes it a potential target for cancer gene therapy. As STAT3 and NF-κB are involved in two of the major cancer-associated inflammatory pathways, 2 inhibition of these two transcription factors would also inhibit inflammation while suppress tumor growth.
Herein, we explored the therapeutic potential of exogenous delivered GRIM-19 gene in cancer treatment, especially in well-vascularized tumor with intratumoral hemorrhage. A cationic polymeric nanoparticle (NP) composed of ethanolamine (EA)-functionalized poly (glycidyl methacrylate) (PGEA) and polycaprolactone (PCL) was used as the delivery vehicle for plasmid DNA (pDNA) that encodes GRIM-19. PGEA-based vectors, with abundant hydroxyl groups shielding excessive positive charges, are promising gene carriers with high transfection efficiency and low cytotoxicity. 14 –16 Combining hydrophobic components with polycation derivatives could further enhance transgene efficiency and biocompatibility. 17 –19 PCL–PGEA (PCG) block copolymers have shown to mediate significantly enhanced gene transfection efficiency compared with polyethylenimine (PEI) but with lower cytotoxicity. 20 In this study, we evaluated the therapeutic effect of PCL–PGEA NP-mediated GRIM-19 gene delivery in well-vascularized Neuro-2a tumors with intratumor hemorrhages. Local and systemic complications from inflammation and endothelial injury were comprehensively studied in tumor-bearing mice. The effects of gene therapy on survival rate and cancer-related complications in vivo were compared with that of conventional chemotherapy.
Materials and Methods
Materials
PEI (Mw 25 kDa), ɛ-caprolactone (ɛ-CL), glycidyl methacrylate (GMA), Hochest33258, cisplatin, Drabkin's reagent, and Brij 35 Solution 30% were purchased from Sigma-Aldrich (St Louis, MO). EA was purchased from Guangfu Fine Chemical Research Institute (Tianjin, China). Deuterohemin-b-Ala-His-Thr-Val-Glu-Lys (DhHP-6) was provided by the College of Life Science, Jilin University (Changchun, China). GRIM-19-encoded plasmid pCDNA3.1-GRIM-19, empty plasmid pCDNA3.1, and pEGFP-N1 were kindly provided by the Basic Medical College, Jilin University (Changchun, China). Trypsin, Dulbecco's modified eagle medium (DMEM) was purchased from Hyclone (Logan, UT). Fetal bovine serum (FBS) was purchased from Bioind (Kibbutz Beit-Haemek, Israel). Phosphate-buffered saline (PBS) was obtained from Sangon Biotech (Shanghai, China). Penicillin–streptomycin solution, phenylmethanesulfonyl fluoride (PMSF), and paraformaldehyde were obtained from Solarbio (Beijing, China). RIPA lysis buffer was obtained from Beyotime Biotechnology (Shanghai, China). TRIzol lysis buffer was purchased from Invitrogen (Carlsbad, California). Diethyl pyrocarbonate-treated water was obtained from Coolaber (Beijing, China). PCR MIX and Super RT Kit were purchased from BioTeke (Beijing, China). Cell Counting Kit-8 (CCK-8) was purchased from MCE (New Jersey). Annexin V FITC Apop Dtec Kit was obtained from BD (Franklin Lakes, NJ). Transwell chamber was purchased from Costar (New York). The antiproliferating cell nuclear antigen (PCNA) (sc-56) was obtained from Santa Cruz (Newark, DE). Cyclin D1 (catalog no.: A11022) was purchased from Abclonal (Boston, Massachusetts). Anti-GRIM-19 (catalog no.: 10986), anti-STAT3 (catalog no.: 10253), anti-NF-κB (catalog no.: 10745), anti-β-actin (catalog no.: 66009), anti-Bcl-2 (catalog no.: 12789), anti-MMPs 2 (catalog no.: 10373), and anti-MMPs 9 (catalog no.: 10375) antibodies were purchased from Proteintech (Chicago, Illinois).
Preparation and characterizations of PCG
The PCL-b-PGMA block copolymer was first synthesized through the combination of enzymatic ring-opening polymerization and enzymatic atom transfer radical polymerization (ATRP), following the previously reported procedure. 20 In brief, PCL-Br was prepared through HEBiB (300 mg)-initiated polymerization of ɛ-CL (3 mL) using Novozym435 (150 mg) as the catalyst. Then, PCL-b-PGMA was synthesized through DhHP-6 (1.8 mg)-catalyzed ATRP of GMA (0.6169 g) using the PCL-Br (Mn = 2,400; PDI = 1.22) as the initiator at 30°C for the desired reaction time. Finally, the cationic block copolymer PCG was synthesized through ring-opening reactions between PCL-b-PGMA and EA at 60°C for 7 h.
The chemical structure of PCG was characterized by nuclear magnetic resonance (NMR) spectroscopy. 1 H NMR spectra were measured on Avance III NMR spectrometer (Bruker, Germany) in CDCl3, in which chemical shifts (in ppm) were recorded using tetramethylsilane as the internal standard.
For the preparation of PCG/DNA complexes, PCG was dissolved in PBS to achieve a concentration of 10 mg/mL and stored at 4°C. Polymer to DNA ratios are expressed as mass ratios. At the desired W/W ratios, equal volumes of polymer and DNA solutions were mixed, vortexed, and incubated for 30 min at room temperature to produce polymer/DNA complexes.
Dynamic light scattering assay
The particle sizes and surface zeta potentials of the PCG/pGRIM-19 complexes at series W/W ratios of 1–50 were measured using a Zetasizer Nano ZS90 (Malvern Instruments, Worcestershire, United Kingdom) with a laser wavelength of 633 nm at a scattering angle of 173°.
Agarose gel electrophoresis assay
pGRIM-19 (1.0 μg) was mixed gently with PCG at various W/W ratios in a total volume of 10 μL and incubated for 30 min at room temperature. The binding ability of PCG to DNA was detected by electrophoresis in 1% agarose with TAE buffer (120 V, 20 min). The gel was assessed by a Tannon 2500R image analysis system (Shanghai, China).
Cell culture and transfection
Human hepatoma cell SMMC-7721, human neuroblastoma cell SY5Y, and murine neuroblastoma Neuro-2a cell were obtained from the Chinese Academy of Sciences Cell bank. All cells were maintained in a high glucose DMEM supplemented with 10% FBS and 1% penicillin–streptomycin solution at 37°C in 5% CO2. Before gene transfection, cells were seeded in six-well plates at a density of 5.0 × 105 cells per well overnight. Each 4.0 μg pDNA sample was mixed with polymer and added to wells with serum-free medium. The medium was changed 6 h later. After culturing for 24 h, the transfection efficiency was measured using an IX71 fluorescence microscope (Olympus, Shinjuku, Japan).
Western blot analysis and reverse transcription polymerase chain reaction
Four micrograms of pDNA samples was mixed with PCG (at W/W ratio = 20) or PEI (at W/W ratio = 1.3) for transfection in six-well plates. Total cellular proteins were extracted in RIPA lysis buffer with 1 mM PMSF. After protein quantification, the samples were separated using 12% sodium dodecyl sulphate-polyacrylamide gel electrophoresis and transferred onto polyvinylidene fluoride membranes. After blocking with 5% skim milk, the membranes were incubated with primary antibodies and secondary antibody and were then detected by Syngene Bio Imaging (Synoptics, Cambridge, United Kingdom).
Total cellular RNA was isolated using TRIzol. After quantification, 2 μg of total RNA samples was used for synthesizing first-strand cDNA with a Super RT Kit. Primers used in reverse transcription polymerase chain reaction were as follows: β-actin sense, 5′-GTGACGTTGACATCCGTAAAGA-3′ and antisense, 5′-GCCGGACTCATCGTACTCC-3′; GRIM-19 sense, 5′-GTCGAAGGTGAAGCAGGACA-3′ and antisense, 5′-CATGATGATGGCTTCCTCCTC-3′. The products were separated using 1% agarose gel electrophoresis and visualized using a Tanon imaging system (Shanghai, China).
In vitro cytotoxicity assay
The effect of PCG/pGRIM-19 complexes on cell proliferation was examined by CCK-8 and colony formation assays. For the CCK-8 assay, cells were seeded in 96-well plates at a density of 3.0 × 103 cells per well. A total of 0.2 μg of pDNA was mixed with PCG (at W/W ratio = 20) or PEI (at W/W ratio = 1.3) for transfection. After transfection for 48 h, CCK-8 was added and incubated for 1 h. The OD450 was measured using a FLUOstar Omega reader (BMG LABTECH, Ortenberg, Germany). For the colony formation assay, 24 h after transfection, 1,000 cells per well were reseeded in 24-well plates. One week later, the colonies were fixed and dyed with 0.2% crystal violet.
The effect of PCG/pGRIM-19 complexes on cell apoptosis was assayed by flow cytometry and Hochest33258 staining. For the flow cytometry, 4 μg of pDNA samples was mixed with PCG (at W/W ratio = 20) or PEI (at W/W ratio = 1.3) for transfection in six-well plates. Then, 48 h later, the treated cells were collected and stained with an Annexin V FITC Kit before detection using an Accuri C6 flow cytometer (Becton Dickinson, Franklin Lakes, NJ). For the Hochest33258 staining, cells were seeded on sterile glass coverslips in 24-well plates at a density of 5 × 104 per well. Then, 0.8 μg of pDNA was mixed with PCG (at W/W ratio = 20) or PEI (at W/W ratio = 1.3) for transfection. Forty-eight hours after transfection, cells were stained with Hochest33258 and observed with a BX53 fluorescence microscope (Olympus).
The effect of PCG/pGRIM-19 complexes on cell migration was analyzed by wound-healing and cell migration assay. For the wound-healing assay, cells were seeded in 12-well plates and achieved 90% confluence. Then, 1.6 μg of pDNA samples was mixed with PCG (at W/W ratio = 20) or PEI (at W/W ratio = 1.3) for transfection. A scratch wound was produced with a sterile pipette tip, and cells were washed twice with medium. After transfection for 24 h, the changes in the cell-covered area were captured with a microscope and analyzed with ImageJ software. For the cell migration assay, transfected cells were seeded into the upper Transwell chamber in 200 μL of serum-free DMEM, whereas 600 μL of DMEM with 10% FBS was added to the lower chamber. After 24 h of culture, the migrating cells on the lower surface of the membrane were stained with 0.2% crystal violet for 20 min and photographed.
In vivo experiments
Neuro-2a neuroblastoma graft in 4-week-old female nude BALB/c mice (Beijing Huafukang Bioscience Co., Inc., China) was established to assess the in vivo antitumor efficiency of pGRIM-19 transfection mediated by PCG. In brief, 1 × 106 cells in 100 μL DMEM were injected into the subcutaneous space of nude mice. Treatment began when the subcutaneous tumors reached 100–200 mm3, which was ∼5 days after implantation. The mice were randomly divided into three groups: group 1, blank control (only PBS); group 2, PEI/pGRIM-19 complexes containing 30 μg of pGRIM-19 (W/W ratio = 1.3); and group 3, PCG/pGRIM-19 complexes containing 30 μg of pGRIM-19 (W/W ratio = 20). The volume of the intratumoral injection was maintained at 100 μL, and all the treatments were, respectively, performed once every other day for 8 days. Tumor size was measured using calipers and tumor volume (mm3) was calculated as (L × W 2 )/2, where L = length (mm) and W = width (mm).
The tumors were imaged, weighed, and dissected before hematoxylin-eosin staining and immunohistochemical analyses. Immunohistochemistry results were quantified using ImageJ software. The lungs were also treated by hematoxylin-eosin staining to assess the severity of pulmonary embolism.
To determine the effect of PCG/pGRIM-19 complexes on survival of tumor-bearing mice, we used cisplatin treatment as a positive control (2 mg/kg, twice a week for 2 weeks, intraperitoneal injection, n = 5 21 ) and counted the days from the initiation of implantation to the time the tumors reached a diameter of 2 cm (end point as required by NIH Animal Care and Use Committee). For hematocrit detection, blood was collected into EDTAK2 tubes and analyzed using an auto-analyzer (XS-1000i, Sysmex, Japan).
Animal welfare and experimental procedures were carried out in accordance with EU Directive 2010/63/EU for animal experiments, and were approved by the Institutional Review Board of the College of Basic Medical Sciences, Jilin University.
Statistics
Data are expressed as the mean ± standard deviation. The differences between two groups were calculated by using the Student's t-test. If more than two groups were compared, evaluation of significance was performed using one-way analysis of variance (ANOVA) followed by Bonferroni's post hoc test. Survival curves were compared using the log-rank Mantel–Cox test. p < 0.05 was used as the criterion of statistical significance. All analyses were performed with GraphPad Prism (Version 6; GraphPad Software, Inc.).
Results
Synthesis and characterization of PCG/pGRIM-19 NPs
The cationic block copolymer PCG was synthesized through the combination of enzymatic ATRP and enzymatic ring-opening polymerization, as shown in Fig. 1A. These amphiphilic block copolymers are able to self-assemble into NPs in aqueous solutions, with a hydrophobic PCL core and a cationic PGEA shell to interact with the DNA of interest. The typical 1 H NMR spectrum of PCG is shown in Fig. 1B. A PCG polymer with a degree of polymerization of 142 for the PGEA segments (Mn = 2.3 × 104 g/mol with polydispersity index of 1.4) was selected as the gene carrier in this study due to its higher performance compared with its lower molecular weight counterparts. 20
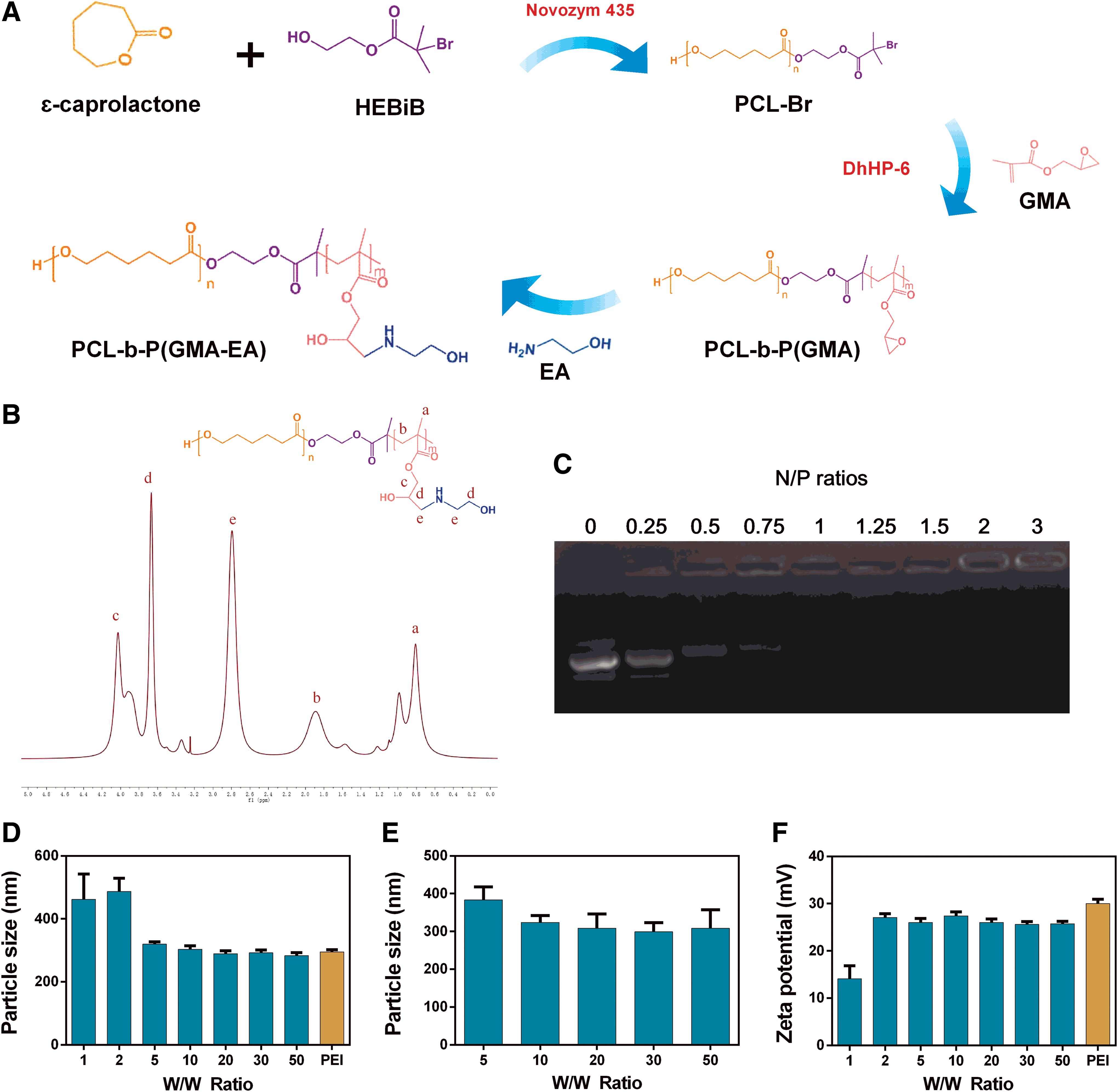
Characterization of PCG/pGRIM-19 nanoparticles.
The ability of PCG to condense pDNA was evaluated by agarose gel electrophoresis and dynamic light scattering. PCG completely condensed pDNA at N/P = 1 (Fig. 1C). Similar to the “gold-standard” PEI (25 kDa), PCG compressed pDNA into suitable particle sizes for cellular uptake, which ranged from 200 to 300 nm at the W/W ratios of 5 to 50 (Fig. 1D). The PCG/pDNA complexes maintained particle sizes within 400 nm in 10% serum. (Fig. 1E). The zeta potentials of the PCG/pDNA and PEI/pDNA complexes were also comparable, varying from 25 to 30 mV (Fig. 1F).
Transfection efficiency and cytotoxicity of PCG/DNA complexes in vitro
The transfection efficiency of the PCG vector was first evaluated using pEGFP-N1 plasmid as the reporter gene in Neuro-2a cells. As shown in Fig. 2A and B, PCG mediated green fluorescent protein (GFP) expression in >50% of the transfected cells at the weight ratio of 20, which is similar or higher than the expression mediated by PEI 25k. Cell viability test showed that PCG/DNA complex at the weight ratio of 20 has significantly lower cytotoxicity than that of PEI. The transfection efficiency and cytotoxicity of PCG/pDNA complexes were also confirmed in two human tumor cell lines. As shown in Supplementary Figs. S1–S4, PCG successfully mediated GFP protein expression in human liver cancer SMMC-7721 and human neuroblastoma SY5Y cells with minimal cytotoxicity. The weight ratio of 20 was selected in the following studies due to the balanced gene transfection performance and cytotoxicity.
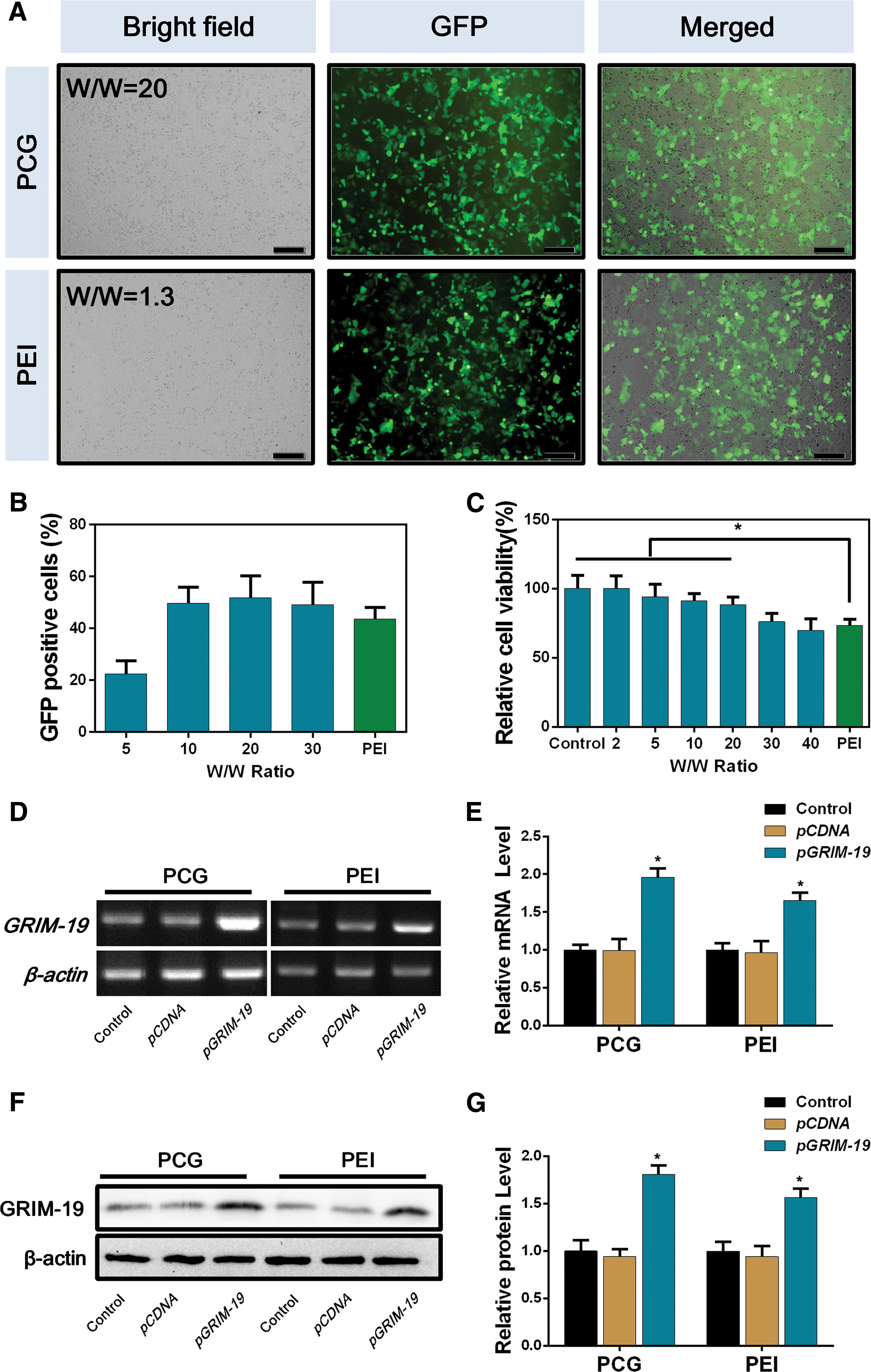
Transfection efficiency and cytotoxicity of PCG/DNA complexes in Neuro-2a cells.
After the demonstration of successful transfection using the reporter gene, we quantified the transcribed mRNA and the expressed GRIM-19 protein after the delivery of GRIM-19 encoding plasmid into Neuro-2a cells, respectively. As shown in Fig. 2D–G, the GRIM-19 mRNA and protein expression in the PCG group were twofold higher than both of the control groups, which were also significantly higher than the PEI-mediated transfection.
In vitro tumor cell inhibition by PCG/pGRIM-19 transfection
The antitumor effects of PCG/pGRIM-19 gene transfection were studied using Neuro-2a cells in vitro. First, we assessed the effect of PCG/pGRIM-19 gene delivery on cancer cell proliferation. As shown in Fig. 3A, the PCG-delivered pGRIM-19 effectively inhibited cell proliferation by 26% compared with the pCDNA control. The colony formation rate in the PCG/pGRIM-19 group also decreased significantly (Fig. 3B, C), confirming the successful inhibition of cell proliferation. Second, PCG/pGRIM-19 complexes-induced cell apoptosis was studied by flow cytometry. As shown in Fig. 3D and E, PCG/pGRIM-19 transfection induced significantly higher apoptosis rate (14%) than the controls (2.6%). Apoptotic alterations such as the loss of nuclear integrity were observed after pGRIM-19 transfection (Supplementary Fig. S5; Supplementary Data). Third, the PCG-delivered pGRIM-19 also strongly reduced cell migration (Fig. 3F–I). Finally, to confirm the mechanism of cellular function inhibition, we assessed the expression of downstream genes of GRIM-19, which were associated with proliferation, apoptosis, and migration, respectively. As expected, PCG/pGRIM-19 transfection reduced the expression levels of Cyclin D1, BCL-2, and MMPs 2 and 9 (Fig. 3J).
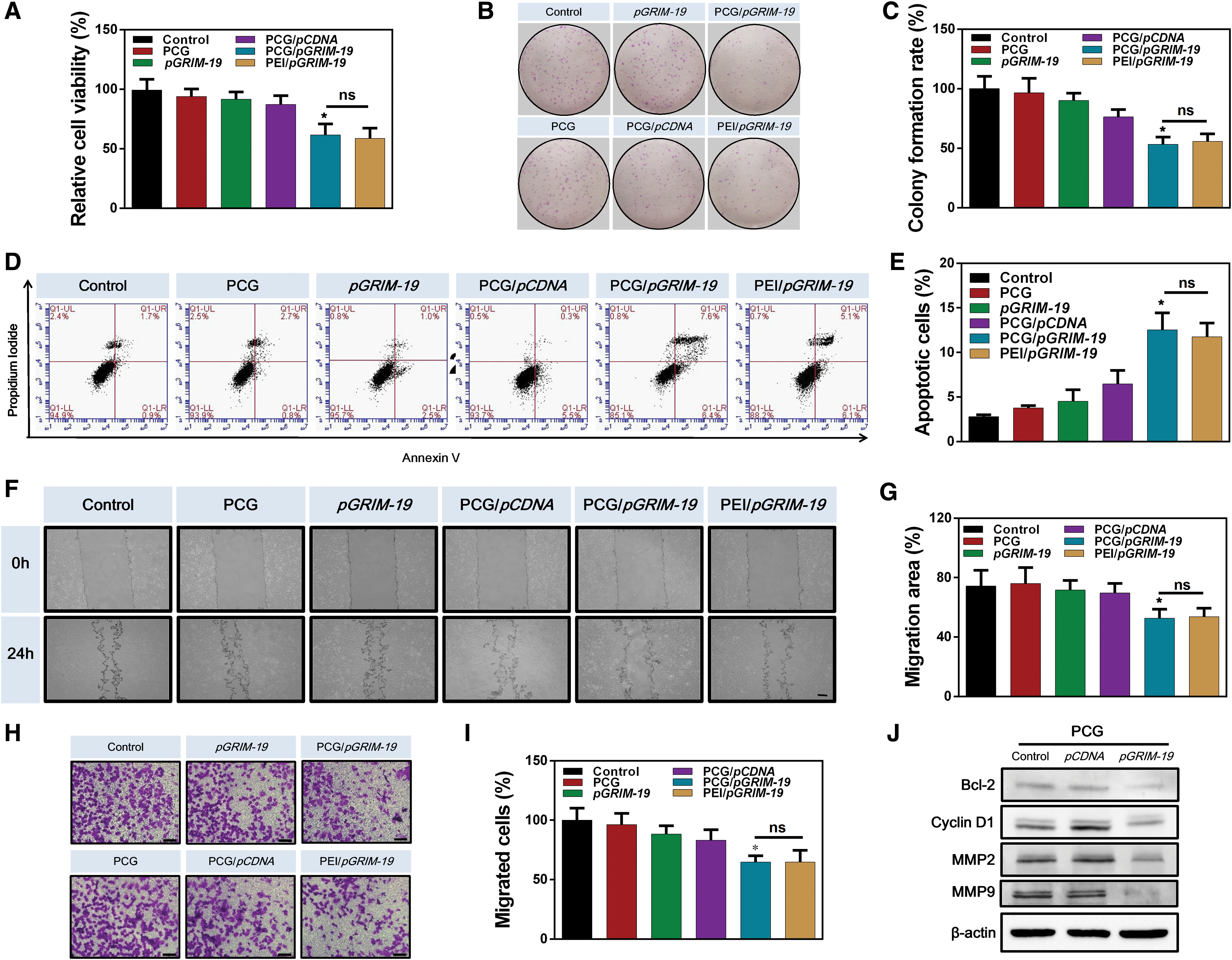
Inhibition of malignant behavior of Neuro-2a cells by PCG/pGRIM-19 complexes in vitro.
In vivo solid tumor inhibition by PCG/pGRIM-19 transfection
We next investigated the antitumor effects of PCG/pGRIM-19 complexes in a Neuro-2a neuroblastoma xenograft mouse model. The xenografted Neuro-2a tumors exhibited soft texture, bruising surface, and rapidly expanding volume, suggesting intratumor hemorrhage. Compared with the control group, tumors in the PCG/pGRIM-19 group showed a pale surface (Fig. 4A), reduced weight (Fig. 4B), and slower growth rate (Fig. 4C). The PCG/pGRIM-19 complexes inhibited tumor volume growth by 60% after four treatments (one injection every 2 days). Notably, PCG/pGRIM-19 exhibited significantly stronger tumor inhibition effect than PEI/pGRIM-19 complexes. Hematoxylin-eosin staining of tumors suggested that pGRIM-19 delivered by PCG effectively reduced intercellular erythrocyte infiltration, consistent with the appearance of the tumor (Fig. 4D). To directly examine the transfected gene expression, immunostaining was used to check the GRIM-19 protein as well as the PCNA expression. 22 The results showed that PCG/pGRIM-19 complexes upregulated GRIM-19 protein expression by eightfold compared with the control group (Fig. 4E). By contrast, PEI/pGRIM-19 only mediated an upregulation of GRIM-19 protein by less than twofold. Correspondingly, the expression of PCNA was also significantly downregulated in the PCG/pGRIM-19 group (Fig. 4F). There was no significant difference in body weight among three groups (Fig. 4G), indicating no appreciable toxicity of the delivered complexes.
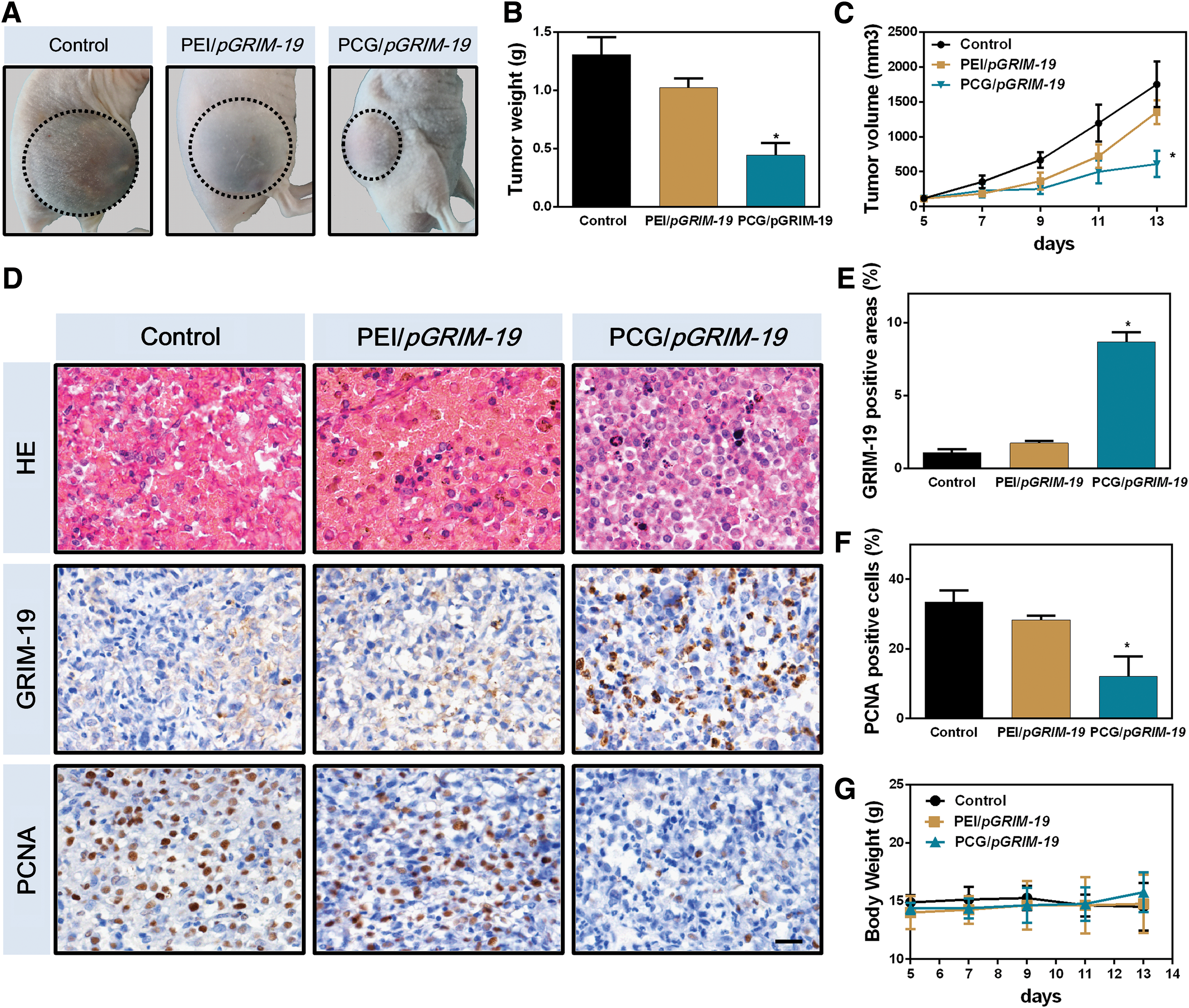
Inhibition of Neuro-2a tumors growth and hemorrhages after intratumoral injection of PCG/pGRIM-19 complexes.
Complications during treatment
To comprehensively evaluate the impact of PCG/pGRIM-19 gene therapy, we performed a survival assay and compared the results with that of cisplatin treatment. As shown in Fig. 5A, the survival rate of the PCG/pGRIM-19 group was significantly improved compared with that of the cisplatin group. Notably, the appearance of the tumors after PCG/pGRIM-19 treatment was pale in color, indicating no severe intratumor hemorrhages (Fig. 5B). By contrast, the color of the tumor became dark red after cisplatin treatment. The hematocrit of the PCG/pGRIM-19 group was higher than that of the cisplatin group (Fig. 5C), possibly due to reduced hemorrhages.

Improvement of prognosis after intratumoral injection of PCG/pGRIM-19 complexes.
Histological analysis was performed to examine the lung tissues after treatment. As shown in Fig. 5D, significant pulmonary thrombosis was observed in the cisplatin group, whereas little pulmonary embolism was seen in the gene therapy group. Blood von Willebrand factor (vWF) levels were also measured in this study, as shown in Fig. 5E. The plasma vWF levels in the cisplatin group were significantly increased (indicating endothelial damage), whereas its levels in the PCG/pGRIM-19 group were downregulated compared with the untreated control. Finally, we examined the expression of inflammatory factors in tumor tissues by immunostaining. As shown in Fig. 5F, the expression levels of STAT3, NF-κB, and MMP9 were all downregulated in the PCG/pGRIM-19 group.
Discussion
Tumor-associated complications, including inflammation, hemorrhages, and thrombosis, remain as challenges in traditional cancer treatment. 1 –3 Instead of reducing these complications, chemotherapy may worsen the situation due to induced endothelium injury. 7,8 Regulation of genes associated with these complications is a potential choice for the treatment. GRIM-19 inhibits STAT3 and NF-κB signaling pathways, which not only suppress tumor growth, but also alleviate cancer-related inflammations. 11,12 Gene therapy with GRIM-19-encoded plasmids might be a therapeutic strategy for tumor treatment to reduce these side effects in cancer patients. In this study, we selected cationic PCG as the delivery vehicle due to its high potency and limited toxicity. 20
GRIM-19 is known to regulate the proliferation- and invasion-associated genes, including CCND1, BCL-2 family, and MMPs. 12 Cyclin D1 is known to regulate G1/S phase transition, thus downregulation of cyclin D1 results in cell cycle arrest. 23 High expression of the antiapoptotic gene BCL-2 is associated with multidrug resistance in tumor cells. 24 Downregulation of BCL-2 makes tumor cells more sensitive to the inducers of apoptosis. 25 The matrix metalloproteinases MMP-2 and MMP-9 are involved in metastatic cell invasion. MMP-9 is also known to affect multiple inflammatory and protumorigenic processes. 26,27 Downregulation of MMP-9 would inhibit cell migration and inflammation. Our in vitro data showed that the downstream protein cyclin D1, BCL-2, and MMPs 2 and 9 were all downregulated after the transfection of PCG/pGRIM-19 complexes. As a result, the cancer cell proliferation and migration were inhibited, whereas apoptosis was promoted. Overall, ectopic expression of GRIM-19 exerted an anticancer effect in Neuro-2a cells, which is consistent with other studies. 28,29
We comprehensively evaluated the performance of PCG/pGRIM-19 treatment in a xenografted Neuro-2a tumor model. Neuro-2a tumor model could represent well-vascularized tumors with intratumor hemorrhages, as many hemocytes can be observed among the tumor cells. 30 Our results confirmed that PCG-mediated pGRIM-19 delivery resulted in high expression of GRIM-19 proteins and downregulated PCNA in tumor tissues, successfully suppressing tumor growth.
The overall treatment outcome of PCG/pGRIM-19 was compared with that of cisplatin. Cisplatin is a chemotherapy medication used to treat various advanced cancers. 31 It interferes with DNA replication, which kills the fastest proliferating cells. The administration of cisplatin caused injury to the endothelium, 1 indicated by increased blood vWF concentrations. 32 Cisplatin treatment also stimulated STAT3 and NF-κB expression, which are the major transcription factors that regulate inflammation levels. 3 As a result, local and systemic complications, including hemorrhage, anemia, and pulmonary embolism, were all observed after cisplatin treatment. By contrast, PCG/pGRIM-19 gene therapy downregulated all these factors, resulting in alleviated inflammation and complications. Overall, the biocompatible PCG-mediated GRIM-19 gene delivery could effectively inhibit the inflammation associated with cancer without causing endothelial damage, thereby reducing local and systemic complications, and prolonging the survival of tumor-bearing mice.
Conclusion
Our studies show that pGRIM-19 delivered by PCG effectively suppressed malignant cells by inhibiting their proliferative and invasive signals. Unlike conventional chemotherapy that causes endothelial dysfunction, PCG/pGRIM-19 gene therapy significantly reduced local complications of intratumor hemorrhages and systemic complications such as anemia and pulmonary embolism. This approach effectively prolonged the survival of Neuro-2a neuroblastoma xenografted mice. These results highlight the potential of gene therapy in reducing tumor burden and complications, providing novel strategies to enhance the clinical cancer therapy.
Footnotes
Author Disclosure
No competing financial interests exist.
Funding Information
This study was supported by the National Natural Science Foundation of China (Grant Nos. 31571443, 81773217, and 81472344), Jilin Provincial Education Department (Grant No. Jijiaokehezi [2016]455), Jilin University Bethune Plan B Projects (Grant No. 2015220), and Jilin Provincial Research Foundation for the Development of Science and Technology Projects (Grant No. 20190701065GH).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.