
Abstract
Both subretinal dosing and intravitreal (IVT) dosing of adeno-associated virus (AAV) in higher species induce mild and transient inflammatory responses that increase with dose. Foreign protein and foreign DNA are known inducers of inflammation, which is also true in the immune-privileged ocular environment. We explored which component(s) of AAV vectors, viral capsid, or viral DNA drive inflammatory responses. Recombinant AAV with three tyrosine to phenylalanine substitutions in the capsid of AAV serotype 2 (rAAV2tYF), and with a generic ubiquitous promoter (cytomegalovirus [CMV]) controlling the expression of humanized green fluorescent protein (hGFP), was processed to enrich for AAV capsids containing genome (full capsids), capsids without genome (empty capsids), and residual material. Nonhuman primate eyes were injected by IVT in both eyes. During in-life, ocular inflammation and development of neutralizing antibodies (NAb) were measured. Following termination, lymph node immunophenotyping was performed, vitreous was processed for cytokine and RNAseq analyses, and ocular sections were assessed for transgene expression (by in situ hybridization) and histopathology. IVT dosing of AAV vectors transiently raised cellular inflammation in the aqueous and induced a more sustained inflammation in the vitreous. Lowering the total capsid dose by removing empty AAV capsids reduced inflammation and improved viral transduction. IVT dosing of AAV induced systemic NAb to AAV irrespective of the vector preparation. Similarly, lymph node immunophenotyping revealed identical profiles irrespective of viral preparation used for dosing. Immune cells in the vitreous were identified based on RNAseq analysis. Three months postdose, cytokine levels were low, indicative of minimal levels of inflammation in agreement with histopathological assessment of the retina.
Introduction
Recombinant Adeno-associated Virus (rAAV) is a popular viral vector for gene therapy. AAV is a helper-dependent single-stranded DNA parvovirus that does not cause disease in humans or animals. AAV can efficiently induce gene expression in dividing and stably in nondividing cells, and the vector has a favorable safety and immune response profile. Furthermore, the AAV capsid and payload (promoter and cDNA transgene) can readily be modulated to impact cellular tropism. 1
rAAV-based gene therapeutic approaches are under continuous development for diseases in ocular, neuromuscular, liver, lung, and central nervous system. As of 2018, more than 230 clinical trials using AAV as the gene delivery vehicle were completed or in progress. * AAV-based gene therapy recently received the first FDA-awarded BLA in the United States for Leber's Congenital Amaurosis 2 (Luxturna®; Spark Therapeutics). Multiple human clinical trials are at present in progress for a variety of inherited retinal diseases (IRDs), including achromatopsia, retinitis pigmentosa, and choroideremia.
To date, routinely used routes of administration (ROA) of AAV-based gene therapy for IRDs are subretinal (SR) or intravitreal (IVT) injection. The different ROAs and associated considerations have been discussed before. 2 SR injection deposits the therapeutic vector between the photoreceptor layer and retinal pigment epithelium (RPE) cells placing the vector directly adjacent to the target cells. SR injection, however, is somewhat more invasive, requires substantial surgical skill, and carries risks of damage to retinal tissue (macular holes, SR hemorrhage, fibrosis, or retinal detachment). Furthermore, SR injection delivers the vectors only to the retinal area lifted from the RPE during surgery; expansion of the AAV vector beyond the dosing “bleb” is limited, and therefore, SR injection by definition is not panretinal.
In contrast to SR injection, IVT injection provides a potential panretinal approach since the therapeutic vector is deposited in the vitreous, enabling exposure of the vector to the entire retina and also off-target tissues within the eye (e.g., ciliary body cells and resident immune cells in the vitreous). In contrast to SR injection, the dosing procedure is minimally invasive; in fact, IVT injection is the most common ocular procedure in many clinics and is routinely performed (e.g., anti-vascular endothelial growth factor [VEGF] therapies). 3 However, to reach photoreceptor and RPE cells, the vector must pass through the inner limiting membrane (ILM) and traverse several retinal layers, which has made targeting of these layers challenging. Substantial resources are being committed to identify AAV capsid variants capable of ILM crossing for (panretinal) delivery to the photoreceptor and RPE cells in higher species. 4 –8
Both SR and IVT ocular AAV vector dosings in higher species induce mild and temporary inflammatory responses, which are stronger when dose levels increase. 9 –12 Foreign protein and foreign DNA are known inducers of inflammation, which is also true in the immune-privileged ocular environment. 13,14 We designed a series of studies to explore which component(s) of AAV vectors, viral capsid, or viral DNA drive inflammatory responses following IVT dosing of AAV vectors.
rAAV with three tyrosine to phenylalanine substitutions in the capsid of AAV serotype 2 (rAAV2tYF), and with a generic ubiquitous promoter (cytomegalovirus [CMV]) controlling the expression of humanized green fluorescent protein (hGFP), was prepared using AGTC's proprietary herpes-assisted vector expansion (HAVE) platform. 15,16 The viral preparations were further processed to enrich for AAV capsids containing genome (full capsids), capsids without genome (empty capsids), and residual material. Nonhuman primate (NHP) eyes were injected IVT in both eyes. During in-life, ocular inflammation and development of neutralizing antibodies (NAb) were measured. Following termination, lymph node immunophenotyping was performed, vitreous was processed for cytokine and RNAseq analyses, and ocular sections were assessed for transgene expression (by in situ hybridization [ISH]) and histology.
We found that IVT dosing of AAV vectors transiently raised cellular inflammation in the aqueous and induced a more sustained inflammation in the vitreous. Lowering the total capsid dose by removing empty AAV capsids reduced inflammation and improved viral transduction. As reported by others, IVT dosing induced an increase of systemic NAb to AAV irrespective of the vector preparation. Similarly, lymph node immunophenotyping revealed identical profiles irrespective of viral preparation used for dosing. Immune cells in the vitreous were identified based on RNAseq analysis. Interestingly, cytokine profiles, 3 months postdose, did not reveal patterns unique for any of the vector preparations. The generally low cytokine levels were indicative of minimal levels of inflammation which were in agreement with histopathological assessment of the retina for all vector preparations.
Materials and Methods
“Master” vector
A research-grade preparation of rAAV2tYF-CMV-hGFP was manufactured using AGTC's proprietary recombinant herpes simplex virus (rHSV) complementation in suspension-cultured baby hamster kidney cells. 17,18 Analytical ultracentrifugation (AUC) was applied to the “Master” vector to assess the fraction of viral vector capsids that contains viral genome (percentage full capsids).
“Enriched Full” and “Enriched Empty” vector
Separation of full and empty capsids was performed as previously described. 19 Briefly, an aliquot of purified Master rAAV2tYF-CMV-hGFP (4.2 × 1013 vg) made by rHSV helper viruses was supplemented with CsCl to a final density of 1.40 gcm−3, loaded into a Beckman ultracentrifuge tube and subjected to ultracentrifugation at 60,000 r.p.m. for 20 h in a 70Ti rotor. Fractions of 1.0 mL each were collected from the bottom of the tube with the aid of a Dynamax peristaltic pump (Rainin, Oakland, CA). All fractions were measured for AAV vector genome concentration, and every second fraction was analyzed for AAV2 capsid using an AAV2 capsid titration ELISA kit (American Research Products, Inc., Belmont, MA). Fractions with corresponding full or empty capsids were pooled and buffer exchanged/formulated in Alcon balanced salt solution supplemented with 0.014% Tween 20 (BSST).
Enriched Full vector was used at two dose levels: the high-dose level “Enriched Full (HD)” and low-dose level “Enriched Full.”
Residual vector
Aliquots of the purified Master rAAV2tYF-CMV-hGFP preparations were filtered over a 20 nm molecular filter. The flow-through of the filtration process was utilized to dose as “Residual” vector.
Characterization of viral vector preparations
Vector concentration (vector genomes/mL), purity (silver-stained sodium dodecyl sulfate/polyacrylamide gel electrophoresis analysis), infectivity (tissue culture 50% infectious dose [TCID50]), endotoxin, sterility, and mycoplasma were determined as described previously, 20 and passed all criteria (Supplementary Tables S1–S4).
HAVE preps typically produce batches of ∼50% full. The “Master” vector (5.22 × 1012 vg/mL; 3.97 × 1013 capsid/mL) batch we elected to use was reported as 22.85% full, which would allow for maximizing the enrichment and our ability to see differences among the groups. IVT dosing of vector per eye based on both vector genome and capsid titer is listed in Table 1.
Vector dose levels
In-life animal studies
Studies were performed at Covance Laboratories, Madison, WI, using cynomolgus monkeys (Macaca fascicularis, female, 3–5 years of age) without pre-existing ocular findings and with NAb titers in serum below 1:16. Animals were distributed based on predose NAb titers assuring similar average NAb titers for each experimental group, and five animals were assigned per group.
IVT dosing
On day 1, all eyes were dosed with 0.1 mL of viral vector (2 × 0.05 mL with 10-min pause between first and second injection to prevent critical intraocular pressure spiking). Briefly, before sedation, the animals were given an intramuscular injection of 2 mg/kg flunixin meglumine, a nonsteroidal anti-inflammatory drug. Following anesthesia, 0.5% proparacaine was instilled in each eye. The eyelids were retracted, and the periorbital region and eyes were cleaned with dilute povidone/iodine and rinsed with sterile saline. Insulin syringes (0.3 mL, 30G needle) were used to inject the eyes at the 10 and 11 o'clock position in the right eyes and at the 1 and 2 o'clock position in the left eyes. Immediately upon completion of the dosing, a topical antibiotic (Tobrex®) was instilled in each eye. Upon recovery from anesthesia, buprenorphine was administered at 0.05 mg/kg intramuscular.
In-life measurements
Animals were checked daily for health monitoring using cage-side observations and had weekly detailed observations. Body weights were determined weekly, and qualitative food consumptions were assessed daily.
Ocular examination
Ocular examinations (OEs) were performed on Day 3, Day 8, Week 2, Week 3, Week 4, Week 5, Week 6, Week 7, Week 8, Week 10, and Week 12 (termination) using slit-lamp biomicroscopy and indirect ophthalmoscopy. Inflammatory scores were based on the Standardization of Uveitis Nomenclature (SUN) criteria system 21 (Supplementary Table S5—7). The ocular examinations included scoring for aqueous cell (AC), aqueous flare (AF), vitreous cell (VC), and vitreous haze (VH). When AC scores were 3 or higher, the eyes were treated with topical steroids (2–4 times/day). Typically, the response to topical steroid application was rapid; steroid treatment was discontinued when AC inflammation scores dropped below 3, typically at the next ocular examination (∼1 week).
Termination
At termination, the eyes were enucleated. After sampling of vitreous (∼0.2 mL) and aqueous (∼0.1 mL) fluids, the ocular globes were injected with freshly prepared (day of use) 4% paraformaldehyde fixative until turgid and placed in 4% paraformaldehyde fixative for tissue preservation (48–72 h at room temperature). Eyes were processed for paraffin embedding and histological analysis (ISH and hematoxylin and eosin [H&E] histopathology).
Postlife analysis
Vitreous sampling and processing
Vitreous samples were collected avoiding nuclease contamination. After collection, samples were immediately centrifuged for ∼10 min (400 × g; 4°C). When a pellet was visible, the supernatant was collected into another tube. When a pellet was not visible, supernatant was collected leaving ∼50 μL remaining in the tube. Both vitreous samples (supernatant and pellet) were snap-frozen in liquid nitrogen and stored at −60°C to −80°C. Samples were shipped to Ocean Ridge Biosciences, LLC (Deerfield Beach, FL), for multiplex protein profiling and RNA sequencing analysis.
Cytokine analysis
Vitreous supernatant samples were assessed in a multiplex immunoassay using the Milliplex Magnetic Non-Human Primate Cytokine Assay (Millipore) designed for simultaneously measuring the following analytes: GM-CSF, TGFα, G-CSF, IFNγ, IL-2, IL-10, IL-15, sCD40L, IL-17A, IL-1ra, IL-13, IL-1β, IL-4, IL-5, Il-6, IL-8, MIP-1α, MCP-1, TNFα, MIP-1β, IL-12/23(p40), VEGF, and IL-18. The assay was performed according to the manufacturer's protocol and was read on the Luminex 100 instrument. Luminex xPONENT 3.1 software was used for assignment of each bead to a specific analyte based on its endogenous red fluorescent signal, and quantification was achieved via detection of green fluorescence from bead-associated phycoerythrin, which correlated with the abundance of the bead-bound target. The standard curve from each assay plate was used to calculate the analyte concentrations of the unknown and control samples using a 5-point parametric nonlinear regression analysis. The reported analyte concentrations were calculated by the xPONENT 3.1 software and then adjusted for dilution. Analytes detected in at least 25% of the vitreous supernatant samples were subjected to further statistical analysis. Limits of detection (LOD) for each analyte are graphically presented.
RNAseq analysis
Total RNA was isolated from the vitreous pellet (AllPrep DNA/RNA Mini Kit; QIAGEN) and quantified (UV spectrophotometry). Following RNase-free DNase I digestion (Epicentre) and repurification (RNeasy MinElute; QIAGEN), RNA was quantified (RiboGreen) and assayed for quality (Agilent 2100 Bioanalyzer). RNA was processed into cDNA using the SMART Seq v4 Ultra Low RNA kit (Clontech). The Nextera XT DNA Library prep kit (Illumina) was used for sequencing library preparation. Quality and size distribution of amplified library were analyzed on chip-based capillary electrophoresis (Agilent 2100 Bioanalyzer). The KAPA library quantification kit (KAPA Biosystems) was utilized to quantify the cDNA libraries. Libraries were pooled at equimolar concentrations for each sequencing run and diluted further for loading onto the flow cell of the Illumina NextSeq 500. The libraries were extended and bridge amplified to create sequence clusters and sequenced with 76 nt paired-end reads plus 8 nt dual-index reads (Illumina NextSeq 500). Real-time imaging and base calling were performed using Real Time Analysis software version 2.4.11. The passed filter 75 nt read pairs were aligned to the M. fascicularis 5.0 genome. The vitreous RNAseq reads were cross-referenced against the annotated M. fascicularis database that contained 944,933 unique exons and 32,809 unique genes. The reads per kilobase million (RPKM), the relative expression of a transcript to the number of cDNA fragments that originate from it, were filtered to retain a list of genes with a minimum of ∼50 mapped reads, which is the reliable quantification threshold, in 25% or more samples.
The normalized gene counts in RPKM values for the detectable genes were imported into the globally optimized non-negative decomposition algorithm DeconRNASeq. The Expression Atlas experiment E-MTAB-3287 gene expression data of common types of cells of hematopoietic lineages were used as a source for gene expression signatures of 27 immune-related cells for this study. Human orthologs of M. fascicularis were obtained from the NCBI mapping table. Analysis in this data set was limited to the top 10 cell types for each sample.
Histopathological analysis
Ocular sections (5 μm formalin fixed, paraffin embedded, and H&E stained) were taken through the fovea and reviewed by a board-certified veterinary pathologist for tissue integrity and presence of inflammation. Findings were scored in the posterior (vitreous/retina/optic nerve) and anterior (ciliary body/iris/angle) areas of the ocular sections and were assessed according to a 5-level scoring key (Table 2).
Histopathological severity scoring
ISH RNAscope analysis
Ocular sections (5 μm formalin fixed and paraffin embedded) were processed for RNAscope® according to the protocol from the manufacturer for the RNAscope 2.5 LS Duplex Reagent platform. Briefly, sections were pretreated with heat and protease (antigen retrieval) before hybridization. RNA integrity was assessed with RNAscope probes specific for PPIB/POL2RA RNA and for background with a probe specific to bacterial dapB RNA. Samples were counterstained with hematoxylin. Probes for CMV-promoter (GenBank AF396260: nt 159-802; 12 probe pairs) and hGFP (GenBank AF275953.1: nt 12-686; 14 probe pairs) were designed by Advanced Cell Diagnostics, Inc.
Specific ISH staining signal for CMV promoter and thus the presence of vector were identified as green dots, which typically colocalized to the nucleus. ISH detection of hGFP was quantified by the number of red punctate dots counted in the peri foveal retinal ganglion cell (RGC) area (600 μm on either side of the foveal pit). The hGFP counts were averaged for each treatment group.
NAb assay
NAb titers in serum were assessed using HEK293 cells. Briefly, after complement inactivation (56°C for 30 min), samples were serially diluted in media and mixed with rAAV2tYF-CMV-luc2 (multiplicity of infection: 100 vg/cell) to measure anti-rAAV2tYF NAb titers. One hundred microliters of serum–vector mixture and controls with no serum were added to 96-well plates, confluent with HEK293 cells, and incubated at 37°C 5% CO2. Luminescence output per well was measured after overnight incubation. NAb titers were reported as the highest dilution showing >50% inhibition of transduction based on luminescence compared with luminescence from the control wells (no added serum).
Immunophenotyping of lymph nodes
The cervical and axillary lymph nodes were collected and single-cell suspensions of each lymph node were prepared. Equivalent numbers of cells were stained with antibodies to enumerate phenotypes. The analysis was based on percent-gated events due to the difficulty in consistently removing connective tissue from the lymph nodes. Heterogeneous leukocyte gating strategies of CD45 fluorescent staining and side-scatter characteristics were used to delineate leukocyte populations on the flow cytometer.
Results and Discussion
AAV vector characterization
rAAV2tYF-CMV-hGFP vector was prepared using AGTC's proprietary HSV-based production platform HAVE. AGTC's HAVE production platform has provided clinical trial material for AGTC's clinical trials (NCT02416622 [retinoschisis], NCT02935517 [achromatopsia A3], NCT02599922 [achromatopsia B3], and NCT03316560 [X-linked retinitis pigmentosa]). The HAVE platform produces vectors with a relatively high percentage of full capsids compared with more standard, and difficult to scale, technologies such as triple-transfection. 15,16,22,23 For this study, a “Master” vector rAAV2tYF-CMV-hGFP with relative low-percentage full capsid was selected to widen the range of full versus empty capsids. AUC determined 23% of the AAV virions in the “Master” vector preparation-contained full-length viral genome. “Master” vector was subsequently processed to further enrich for “Enriched Full” vector, for “Enriched Empty” vector, and to remove viral particles (“Residual”) (Fig. 1). CsCl density centrifugation separates full and empty capsids efficiently producing >95% full capsids. 24 Relative approximation of the percentage of full viral particles in enriched vector and residual material was based on their respective vg titer and capsid titer ratios relative to the AUC value of the “Master” vector preparation (Supplementary Table S8). The CsCl density gradient-purified “Enriched Full” vector was enriched over 3.2-fold from the “Master,” while the “Enriched Empty” contained less than 0.5% full capsids. Filtration removed more than 99% of viral particles (“Residual”), while the percentage full remained similar to the “Master” vector (Supplementary Table S5). Enriched Full vector was used at two dose levels: “Enriched Full (HD)” (high vg dose) and “Enriched Full” (low vg dose).
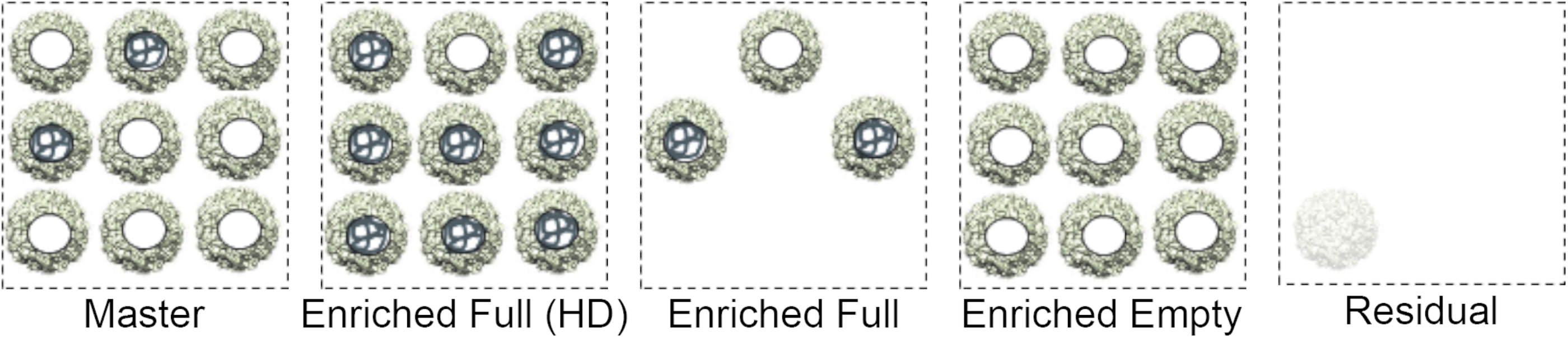
Presentation of dosing vector preparations. Spheres with filled centers represent full capsids, and the empty centers depict empty capsids. AUC was used to determine the full percentage in “Master” vector preparation. AUC, analytical ultracentrifugation.
AAV transgene expression
Levels of hGFP expression by immunohistochemistry proved insufficient to perform reliable comparison of hGFP expression between the different AAV preparations. Alternatively, we selected ISH to quantify hGFP expression in the retinal sections. AAV reporter transgene expression (hGFP) was determined using RNAscope© ISH (Fig. 2) with probe sets for the hGFP transgene designed by ACD Bio. Retinal hGFP ISH signal was limited to the RGC ring in the perifovea. Only vectors with sufficient vg content (“Master,” “Enriched Full (HD),” and “Enriched Full”) produced a robust hGFP ISH signal in the RGC ring surrounding the fovea (Supplementary Fig. S1). Dosing with “Enriched Full (HD),” equal capsid, and higher vg dose than “Master,” only minimally improved viral transduction. In contrast, dosing with “Enriched Full,” equal vg, and lower capsid dose than “Master,” enhanced viral transduction efficiency (Fig. 2). This finding was indicative of an adverse effect of empty viral capsids on transduction efficiency.
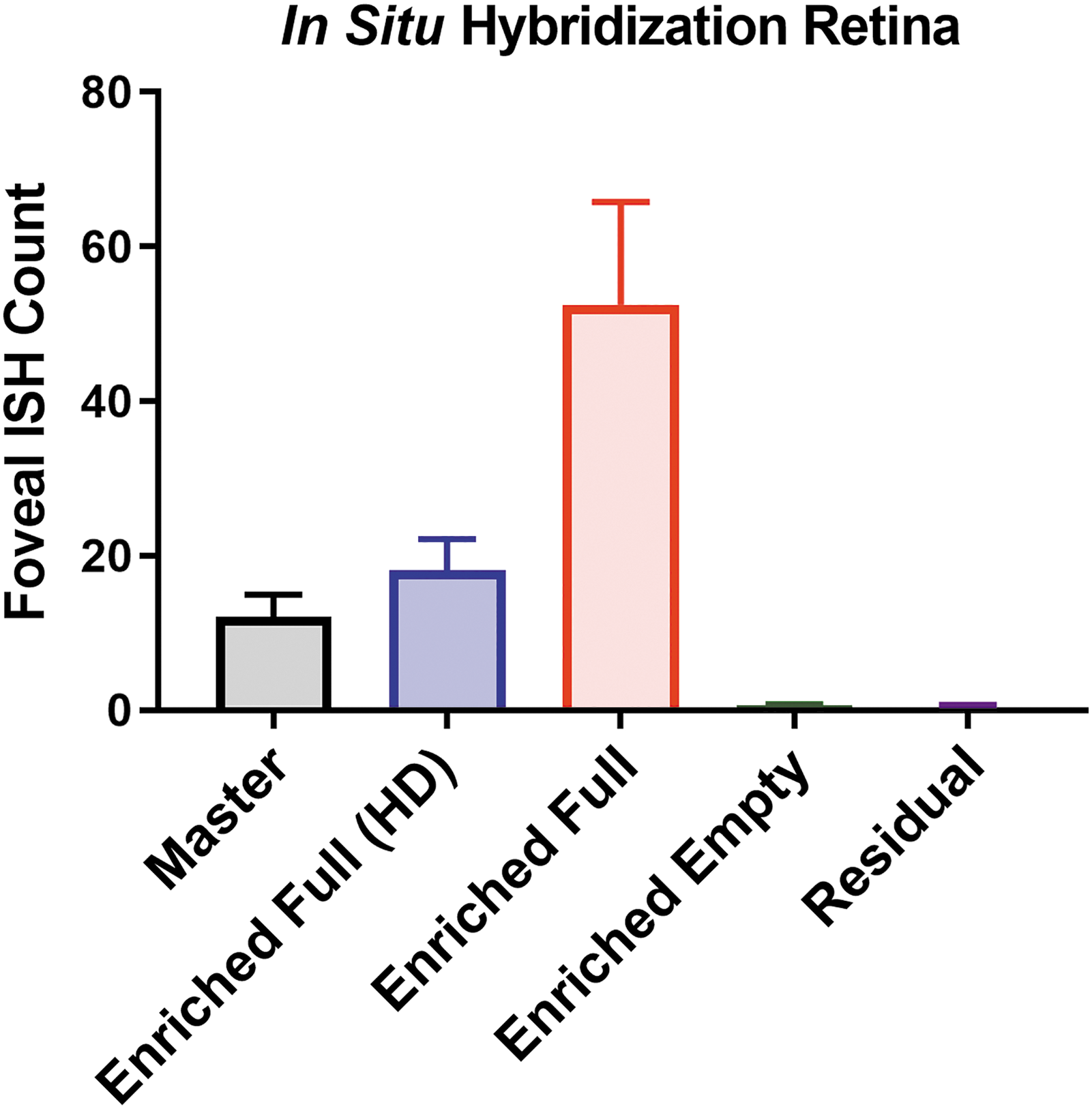
RNAscope© ISH analysis. Average counts of hGFP ISH signal in RGC in perifoveal area. Ten NHP retinas per group were sectioned through the fovea and optic nerve head. One-way ANOVA showed statistical significances with “Master.” ***p < 0.001. ANOVA, analysis of variance; hGFP, humanized green fluorescent protein; ISH, in situ hybridization; NHP, nonhuman primate; RGC, retinal ganglion cell.
Assessment of ocular inflammation
Ocular inflammation during the 3-month in-life period was assessed by scoring AC, AF, VC, and VH according to SUN guidelines. Different patterns emerged for cellular (AC and VC) and noncellular (AF and VH) inflammation. The noncellular inflammation (AF and VH) increased minimally and did not exceed an average score of ∼1 (Supplementary Fig. S2). Of note, eyes dosed with vectors with insufficient vg content (“Enriched Empty” and “Residual”) maintained baseline inflammation scores throughout the study period. Minor increases for AF and VH inflammation were observed only in eyes dosed with vectors with sufficient vg content (> 3.7 × 109 vg/eye).
IVT AAV dosing induced more robust cellular inflammatory profiles (AC and VC) (Fig. 3). The AC inflammation score in groups dosed with vector with insufficient vg content (“Enriched Empty” and “Residual”) generally remained low (≤0.5). Eyes dosed with “Enriched Full” showed AC scores slightly lower than in eyes dosed with “Master.” AC scores in eyes dosed with “Enriched Full (HD)” and “Master” showed a rapid increase. AC scores in eyes dosed with “Master,” “Enriched Full,” and “Enriched Full (HD)” vectors displayed inflammation profiles peaking at 3–4 weeks followed by a decline similar as described Reichel et al. 25 Only vectors with sufficient vg content induced observable inflammation, empty capsids alone did not induce inflammation. These findings indicate a critical role of the vector genome in cellular inflammation in the aqueous chamber.

AC and VC inflammatory scores over 12-week in-life. Inflammation score for AC and VC in eyes (n = 10) dosed with vector. Profile of average score ± SEM is plotted over the course of the in-life period. AC, aqueous cell; SEM, standard error of the mean; VC, vitreous cell.
As in the anterior chamber, eyes dosed with “Residual” vector scored low for VC inflammation throughout the study except for a brief spike at 7 weeks. However, in contrast to AC, “Enriched Empty” showed VC inflammation although at a level lower than groups where the capsids contained genome. Thus, dosing with capsid with insufficient vg (“Enriched Empty”) induced a measurable inflammation in the vitreous, which is suggestive of a role for capsids in VC inflammation. Eyes dosed with “Enriched Full” vector (vg dose equal to “Master” but with low capsid) showed substantially lower VC scores, which is also suggestive of a role for capsids in VC inflammation. Dosing with vectors with equal capsid to “Master” and increased vg content (“Enriched Full (HD)”) resulted in similar VC scores to “Master.” Thus, high capsid dose induced VC inflammation, indicative of the critical role of capsids in VC inflammation. VC scores in eyes dosed with vectors with high vg and high capsid content (“Master” and “Enriched Full (HD)” displayed a steady increase of VC inflammation plateauing at 4 to 5 weeks postdose similar as described by Reichel et al. 25 Eyes dosed with vector with “Enriched Full” or “Enriched Empty” displayed delayed inflammation that plateaued at 7–8 weeks postdose at lower levels than recorded for “Master” (Fig. 3).
In summary, IVT dosing of AAV vector induced primarily cellular inflammatory response. Induction of inflammation in the aqueous chamber required sufficient vg content (AC scores were absent when dosed with “Enriched Empty” and “Residual” vector). In contrast, VC inflammation could be induced by capsid only.
Ocular histopathology
At termination, eyes were enucleated and processed for histopathological analysis. Ocular cross-sections through the fovea were assessed for general health and inflammation (mononuclear cell infiltrates) by a board-certified veterinary pathologist. All eyes in all dosing groups showed unremarkable retinal morphology.
Histopathological inflammation scores were generally low (Table 3). In both the anterior and posterior compartments of the eye, vector with sufficient vg (“Master,” “Enriched Full (HD),” and “Enriched Full”) resulted in a higher fraction of eyes with minimal to slight mononuclear cell infiltrate. The vectors with insufficient vg (“Enriched Empty” and “Residual”) did not yield discernable inflammatory scores in the anterior or posterior chamber. The histopathological findings in the anterior chamber aligned with AC inflammation recorded during the ocular examinations. The histopathological findings in the posterior chamber aligned with VC inflammation recorded during the ocular examinations except for “Enriched Empty.” In this group, cell infiltrates were absent in the histopathological assessment of vitreous body/optic nerve/retina, while there was a measurable VC inflammation score from the in-life OE data. Histopathological assessment of inflammation in eyes dosed with AAV vector suggested that inflammation was mainly influenced by the vector genome since only vector with sufficient vg content revealed detectable inflammation in the microscopic sections.
Histopathological scoring of inflammation in nonhuman primate retinas
Lymph node immunophenotyping
We assessed the potential impact of ocular inflammation on inflammation outside of the eye by lymph node immunophenotyping. Cervical (proximal) and axillary (distal) lymph nodes were collected at termination for cell sorting to determine immune cell phenotypic profiles utilizing the following standard markers: CD3+ (total T cells), CD3+CD4+CD8+ (helper T cells), CD3+CD4−CD8+ (cytotoxic T cells), CD3−CD20+, and CD3−CD16+ cells (Fig. 4 and Supplementary Fig. S3). No differences were discernable between the AAV dosing groups for either type of lymph node or any type of cell. This indicates that the immune response did not spread widely outside of the eye or that any change was resolved before termination.

Immunophenotyping in cervical lymph nodes. Each panel compares the relative levels of immune cells across dosing groups. Analysis of axillary lymph nodes was similar to cervical lymph nodes (Supplementary Fig. S3).
Neutralizing antibodies
Serum samples were collected before the start of the study and during in-life. Serum was analyzed for NAbs against the viral vector. Figure 5 shows the NAb titers for all AAV dosing groups. Despite a wide range of ocular doses (capsid dose per eye varied from 2.9 × 1010 to 7.6 × 1011, and vg dose varied from 3.4 × 109 to 3.2 × 1011), NAb responses showed similar rates of increase for all dosing groups, and all groups reached the same maximum level. The total amount of AAV capsid or genome dosed did not impact the slope or the magnitude of the response even when only a small amount of capsid was present.

Neutralizing anti-AAV antibodies measured in serum collected on the day of dosing and 6 and 12 weeks postdosing. The NAb serum titers from each group (n = 5) were averaged and plotted ±SEM. AAV, adeno-associated virus; Nab, neutralizing antibodies.
RNAseq and cytokine analysis of vitreous samples
To obtain further insight into the cells associated with the inflammatory process in the vitreous, samples collected at termination were analyzed for cytokines and cellular RNAseq profiles. The cellular material was concentrated by low-speed centrifugation and processed for RNAseq analysis. The acellular component (supernatant) of the collected vitreous samples was analyzed for cytokines. The majority (9 of 10) of eyes dosed with “Residual” yielded insufficient total RNA, and thus, RNAseq analysis for the “Residual” viral vector group was excluded from our analysis. The limited amount of RNA obtained was consistent with the virtual absence of inflammation in eyes dosed with “Residual.” The RNAseq expression signatures resulted in the identification of a population of unique immune cell types. Analysis of variance (ANOVA) showed that two cell types (neutrophilic metamyelocytes and cytotoxic CD56-dim natural killer cells) had higher abundance and two cell types (CD38− naive B cells and macrophages) had lower abundance relative to “Enriched Empty” (Supplementary Fig. S4).
Cytokine analysis of the vitreous acellular supernatant was performed using a Millipore 23-Plex NHP Cytokine Assay. Cytokines with levels below or not significantly different from LOD in all ocular dosing groups were excluded from further analysis. Additional selection was based on one-way ANOVA comparison against “Residual” (Supplementary Fig. S5). At termination, cytokine levels for all dosing groups were close to LOD indicative of limited or absent inflammatory activity at this time. These findings contrast with VC scored in ocular examinations, but it is possible that the cells scored in the vitreous were inactive toward the end of the study and had not yet been cleared from the vitreous.
Summary and Conclusions
We investigated the inflammatory response following IVT dosing of rAAV2tYF-CMV-hGFP in NHP eyes. The transgene utilized in the studies was hGFP, which may have impacted the immunogenicity profile of the vectors by potentially increasing the inflammatory response due to the foreign nature of the transgene. However, in our experience, hGFP seems to be well tolerated. We evaluated the contribution of vector capsid and genome to ocular inflammation and vector transduction by varying the ratio of empty versus full capsids in the dosing vector. We observed different inflammatory profiles between the ocular compartments (anterior and posterior) and between vector capsid and genome (Fig. 3 and Table 3). The typical pattern of viral transduction of the retina by rAAV2tYF vector 11,26 was observed, but the efficiency of transduction was impacted. In general, the inflammatory response was mainly cellular; the noncellular aspects (AF and VH) remained at baseline levels. AAV vector injected intravitreally increased circulating AAV NAb titers irrespective of the vector titer or genome content (Fig. 5). Immune cell profiles of the lymph nodes were not impacted by the ocular inflammation and did not reveal differences between the vector preparations (Fig. 4).
Cellular inflammation in the anterior compartment increased temporarily and was triggered mainly by vector containing sufficient viral genomes, while empty capsids did not induce AC inflammation (“Enriched Empty” vector). In addition, the severity of AC inflammation showed weak if any correlation with vg dose. In contrast, cellular inflammation in the posterior compartment (VC) lasted longer. VC inflammation was lower upon enrichment to remove empty capsids, which also improved efficacy of vector transduction supporting that hGFP was well tolerated. The severity of VC inflammation correlated with dosing level: higher dose of vector capsid resulted in higher VC scores. Assessment of inflammatory markers was performed at termination (12 weeks postdose). The cytokine levels in the vitreous of eyes dosed with “Residual” and “Enriched Empty” were at baseline levels (similar to naive eyes) indicative of limited active inflammation in these eyes, however, “Enriched Empty” had elevated VC scores. Ocular examinations, however, do not distinguish between active or inactive VC. The low cytokine levels therefore suggest inactive VC contributing to the VC score (Supplementary Fig. S5). Inflammatory cytokine markers were more likely to have been elevated earlier during in-life, during the maximum inflammation, but if so they clearly dissipated with time. Limited inflammation was also demonstrated by the histopathological analysis, which showed minor signs of inflammation in retinas (Table 3).
RNAseq analysis identified a few immune cells related specifically to capsid (“Enriched Empty”) and capsid with genome (Supplementary Fig. S4). Further research will be required to begin to understand the roles of these cells during the IVT inflammatory process. Furthermore, the present study was not designed to differentiate whether the vg content-related responses were due to the mere presence of AAV genome or whether transcriptional and/or translational activities had a role in inflammation. Additional research will be necessary to address these intriguing questions; reports by Marangoni et al. 12 and Maclachlan et al., 9 however, indicated minimal impact of gene expression on inflammation. Regardless, the data presented in this study suggest that enrichment for genome containing capsids improves retinal transduction and reduces VC inflammation.
Footnotes
Acknowledgments
We acknowledge the contribution from Jacqueline Miller, Peter Sonnentag, and contributing scientists at Covance, and ACD's Pharma Assay Services in Newark, CA, for performing the RNAscope ISH assays, including optimization and QC assessment. We acknowledge the contribution from the team at Ocean Ridge Biosciences, LLC, for their performance of multiplex protein profiling and RNA sequencing assays as well as analysis of the data sets generated. In addition, we thank Daniel R. Saban for critical review of the RNAseq and cytokine analysis.
Author Disclosure
J.A.N., H.T.T., T.F., J.L., C.S., G-J.Y., S.P, C.G., D.R.K., M.S.S., and A.M.T. are or were employees and shareholders of AGTC and have a conflict of interest to the extent that this work potentially increases their financial interests. H.T.T. and C.G., coauthors, have left AGTC.
Funding Information
No external funding was received to perform these studies.
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
Supplementary Table S5
Supplementary Table S6
Supplementary Table S7
Supplementary Table S8
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.