
Abstract
High interleukin 17A (IL-17A) expression in hepatocellular carcinoma (HCC) tissue promotes HCC development. This study explores a method to inhibit HCC growth by neutralizing IL-17A in the HCC microenvironment. A novel type 5 adenoviral vector (Ad5) that carries DNA sequences encoding specific neutralizing IL-17A recombinant antibody fragments was developed in this research. After locally injecting into tumor tissues, the Ad5 transduced into tumor cells. This leads to the expression of the anti-IL-17A recombinant antibody fragments in the HCC tissue and consequently to an inhibition of HCC growth by neutralizing IL-17A. The stability of the antibody fragments was optimized by different structures design. Stable HCC cell lines that secrete IL-17A continuously were constructed, which showed stronger invasion and migration ability than control HCC cell lines. In addition, the enhanced migration and invasion ability were partially reversed by applying the adenoviral vectors. These results suggest that IL-17A might promote HCC growth by enhancing the invasion and migration ability of hepatoma cells. The antibody fragments from Ad5 neutralized IL-17A locally, in turn inhibiting the growth of HCC tumors. In conclusion, the local administration of Ad5 vectors encoding IL-17A-neutralizing antibody fragments provides a new option for HCC immunotherapy.
Introduction
According to the GLOBOCAN statistics of 2018, hepatocellular carcinoma (HCC) ranks sixth in the global cancer incidence and fourth in mortality. 1 More than 70% of patients cannot undergo curative treatment at diagnosis, and the recurrence rate is high, even after surgical resection (recurrence up to 70% within 5 years). 2 At present, the available treatment for such recurrent and incurable HCC patients is adjuvant therapy, which includes traditional chemotherapy and targeted therapy. However, the outcome is still poor. There is an urgent need to develop new treatments. 3
With the rapid development of immunotherapy in recent decades and the understanding of the unique immune microenvironment in the liver, immunotherapy has brought new hope for HCC treatment. 4,5 A number of different immunotherapeutic approaches have been applied to HCC therapy in clinical and preclinical studies, including vaccines, adoptive T cell therapies, cytokines, gene therapy, and monoclonal antibodies to the targeted immune checkpoint. 5,6 Some of these therapies have shown good outcomes in clinical studies. For example, Nivolumab, an antibody against the immune checkpoint PD-1 that prevents T cell inhibition and T cell depletion, shows a better response than Sorafenib in HCC therapy. 7,8
Interleukin 17A (IL-17A) plays an important role in the immune microenvironment of HCC. IL-17A is an inflammatory cytokine that was identified decades ago. It was originally discovered to protect the host against pathogens, especially in the mucosa and skin, through the induction of chemokines and other inflammatory factors, the recruitment of neutrophils and other mechanisms. 9,10 In recent years, IL-17A has received more attention because T helper 17 cells (Th17) that produce IL-17A have been defined (this is the third type of helper T cells after Th1 and Th2). With further research on the tumor immune microenvironment, it has been found that in some kinds of cancer, IL-17A and Th17 cells are enriched in tumors, and are negatively correlated with the disease-free survival and overall survival of patients. IL-17A contributes to HCC growth mainly by promoting tumor angiogenesis, recruiting neutrophils and inducing IL-6 that activates signal transducer and activator of transcription (Stat) 3 and other mechanisms. 11 –13
In addition to Th17, other types of cells produce IL-17A in the HCC microenvironment, such as CD8+ cells, NKT cells, and granulocytes (Fig. 1a). 14 Based on the high expression of IL-17A in liver cancer tissue and its promoting effect on HCC development, neutralizing local IL-17A may prevent the progression of HCC and represent a promising strategy for HCC treatment.
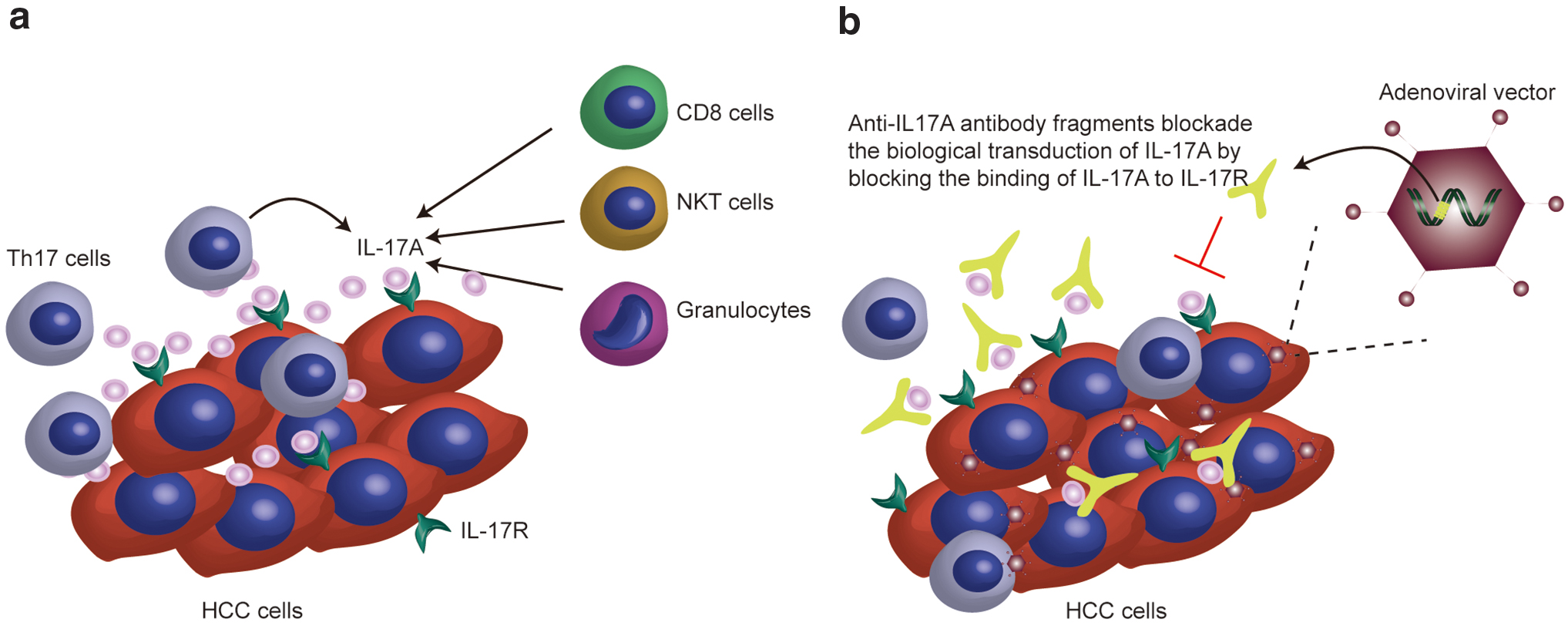
Schematic diagram showing the mechanism of the adenoviral vectors working in HCC tissue.
Anti-IL-17A antibody (AIN-457) is a human IgG1 monoclonal antibody that targets IL-17A and has been approved by the Food and Drug Administration (FDA) for the treatment of severe psoriasis and psoriatic arthritis. It selectively binds to IL-17A, preventing its binding to the IL-17 receptor. 9 However, in clinical trials, adverse events, such as upper respiratory, skin and mucous membrane infection, have been observed in patients treated systemically with the anti-IL-17A antibody, mainly due to the systemic neutralization of IL-17 that prevents the role of IL-17A in the host defense. 15,16 Therefore, when applying the anti-IL-17A antibody in HCC treatment, clinicians should consider ways to make the antibody more distributed in the lesion site and to reduce toxic or adverse effects on the normal tissue. The conventional approach is to select the appropriate targeting vector to carry the gene encoding the antibody, and this way is simpler and more efficient than applying antibodies directly. 17 On the one hand, it can reduce the binding of antibody fragments to receptors outside of the target site; on the other hand, it can protect the antibody fragments from degradation during delivery, enable the antibody fragments to be expressed locally in the tumor, and prolong the duration of the antibody fragments in the tumor. This strategy of using vectors to deliver genes is used in many antibody drugs. 18
Adenoviral vectors are one of the most commonly used gene delivery systems and are capable of infecting a broad range of cell types. 19 Among them, the five-type adenoviral vectors exhibit liver tropism and have the intrinsic ability to transduce hepatocytes, making them a preferred carrier for high-level transgene expression in hepatocytes in vivo and giving them an advantage in treating liver diseases, which have been demonstrated in many clinical trials over the past few decades. 20,21
Among the multiple potential delivery routes of HCC gene therapy, intratumoral injection and hepatic artery administration are the most advantageous, mainly due to the special blood supply mode of liver cancer (80–90% from the hepatic artery) and the mature application of image-guided techniques in the interventional treatment of liver cancer. Moreover, these two methods have been used in clinical trials. 22,23
Based on these findings, we constructed type 5 adenoviral vectors (Ad5) that carry the gene sequences encoding the IL-17A-neutralizing antibody fragment, and the vectors were administered by intratumoral injection. In our design, IL-17A-neutralizing antibody fragments should be expressed in tumors through the in situ administration of adenoviral vectors that transduce into liver cancer cells or hepatocytes, and the growth of HCC should be inhibited by neutralizing IL-17A (Fig. 1b). The results showed that the functional antibody fragments were expressed successfully in vitro and in vivo through the administration of the viral vectors; the stable HCC cell lines expressing IL-17A showed stronger invasion and migration ability, and the migration and invasion abilities were partially inhibited by the adenoviral vectors in vitro. In animal models, tumor growth is effectively inhibited by the administration of adenoviral vectors
Materials and Methods
Cell lines, animals, and adenoviral vectors
Human HCC cell lines: Hep3B, Huh7, SMMC-7721, HEK293, and 293T were obtained from the Shanghai Cell Bank of the Chinese Academy of Sciences and cultured in Dulbecco's Modified Eagle's Medium (DMEM; Gibco) plus 10% fetal calf serum (HyClone) and 1% (v/v) penicillin-streptomycin. Cells were incubated at 37°C and 5% CO2 under a fully humidified condition.
Male BALB/c nude mice, aged 4–6 weeks, were purchased from the Shanghai Experimental Animal Center of the Chinese Academy of Sciences (Shanghai, China) and housed under a specific pathogen-free condition. The study protocol was approved by the Institutional Laboratory Animal Care and Use Committee of East China Normal University, and performed according to the Guidelines of the Committee on Animal Use and Care of Chinese Academy of Sciences.
The adenoviral vectors we used were replication-deficient E1 and E3 type 5 adenoviral vectors expressing or not expressing (control adenoviral vectors) anti-IL-17A antibody fragments under the transcriptional control of the cytomegalovirus (CMV) promoter. 24
Construction and generation of viral vectors
Adenoviral vectors (AdIL-17-s, AdIL-17-sC, AdIL-17-ss, and AdIL-17-sF) carry antibodies with different structures (single-chain fragment variable [scFv], scFv-CH3, scFv-scFv, and scFv-Fc) separately. The DNA sequences of the scFv, CH3, and Fc fragments were synthesized by Sangon Biotech Corporation and amplified by quantitative polymerase chain reaction (qPCR). The amino acid sequences of these constructs are listed in Supplementary Table S1. All elements were cloned into a modified plasmid with drosophila metallothionein promoter (pMT) shuttle plasmid. In brief, the synthetic sequences were cloned into an original plasmid that had been digested using restriction enzymes by the method of homologous recombination. To equip the adenoviral vectors with the capability of secreting scFv, we cloned a signal peptide ahead of the sequence of scFv. CMV was used as the promoter in the plasmid. All constructs were cloned through ClonExpress (Vazyme, ClonExpress® II One Step Cloning Kit).
Adenoviral vector production
Adenoviral vectors were produced by cotransfecting the pMT plasmids and plasmids encoding packaging proteins (pBHG lox ΔE1,3 Cre plasmid) using Lipofectamine™ 2000 Transfection Reagent (Thermo Fisher) in HEK293 cells. After several days, the medium and cells were harvested, and then freeze-thawed several times to release the adenoviral vectors. These vectors were purified through CsCI gradient ultracentrifugation.
Expression and generation of antibodies in vitro (enzyme-linked immunosorbent assay)
The adenoviral vectors were transduced into 293T or HEK293 cells, which were cultured for 24–48 h without fetal bovine serum (FBS). The supernatant was collected and tested with enzyme-linked immunosorbent assay (ELISA) using IL-17A (Thermo Fisher, American) as the antigen and protein-L-biotin (Genscript, China) as the primary antibody.
Quantitative reverse-transcriptase polymerase chain reaction
Total RNA was extracted from cells with an RNAprep Pure Cell Kit (TIANGEN, Beijing, China) according to the manufacturer's instructions. cDNA was synthesized with a PrimeScript RT reagent Kit (TAKARA, Dalian, China). Quantitative RT-PCR was performed with a SYBR® Premix Ex Taq™ Kit (TAKARA) on a CFX96™ Real-Time System (BIO-RAD, Hercules, CA). Primer pair sequences for IL-17A and IL-17A scFv are listed in Supplementary Table S2. Relative gene expression was normalized to that of the housekeeping gene, β-actin, and is expressed as 2−ΔCT.
Invasion assay
The chambers were coated with Matrigel matrix, and then cells were seeded according to the manufacturer's instructions in the presence or absence of AdIL-17-sF. 25 After 2 days of culture, migratory cells were stained with crystal violet and then counted under a microscope. Nonmigratory cells were removed from the top of the membrane with a cotton swab.
Wound closure assay
A wound closure assay was used to assess migration capacity.
26
A total of 5 × 105 cells were seeded into 12-well flat-bottomed plates and incubated overnight. When 95% confluence was reached, the cells were incubated in the presence or absence of AdIL-17-sF, and wounds were made using a sterile 200 μL pipette tip. Then, we took pictures at 0, 24, and 72 h. The wound area was analyzed and quantified with ImageJ software (
Immunohistochemistry
Tumor tissues were fixed in 4% buffered paraformaldehyde and embedded in paraffin. After deparaffinization and antigen retrieval, tissue sections were blocked and incubated with the indicated primary antibody. The primary antibodies used included rabbit monoclonal antibodies against CD31 (Invitrogen, Carlsbad, CA) and protein-L-biotin (Genscript). The results were analyzed and quantified with ImageJ software (p < 0.05).
Tumor formation in animals and AdIL-17-sF treatment
Male BALB/c nude mice were inoculated subcutaneously with 5 × 106 Huh7-IL-17+ cells or Hep3B-IL-17+ cells (200 μL). When tumors reached 30–100 mm3 in size, mice were randomly allocated and treated with AdIL-17-sF twice, once per week. To better distribute the viral vectors in xenograft tumors, the vectors were injected into tumors at 3–4 locations. Tumor volume was calculated using the following formula: tumor volume = (W2 × L) × 0.5.
Statistical analysis
Data were analyzed using Student's t-test or two-way analysis of variance (ANOVA) to examine differences between groups. Multiple comparisons were used for comparison of given two groups after the ANOVA test. All data analyses were performed using GraphPad Prism software. The results are expressed as the mean ± standard error of the mean. The p-values are presented, and a p-value <0.05 was considered significant.
Results
Construction of adenoviral vectors with different structures of anti-IL-17A antibody fragments
The optimization of antibody fragments is mainly considered from the aspects of stability and biodistribution. It has been proven that antibody fragments with small sizes exhibit rapid biodistribution and high-level tissue penetration compared with full-length antibodies. However, antibody fragments with small sizes are always unstable. For example, the biological half-life of scFv is <1 h, and it shows short retention duration on its target. 27 Fc fragments play an important role in improving antibody stability and prolonging the half-life by FcRn-mediated recycling. The Fc fragment of IgG has been broadly used. 28 Based on the above considerations, four different IL-17A-binding antibody structures were designed using the variable regions of the heavy chain (VH) and the variable regions of the light chain (VL) from AIN-457, which has been shown to possess the ability to block IL-17A. Then, the DNA sequences of these four antibody fragments were cloned into adenoviral vectors (AdIL-17-s, AdIL-17-sC, AdIL-17-ss, and AdIL-17-sF). The deduced structures of the designed antibody fragments are shown in Fig. 2a. In brief, the VH and the VL are joined through a short, flexible serine/glycine-rich peptide linker (G4S)3, which allows the correct orientation of the VH and VL domains without interfering with their folding. 29 Based on this scFv fragment, the other three different antibody formats (scFv-scFv, scFv-CH3, and scFv-Fc) were generated. CH3 and Fc were derived from human IgG1 and fused to the C-terminus of the scFv fragments. All DNA constructs were cloned into the shuttle plasmid, and all plasmids were identified by DNA sequencing. The expression of anti-IL-17A antibody fragments was tested using qPCR (Fig. 2b).
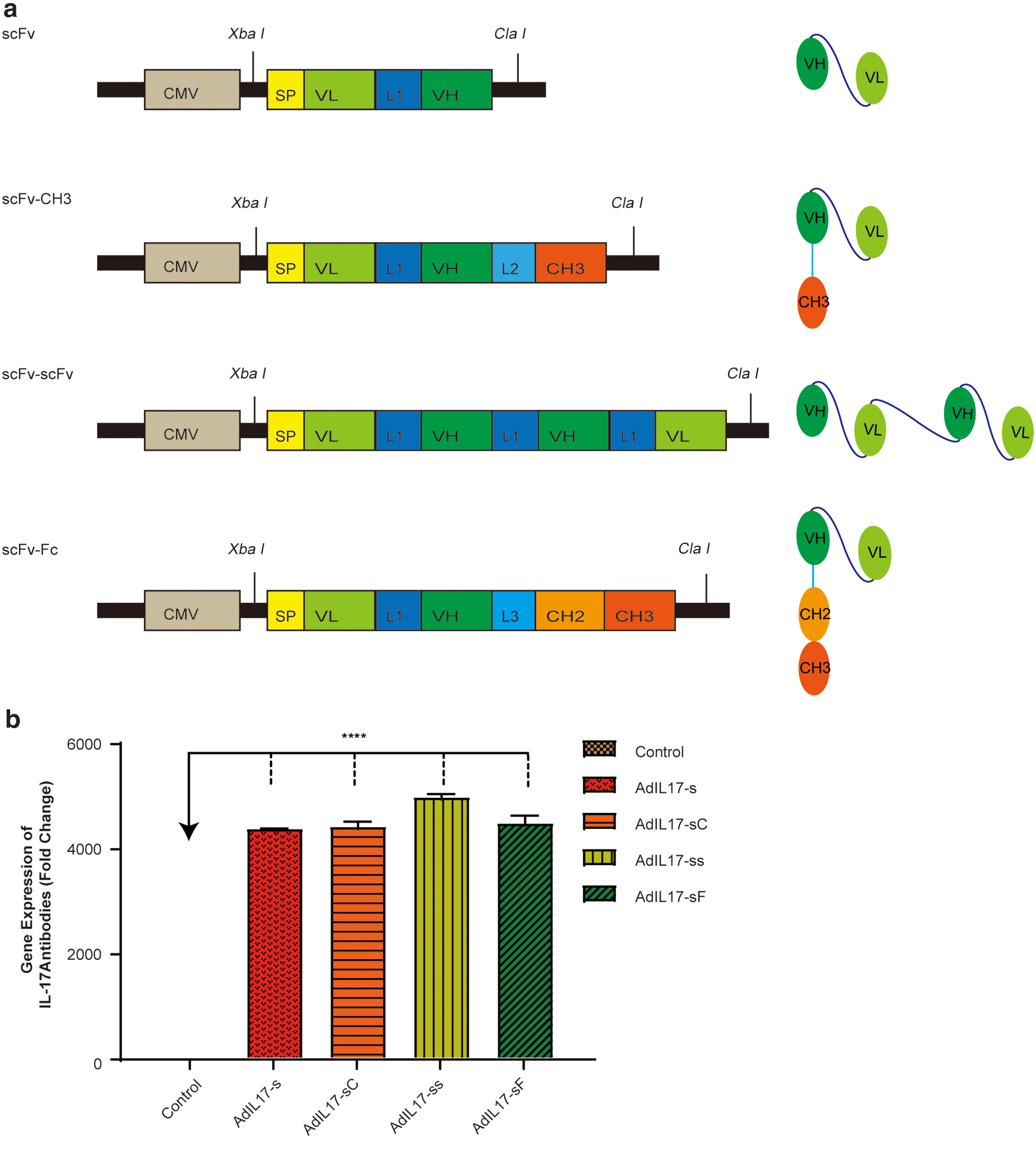
Schematic representations of the DNA templates of the adenoviral vectors (AdIL-17-s, AdIL-17-ss, AdIL-17-sC, and AdIL-17-sF) and the structures of the antibody fragments (scFv, scFv-CH3, scFv-scFv, and scFv-Fc) designed in this study and their mRNA expression levels.
The secretory IL-17A antibody fragments bind to IL-17A in vitro
To test the binding ability of the antibody fragments, 293T cells were transfected with the AdIL-17-sF plasmids and cultured for 48 h in medium without FBS. The supernatant with antibody fragments was subjected to an ELISA in four serial concentration gradients. The IL-17A protein was coated onto ELISA plates, and the supernatant of 293T cells transfected with the plasmids that did not have antibody fragment sequences was used as a control. The results showed that the cell supernatant with antibody fragments in experimental group had higher absorbance and showed a concentration-dependent response to IL-17A compared with the control group, indicating that the antibody fragments we designed could bind to IL-17A in vitro (Fig. 3a). The binding ability of the other three antibody fragments from AdIL-17-s, AdIL-17-ss, and AdIL-17-sC was also tested in the same way. The results showed that all four antibody fragments we designed could bind to IL-17A (Fig. 3b).
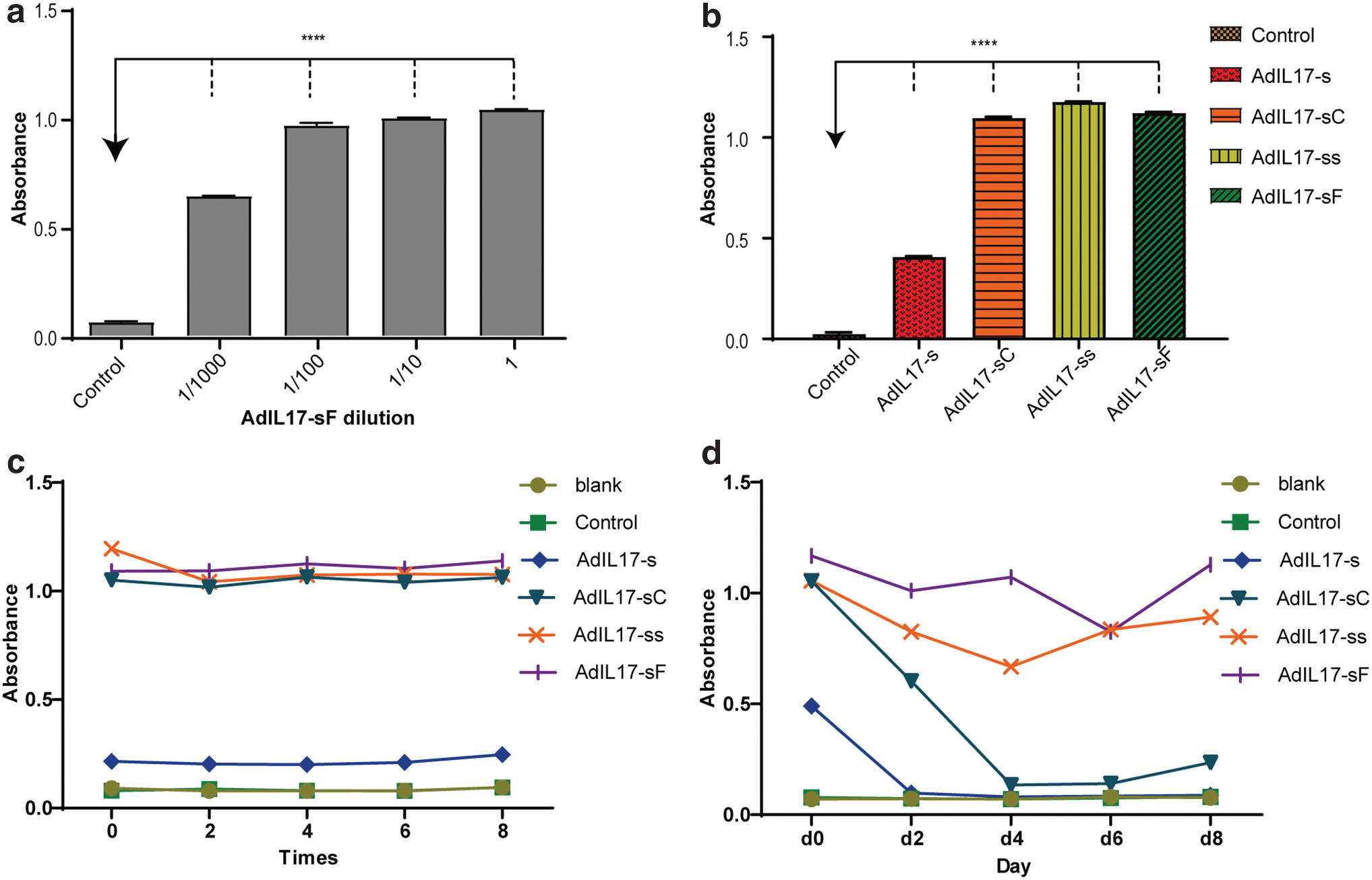
The binding ability and stability of secreted antibody fragments tested with ELISA.
Four IL-17A antibody fragments show different stabilities
To evaluate the potential instability of the antibody fragments, the supernatant with antibody fragments was freeze-thawed several times. Repeated freeze-thaw cycles can induce complex physical and chemical changes for fragments, such as a decrease in pH and oxidative stress. These stress factors may lead to conformational changes, aggregation, and denaturation, which are related to the loss of binding activity of antibody fragments. 30 –32
As shown in Fig. 3c, antibody fragments with the constant region or large molecular weights displayed higher stability at the beginning of the cycle, and over eight times, their binding activity did not decrease significantly. To further evaluate the stability of the antibody fragments in vivo, the supernatant with antibody fragments was cultured in FBS at 37°C for 8 days. From the results shown in Fig. 3d, we can see that the binding activity of fragments with scFv and scFv-CH3 decreased rapidly to the same level as the control over 4 days. However, AdIL-17-ss and AdIL-17-sF still showed high binding activity with IL-17A after 8 days. This result indicated that the antibody fragments of scFv-scFv and scFv-Fc had better stability than scFv and scFv-CH3. Therefore, AdIL-17-sF was chosen for the follow-up experiments.
Establishment of IL-17A stable HCC cell lines that constitutively express IL-17A
To determine the efficacy of the adenoviral vectors, stable HCC cell lines that secrete IL-17A continuously were needed. First, the expression level of IL-17A mRNA in seven HCC cell lines (97H, Hep3B, HepG2, Huh-7, 97L, SMMC-7721, and SNU-739) was determined by quantitative reverse transcriptase polymerase chain reaction (qRT-PCR). The results showed that all HCC cell lines expressed low levels of IL-17A (Fig. 4a). Then, we selected two cell lines with low IL-17A expression, Hep3B and Huh-7, to construct stable cell lines. In addition, the expression level of AdIL-17-sF mRNA was tested (Supplementary Fig. S1c). In brief, Hep3B and Huh-7 cell lines were transduced with the IL-17A gene by lentiviral vectors that also contain the puromycin antibiotic resistance gene. Then, the transduced cell lines were subjected to puromycin selection for 2 weeks. Expression of exogenous IL-17A protein in the established cell lines was detected by qRT-PCR and Western blot. As shown in the results, the mRNA level of IL-17A in Hep3B-IL-17+ and Huh7-IL-17+ cells was significantly higher than that in the control group (Fig. 4b); and only Hep3B-IL-17+ and Huh7-IL-17+ cells expressed IL-17A on protein level (Fig. 4c). It indicated that the stable strain was successfully constructed. The stable cell lines Hep3B-IL-17+ and Huh7-IL-17+ successfully produced IL-17A.

IL-17A expression in stable HCC cell lines.
AdIL-17-sF partially reversed the stronger invasion and migration ability of stable cell lines
First, the influence of IL-17A on HCC cell invasion was investigated. Transwell Matrigel invasion assays were performed on the Hep3B-IL-17+ and Huh7-IL-17+ stable cell lines. To rule out the effect on cell invasion ability by the viral vector itself, the stable cell line was transduced with lentiviral vectors without IL-17A (control group). As shown in Fig. 5a and c, there was no significant difference between the blank and control groups; however, more IL-17+ cells invaded through the Matrigel matrix and the membrane pores (stained with crystal violet). The invasive cells were quantified with ImageJ (p < 0.05). Therefore, the stable cell lines showed stronger invasion capability than control HCC cells, which indicates that IL-17A may promote the invasion ability of hepatoma cells in vitro.
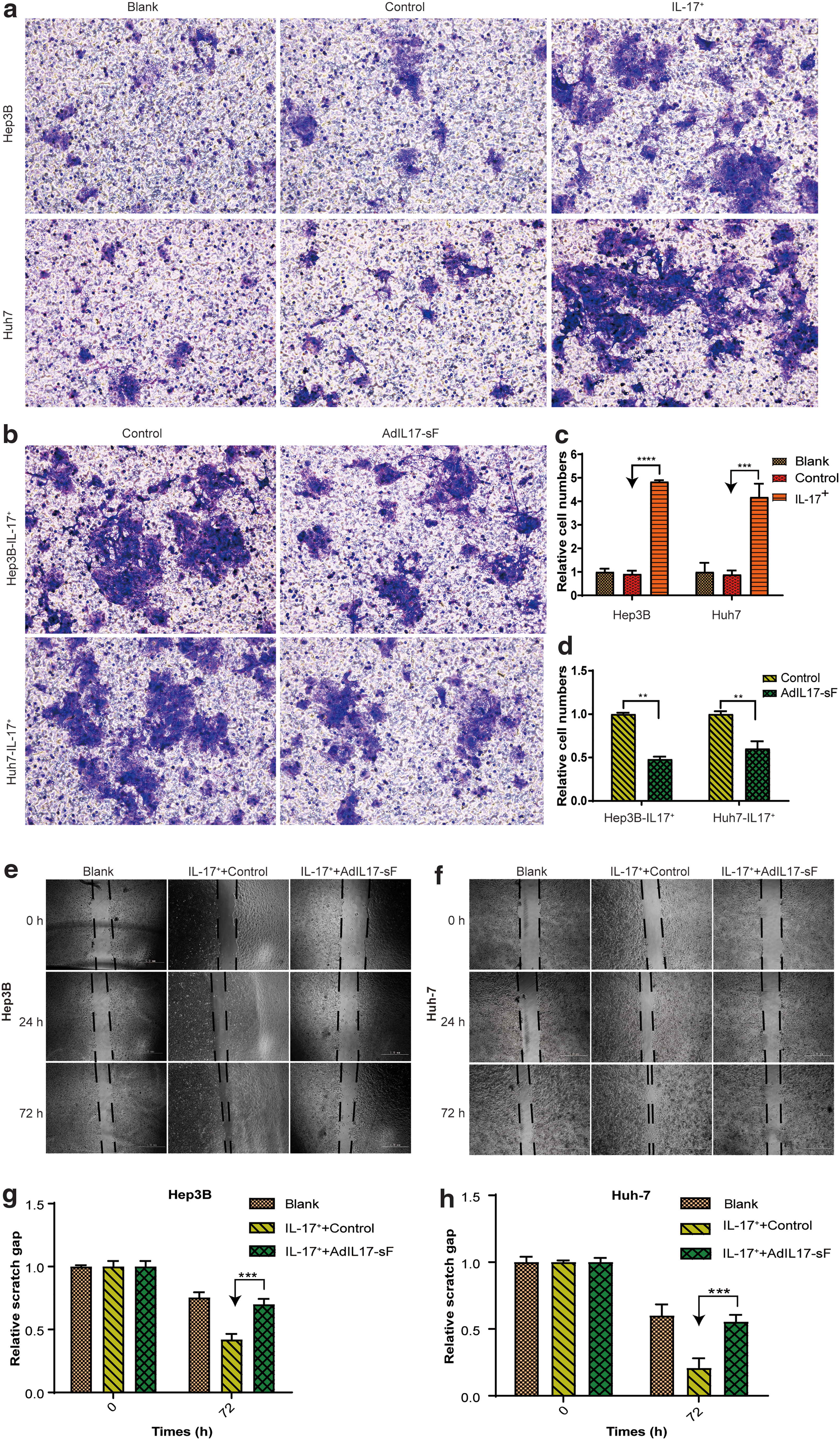
Invasion and migration capacity of stable HCC cell lines and the influence of AdIL-17-sF.
To investigate if AdIL-17-sF could successfully reverse this stronger invasive capacity induced by IL-17A, AdIL-17-sF was administered to the stable cell lines. The number of cells passing through the Matrigel matrix and membrane pores was reduced (Fig. 5b, d), indicating that the enhanced cell invasive ability of IL-17A expression was partially inhibited. This finding also indicates that after the administration of AdIL-17-sF, the expressed anti-IL-17A scFv-Fc antibody fragment neutralized IL-17A, thereby partially reversing the invasion induced by IL-17A.
The same trend in migration ability was observed in wound healing assays. The gaps in the IL-17A-positive stable strains (Hep3B-IL-17+ and Huh7-IL-17+) almost healed after 72 h (Fig. 5e, g), suggesting that these stable cell lines exhibit stronger migration ability than the control HCC cell lines (Hep3B-IL-17− and Huh7-IL-17−). The enhanced migration ability was also partially reversed after AdIL-17-sF administration (Fig. 5f, h). The gaps were quantified with ImageJ (p < 0.05) (Fig. 5g, h).
Taken together, our results suggest that IL-17A enhances the invasion and migration of hepatoma cell lines. The constructed adenoviral vector AdIL-17-sF was able to partially reverse this enhanced invasion and migration capacity, which indicates that after the administration of AdIL-17-sF, the expressed anti-IL-17A scFv-Fc antibody fragment can neutralize IL-17A, thereby reversing the enhanced invasion and migration induced by IL-17A.
We also investigated whether IL-17A or AdIL-17-sF could enhance the proliferation of HCC cells directly; the proliferation of the control cell lines and the established stable cell lines was compared. The results showed no significant difference in proliferation between the two groups during the time examined (Supplementary Fig. S1a). The effects of IL-17A or AdIL-17-sF on apoptosis have also been tested. The results showed that there was no significant difference between each group (Supplementary Fig. S1b). These results indicate that IL-17A does not promote liver cancer progression by directly promoting hepatocellular proliferation.
In vivo antitumor effect of AdIL-17-sF on xenograft models of human stable HCC cell lines
The expression of anti-IL-17A scFv-Fc antibody fragments in the tumor tissue was confirmed by staining for anti-scFv-Fc using a monoclonal protein-L-biotin antibody. From the results, local expression of anti-IL-17A scFv-Fc antibody fragments was visualized in the tumors administered with AdIL-17-sF (Fig. 6a, d, arrows). This finding suggests that the adenoviral vectors were transduced into tumor cells, and that IL-17A scFv-Fc antibody fragments were expressed in the tumor. Angiogenesis and apoptosis were also detected in situ by immunohistochemistry. It has been reported that IL-17A promotes angiogenesis and supports resistance to antiangiogenic therapy, and that IL-17A-neutralizing antibodies inhibit the growth of colon tumors. 32,33 We hypothesized that if IL-17A promotes HCC tumor angiogenesis in vivo, AdIL-17-sF could inhibit angiogenesis in the Huh7-IL-17+ stable cell line through IL-17A neutralization. The results showed that the microvessel density in the group treated with AdIL-17-sF was lower than that in the control group (Fig. 6b [arrows], e), which demonstrated that AdIL-17-sF could inhibit tumor growth by an antiangiogenic effect in vivo. There were more TUNEL-positive tumor cells in the AdIL-17-sF-treated group than in the control group, indicating that the cells in the AdIL-17-sF group underwent more apoptosis (Fig. 6c, f). This finding also indicates that the local administration of AdIL-17-sF to the tumor may induce HCC cell apoptosis.

AdIL-17-sF shows potential antitumor effects on xenograft models, and histological changes in each group were tested.

To investigate antitumor efficacy, animal models of subcutaneous tumors were established in BALB/c nude mice using the Huh7-IL-17+ and Hep3B-IL-17+ stable cell lines, and were treated with AdIL-17-sF. To exclude the possibility of tumor elimination caused by the viral vector itself, the mice were injected with control viral vectors without IL-17A antibody fragments. Mice of Huh7-IL-17+ xenograft models were sacrificed 2 weeks after first adenoviral administration; the tumor volumes of group AdIL-17-sF were smaller than those of the control groups (p < 0.05) (Fig. 6g). To explore the duration of the effect, with the same adenoviral administration on xenograft models of Huh7-IL-17+, the mice of Hep3B-IL-17+ xenograft models were sacrificed until day 26 after first adenoviral administration, and there was still inhibitory effect on the tumor growth in AdIL-17-sF group (Fig. 6i). This result indicated that tumor growth was suppressed after AdIL-17-sF treatment compared with no treatment. The body weight of mice did not change significantly, indicating that the AdIL-17-sF treatment had no obvious side effects on the mice (Fig. 6h, j).
Discussion
Here, we report a type 5 adenoviral vector carrying gene sequences encoding IL-17A-neutralizing antibody fragments in which the scFv fragments specifically bind to IL-17A. Two stable HCC cell lines Hep3B-IL-17+ and Huh7-IL-17+ were established and were used in the study. There was found to be no significant difference in the proliferation ability between the stable cell lines and the corresponding control cell line in vitro, which indicates that IL-17A does not promote the proliferation of HCC cell lines directly. However, it was found that stable cell lines possessed stronger invasion and migration ability. Therefore, we propose that IL-17A may promote HCC growth by enhancing the invasion and migration ability of hepatoma cells. This speculation was approved in this study. This IL-17A's promotion on tumor invasion has also been observed in breast cancer and colorectal cancer. 33,34 Thereafter, we tested the antitumor efficacy of the adenoviral vectors on two xenograft models of Hep3B-IL-17+ and Huh7-IL-17+,and found that tumor growth was inhibited.
In the selection of viral vectors, we chose the type 5 adenoviral vector, which is often used in experimental studies based on its clinical application and specific targeting ability. Adenoviral vectors have certain advantages, such as higher transduction efficiency, a higher level of transgene expression, and being well studied. To date, adenoviral vectors have been the most frequently used viral vectors in clinical (20.5%) and preclinical trials. 19 Moreover, the type 5 adenoviral vector exhibits intrinsic liver tropism, and Ad5 has a preferred accumulation in the liver and infects liver cells through a coagulation factor-dependent pathway and a CAR (coxsackie virus-adenovirus receptor)-dependent pathway. 20 Therefore, even if administered intravenously, the Ad5 vector is more preferentially transduced into liver tissue for targeted gene therapy. 35,36 Although we used intratumoral injection, this approach aims to achieve a high accumulation of Ad5 vectors in tumor tissue.
In the design of antibody fragments, the structure was carefully considered and optimized. Antibody drugs do not always require the Fc segment, and to some extent, this structure may increase the unpredictable immune response. However, binding between Fc and the Fc receptor enables the antibody with the Fc fragment to exhibit a longer half-life. In addition, the Fc fragment of IgG1 shows higher stability and affinity to the receptors than other antibody isotypes. Most of the FDA-approved therapeutic antibodies are in IgG1 format. 37 In addition to the Fc fragment, the size of the antibody is an important factor affecting the stability of the antibody. Under certain conditions, reducing the molecular size of the antibody may be beneficial for tissue distribution but may reduce antibody stability. Considering the size, specificity, isotype, and other factors of the antibody, we designed four antibody structures in the experiment, namely scFv, scFv-scFv, scFv-CH3, and scFv-Fc. We examined changes in the binding ability of the antibody fragments to IL-17A under different conditions to evaluate the stability of the antibody fragments. We found that the structure of scFv-Fc showed superior stability.
Although tumor growth was inhibited after the local injection of AdIL-17-sF into the xenograft model, tumor elimination was not achieved. To some extent, this finding was consistent with our expectations. IL-17A cannot directly inhibit tumor cell proliferation, which was confirmed in our in vitro cell proliferation assay. Therefore, the combination of AdIL-17-sF and chemotherapeutic drugs may be a promising way to increase the tumor inhibitory effect, which is one of our future research directions.
The immune microenvironment in the liver is special. A large number of antigenic substances enter the liver through the mesenteric vein and the portal vein. To avoid an excessive immune response, the liver acquires specialized mechanisms of immune tolerance. 38 In HCC, these mechanisms can be used by tumor cells to evade detection and clearance of the immune system. Compared with other tumors, ∼90% of HCC develops from chronic inflammation. HCC generally undergoes the classic process of the inflammation–fibrosis–cancer axis. Therefore, the immune microenvironment is crucial in the development and progression of liver cancer. The regulation of IL-17A as an inflammatory factor in the liver cancer microenvironment needs further study. Considering the complexity of the HCC microenvironment and the possible synergistic effect between the various mechanisms involved, we believe that combination therapy strategy holds a new hope. At present, there are many combination therapies in clinical trials, including the combination of traditional methods (such as TACE or chemotherapy) with immunotherapy and the combination of targeted therapies with immunotherapy, and some of them have obtained satisfactory results. 5,39 Thus, whether IL-17A and other elements in the HCC microenvironment have synergistic effects on promoting HCC development and whether the combination of gene therapies for IL-17A neutralization with other treatments can produce unexpected results remain to be further investigated.
In summary, this study demonstrated the feasibility of using Ad5 adenoviral vectors carrying a gene encoding antibody fragments to treat HCC, which confers a new approach for the treatment of solid tumors by neutralizing cytokines from the HCC microenvironment. A combination of antibody gene delivery approach with other therapeutic approaches may shed new light on HCC patients.
Footnotes
Acknowledgments
The authors thank Dr. Xiaojiang Wu and Wei Qi for their technical support.
Author Disclosure
No competing financial interests exist
Funding Information
This work was supported by the National Natural Science Foundation of China under Grant 81872812; “985” grants of East China Normal University (ECNU).
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.