
Abstract
Refractoriness to conventional chemotherapy is a major challenge in the treatment of advanced ovarian cancer (OC). There is increasing evidence that mitochondrial priming correlates with cisplatin response in various cancers. Notably, Bim and Bid, two of the proapoptotic BH3-only proteins, are recognized as the most effective inducers of mitochondrial priming in OC. In this study, we constructed two tumor-specific oncolytic adenoviruses (Ads) coding for Bim (Ad-Bim) or truncated Bid (Ad-tBid), respectively, and performed gain-of-function assays in nine OC cell lines. Ad-tBid exhibited significant antitumor efficacy than the controls. On addition of Ad-tBid pretreatment, mito-primed cells displayed more sensitivity to cisplatin both in vitro and ex vivo. We also found that Ad-tBid induced mitochondrial apoptosis in a Bak-dependent manner. Furthermore, a combined cisplatin plus Ad-tBid therapy markedly inhibited tumor growth in a subcutaneous xenotransplanted tumor model. In mice bearing peritoneal disseminated OC, intraperitoneal administration of Ad-tBid potentiated the antitumor effect of cisplatin. Our findings suggest that Ad-tBid enhances cisplatin response in OC cells, establishing the potential treatment of advanced OC via a combination of cisplatin and Ad-tBid.
Introduction
Ovarian cancer (OC) is the most lethal gynecological cancer that annually accounts for 150,000 deaths among women worldwide. 1 About 80% of patients with advanced disease will relapse within 5 years after diagnosis, and most will eventually become resistant to chemotherapy. 2 Once OC patients progressed to resistance to cisplatin, one of the most commonly used chemotherapeutic agents against OC, they have an average progression survival duration of 3–4 months and a median overall survival duration of 9–12 months. 3 More frustratingly, <15% of these patients will respond well to further treatment. 3
Considerable efforts have been devoted to exploring novel strategies to treat patients with advanced OC. Numerous results have suggested that mitochondrial priming is the major determinant of chemotherapy effectiveness and should be exploited for cancer therapy. 4 Mitochondrial priming is defined as a measure of the proximity of mitochondria to the threshold of apoptosis. 5 Mitochondria loaded with proapoptotic proteins are more predisposed toward apoptosis, and this cellular milieu is referred to as “primed to death.” 6 A previous study demonstrated that chemosensitive tumors were highly primed, while chemoresistant tumors were poorly primed. 7 However, it is unclear what determines the level of priming within tumor cells.
A compelling hypothesis is that the level of priming is determined, at least in part, by the degree of expression and posttranslational modification of the BCL-2 family proteins. 8 Simply, BCL-2 family members are divided into three categories based on their sequence homology and intracellular function. 9,10 One category includes the prosurvival proteins BCL-2, BCL-w, MCL-1, BFL-1, and BCL-XL, which contain all four BCL-2 homology domains (BH1-4). The second category, the BH3-only proteins, includes Bim, Bid, Puma, Bad, Bik, Noxa, and Bmf, which contain only the BH3 domain. 11,12 The final category consists of the effectors Bak and Bax, whose activation requires the binding of the proapoptotic BH3-only “activator” protein Bim or Bid. On activation, BAK and BAX oligomerize and directly cause mitochondrial outer membrane permeabilization (MOMP), a critical event during apoptosis. 13
The development of oncolytic viruses as anticancer therapeutics has recently accelerated. 14,15 Among various vectors, adenoviruses (Ads) have efficacy, safety, and manufacturing characteristics that make them attractive as oncolytic virus candidates and suitable gene delivery vectors. 16,17 Replication-selective oncolytic Ads, which specifically infect tumor cells while sparing normal cells, represent a novel cancer treatment platform. 18 We have previously reported a type of oncolytic adenoviral vector system by substituting the Ad death protein (ADP) gene in the E3 region with a fragment of an extraneous transgene in an E1A CR2 region-deleted adenoviral mutant. 19 –21 Based on this system, we have constructed several kinds of oncolytic Ads, which showed potent antitumor efficacy by causing selective cytolysis with no obvious adverse effects. 22,23
Considering that Bim and Bid are recognized as the most effective inducer of mitochondrial priming in OC, 5 we armed two oncolytic Ads with Bim or tBid to generate Ad-Bim/Ad-tBid. Our results demonstrated that Ad-tBid enhanced cisplatin-induced apoptosis in OC cells even at a low multiplicity of infection (MOI). This effect was further confirmed in primary OC cells and xenograft mouse models, suggesting that Ad-tBid-cisplatin combinational therapy may serve as a novel therapeutic strategy against advanced OC.
Materials and Methods
Clinical tissue samples
Fifteen fresh female ascites samples were obtained from serous OC patients staged IIIC or IV (International Federation of Gynecology and Obstetrics staging). All patient tissues were collected from the Department of Gynecological Oncology (Tongji Hospital, Huazhong University of Science and Technology, Wuhan, China) between September 2015 and June 2017 with the informed consent of patients and the authorization of the Ethics Committee of Tongji Hospital.
Isolation of cancer cells from OC ascites fluid
Ascites were dispensed into 50 mL centrifuge tubes. Then, 1 mL of 0.25% trypsin was added into the tube and incubated at 37°C shaking table for 30–40 min to disaggregate the sphere cells. After incubation, the tubes were centrifuged at 1,000g for 5 min and the cell sediment was resuspended in 4 mL McCoy's 5A medium. Density gradient centrifuge tubes were paraded by filling in 4 mL of 90% Percoll Plus (GE Healthcare Life Science, Pittsburgh), and then filled up softly with 4 mL of 45% Percoll Plus. Then 2–4 mL of the cell suspension was applied to the top gently and the mixture was centrifuged at 1,500g for 30 min at 4°C. After concentration, the culture medium is layered on the top and epithelial OC cells remain in the layer above the 45% Percoll Plus. 24 Then the cancer cells were carefully removed with a pipette and washed twice with McCoy's 5A medium containing 15% fetal bovine serum (FBS). At last, the cancer cells were transferred to the culture plates with fresh medium.
The purity of primary cells was verified with epithelial cell adhesion molecule (EPCAM) (the epithelium-specific marker) (ab8666; abcam)-based flow cytometry analysis, and only those with epithelial percentage >90% were used.
Viruses
Four different viruses were used in this study. First, Adv5/dE1A with deletion of 920–946 bp in E1A CR2 was constructed and stored in our laboratory as described previously. 19,22 Second, the replication-deficient Ad-dE1A is an E1A CR2 and E3 ADP deletion adenoviral mutant containing the enhanced green fluorescent protein (EGFP) gene in the deleted ADP region. Finally, on the basis of Ad-dE1A, Ad-Bim and Ad-tBid were constructed by replacing EGFP with Bim/tBid. All the viruses were derived from the AdEasy system (Agilent), 25 produced in HEK293 cells, and purified by a two-step CsCl gradient ultracentrifugation as described. 26,27
Cell lines
All human OC cell lines were purchased from American Type Culture Collection (Manassas, VA). SKOV3 was cultured in McCoy's 5A medium; TOV112D, OV90 and TOV21G were cultured in 1:1 medium 199/MCDB105; A2780, OVCAR3, OVCAR4, OVCAR8, and HOC7 were cultured in Roswell Park Memorial Institute (RPMI) 1640. All the mediums were supplemented with 10% FBS (Gibco, Carlsbad, CA), and all cells were cultured at 37°C in a humidified atmosphere containing 5% CO2.
Reverse transcription quantitative polymerase chain reaction
Total RNA was routinely extracted using TRIzol reagent (Invitrogen, Inc., Carlsbad, CA), and RNA quality was examined by NanoDrop (Thermo, Waltham, MA) analysis. Retrotranscription polymerase chain reaction was performed using the Maxima First Strand cDNA Synthesis Kit (Fisher Scientific, Pittsburgh, PA). The relative quantity of mRNA was determined by quantitative polymerase chain reaction (qPCR) using a CFX96 Touch Real-Time PCR Detection system (Bio-Rad Laboratories, Inc., Hercules, CA) with SYBR Green Supermix (Bio-Rad Laboratories, Inc.). The forward sequence and reverse sequence of Bim were 5′-TAAGTTCTGAGTGTGACCGAGA-3′ and 5′-GCTCTGTCTGTAGGGAGGTAGG-3′, respectively. The forward sequence and reverse sequence of Bid were 5′-ATGGACCGTAGCATCCCTCC-3′ and 5′-GTAGGTGCGTAGGTTCTGGT-3′, respectively. Glyceraldehyde phosphate dehydrogenase (GAPDH) was used as the internal control (forward sequence: 5′-GGAGCGAGATCCCTCCAAAAT-3′; reverse sequence: 5′-GGCTGTTGTCATACTTCTCATGG-3′).
Antibodies and Western blot assay
To prepare whole-cell lysates, cells were collected and washed with precooled phosphate-buffered saline (PBS) three times and were lysed by adding radio immunoprecipitation assay (RIPA) buffer (Beyotime Biotechnology Co., Shanghai, China). After thorough mixing and incubation on ice for 30 min, lysate was centrifuged at 1,2000 rpm at 4°C for 15 min, and supernatants were collected and packed at −20°C for preservation. After determination of the protein content, 30 μg cellular lysates were separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and electrotransferred onto polyvinylidene difluoride membranes. After being blocked with 5% bovine serum albumin (BSA) in tris buffered saline tween (TBST), the membranes were incubated with primary antibodies at 4°C for at least 12 h, followed by 1:2000 horseradish peroxidase (HRP)-conjugated secondary antibody for 1 h at 37°C. Relative expression level of proteins was analyzed using Image Lab software (Bio-Rad Laboratories, Inc.). The following primary antibodies were used: Bid (no. 8762), Bim (no. 2933), Bak (no. 6947), Bax (no. 5023), Cleaved-parp (no. 5625), Phospho-histone H2AX (no. 9718) from Cell Signaling Technology; GAPDH (sc-32233) and α-Tublin (sc-53646) from Santa Cruz.
3-(4,5-Dimethylthiazol-2-yl) -2,5-diphenyltetrazolium bromide assay
Cells were seeded in 96-well plates at 5,000 cells per well in 100 μL medium. After treatment described in legends of Figures 2A, 3A, 3F, and 4F, 10 μL 3-(4,5-dimethylthiazol-2-yl) -2,5-diphenyltetrazolium bromide (MTT) solution (Beyotime Biotechnology Co.) (5 mg/mL) was added to the cells, and cells were allowed to grow at 37°C for 4 h. MTT-containing medium was removed, and 100 μL MTT formazan dissolution was added, and cells were allowed to grow at 37°C for another 3–4 h. The optical absorbance was determined at 570 nm using a microplate reader (iMark; Bio-Rad Laboratories, Inc.).
Apoptosis and flow cytometry analysis
For analysis of the EpCAM-positive ratio, the fluorescein isothiocyanate (FITC)-conjugated anti-human EpCAM antibody (ab8666; abcam) was used. For apoptosis assays, cells were cultured in six-well plates and were harvested 72 h later. Then the apoptosis rate was analyzed using the Annexin V/PI or APC/7-AAD antibodies (BD Biosciences). Data were acquired on an FACSCalibur flow cytometer (BD Biosciences) and analyzed with CellQuest software.
Colony formation assay
One thousand two hundred cells per well were seeded in a six-well plate and treated as described in the legend of Figure 3C, after adherence by the next day. Two weeks later, cells were fixed with 4% paraformaldehyde and stained with crystal violet. After drying, we photographed the plates and Image Pro Plus software was applied to calculate the number of colony. Colony formation rates were expressed as the percentage of colonies in cultures compared with that in Ad-tBid/cisplatin cotreated cultures. The assay was performed in triplicate.
Caspase activity analyses
Cells were cultured in 96-well plates and treated as described in legend of Figure 4B. Caspase-3 activities were determined by the Caspase 3 Activity Assay Kit (Beyotime Biotechnology Co.) according to the manufacturer's protocols.
Measurement of mitochondrial membrane potential
The mitochondrial membrane potential assay kit with JC-1 (Beyotime Biotechnology Co.) was used to measure the potential of mitochondrial membrane. Cells were infected with Ads as indicated in legends of Figures 4A and 4E and cultured for 72 h. The slides were then washed with ice-cold PBS and incubated with JC-1 staining solution for 20 min at 37°C. After two washes with filtered PBS, the slides were observed for morphology with a confocal laser scanning microscope (true confocal scanner spectrophotometry; German Leica, Mannheim, Germany). The ratio of monomer (green) and J-aggregate (red) after JC-1 staining represents the degree of MOMP.
Immunohistochemistry assay
Mouse tumor tissues were fixed in 10% formalin overnight and embedded in paraffin. Then paraffin-embedded sections were deparaffinized in xylene. Immunohistochemistry (IHC) was carried out using an avidin/biotin complex Vectastain Kit (Zsgb-Bio, Beijing, China) following the manufacturer's protocols. Endogenous peroxidase was blocked by incubation with 0.3% hydrogen peroxide for 30 min at 37°C. Antigen retrieval was performed by boiling the slides in a citrate buffer water bath for 10 min. Slides were washed in PBST and blocked for 30 min with 5% BSA. Slides were incubated overnight at 37°C with primary antibodies, followed by 1 h with a labeled polymer-HRP secondary antibody at room temperature. Negative controls were treated with PBS, but without a primary antibody. Subsequently, slides were incubated with diaminobenzidine (DAB) dyestuff for 3–5 min. After fixation, slides were observed under a microscope and photographed. The following primary antibodies were used in the IHC assay: Bid (ab201754; abcam) and adenovirus fiber (ab3233; abcam).
Positive and negative controls were evaluated in each experiment to control for staining variability among batches of experiments. For semiquantitative evaluation of protein levels in tissues, an immunoreactivity-scoring system (H-SCORE) was used. Briefly, the staining intensity was graded (0, absence; 1, weak; 2, moderate; 3, strong). The H-SCORE was calculated using the following formula: H-SCORE = ∑Pi × i, where i is the staining intensity of tumor cells, and Pi is the percentage of corresponding cells at each level of intensity. Each data point represents the mean score of two pathologists, who were blinded to all clinicopathologic variables.
In vivo animal studies
Female BALB/c-nu mice (4–5 weeks old) were purchased from Beijing HFK Bio-Technology Co. Ltd (Beijing, China). All animal experiments were approved by the Ethics Committee of Animal Experiments of Tongji Medical College and all mice were housed under pathogen-free conditions at Tongji Medical College.
For the subcutaneous xenograft models, a total of 5 × 106 HOC7 cells diluted in 50 μL serum-free growth medium were implanted subcutaneously. The tumor volume was determined every 4 days using a digital caliper. After 2 weeks, when the tumor volume reached 200–250 mm3, 21 mice were stratified into six groups at random (3–4 mice per group) to receive 50 μL intratumoral (i.t.) injections of 1 × 108 pfu Ad-dE1A, Ad-tBid, or PBS for four times and subsequently cisplatin (5 mg/kg/day) was injected intraperitoneally (i.p.) every other day for three times. 28,29 Tumor volumes were estimated (volume = length × width 2 × 0.52) every 4 days until tumor volume reached 2,500–3,000 mm3 or tumor ulceration occurred. Six weeks after tumor cell inoculation (day [D] 42), mice were killed and tumor tissues were dissected immediately after death. Investigators were blinded to the treatment groups. The assay was performed twice.
For intraperitoneally disseminated xenograft models, 2.53 × 106 SKOV3 tumor cells were intraperitoneally inoculated. Tumor growth was dynamically monitored in living mice by optical imaging of luciferase activity using the IVIS SPECTRUM system (Caliper, Xenogen). After tumor establishment (∼21 days), the mice were randomized into six groups of 11 animals per group. Mice were cotreated with 100 μL PBS, 1 × 108 pfu Ad-dE1A, Ad-tBid by i.p. injection on D21, D23, D25, and D27, or cisplatin (5 mg/kg/day) by i.p. injection three times on D30, D32, and D34. Two weeks after the last injection (D48), three mice per group were killed and tissues dissected immediately after death.
Statistical analysis
Unless otherwise specified, statistical analysis was carried out using GraphPad Prism 6.0 and SPSS 19.0 software. Differences between groups were analyzed using the Student's t-test and one-way analysis of variance (ANOVA). p < 0.05 was considered significant. Results are presented as mean ± standard deviation.
Results
Construction and validation of oncolytic Ads Ad-Bim and Ad-tBid
Gathering of proapoptotic proteins in the outer mitochondrial membranes of cancer cells may render them primed to death and more sensitive to cytotoxic chemotherapy. A previous study reported that Bim and Bid peptides could induce mitochondrial priming in OC clinical samples, suggesting that Bim and Bid have the potential to induce mitochondrial depolarization in OC. 5 To investigate their potential as a novel strategy for OC, we used the oncolytic adenoviral vector system. First, we deleted the E1A CR2 and E3 ADP region from the Adv5 genome. Then, we inserted the EGFP cDNA into the E3 ADP region to generate a control-armed Ad (Ad-dE1A). The Bim and tBid (the active form of Bid) open reading frames were assembled into the E3 ADP region to generate the novel tumor-lysing Ad, respectively (Ad-Bim and Ad-tBid) (Fig. 1A).
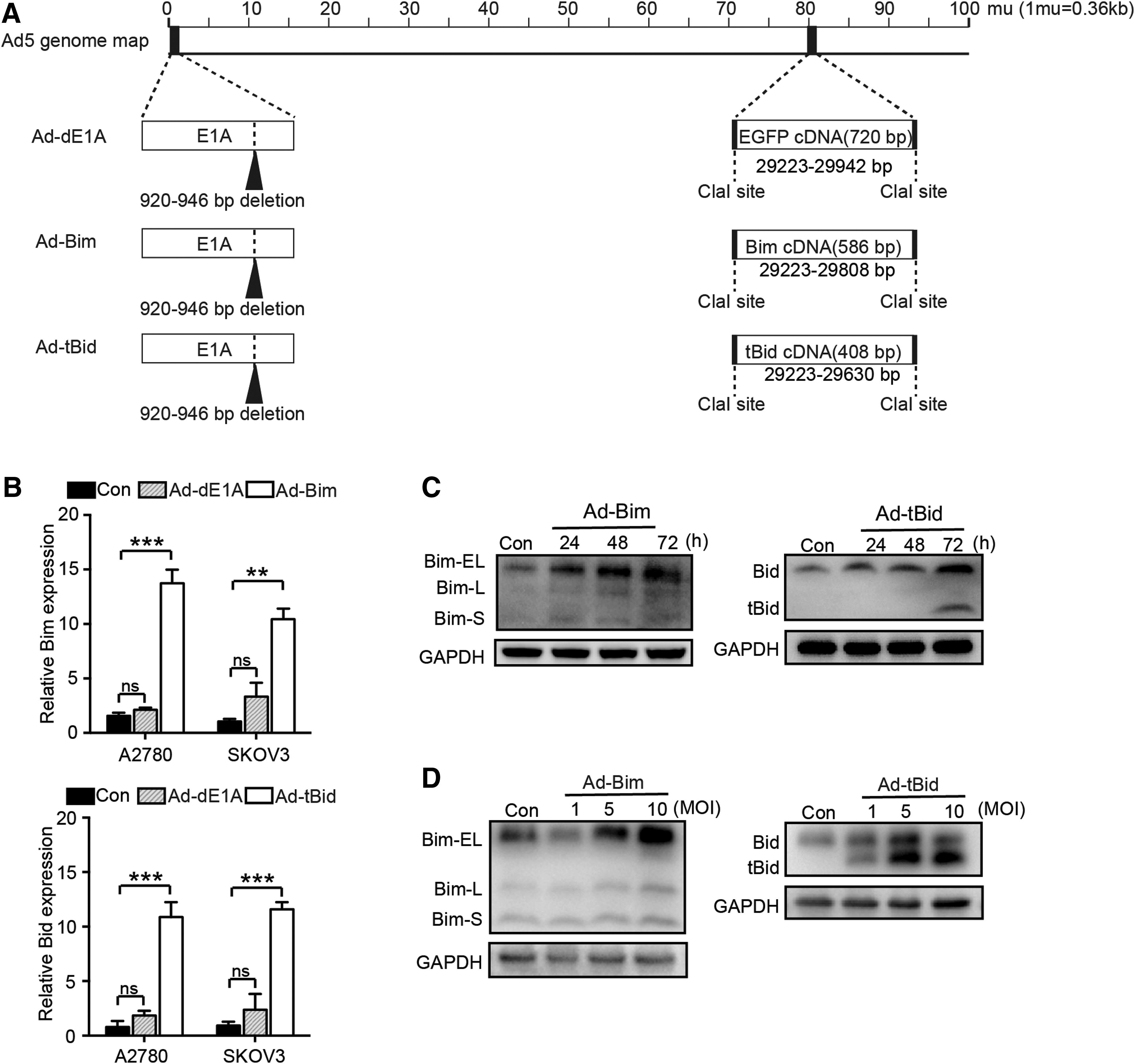
Recombinant Ad vector-mediated Bim/tBid gene transfer in OC cells.
To determine whether the original E3 promoter could drive the expression of the embedded transgene, reverse transcription-qPCR was used to detect the expression in infected A2780 and SKOV3 cells (Fig. 1B). The results indicated that cells infected with Ad-Bim or Ad-tBid significantly increased Bim or Bid mRNA expression compared with Ad-dE1A-infected cells (both p < 0.05). Subsequently, Western blot was used to further confirm the effects of recombinant Ad vector-mediated gene transfer in SKOV3 cells. As Bim/tBid protein could be detected even at a low MOI of 5 after 72 h of incubation, we set the MOI of 5 and the time point of 72 h as the optimal condition (Fig. 1C, D). Together, these results indicated that Ad-Bim and Ad-tBid can efficiently increase the expression of proapoptotic transgene in OC cells.
Ad-tBid displayed superior antitumor effects compared with Ad-Bim
Next, we evaluated the potency of Ad-Bim/Ad-tBid as a single agent for the lysis of cells in nine OC cell lines. As shown in Fig. 2A and B, all of the nine cell lines demonstrated high sensitivity to Ad-tBid. A majority of the cell lines (7/9), except for HOC7 and SKOV3, exhibited a significant decrease in cell viability to Ad-tBid infection even at a relatively low MOI (0.1–5). However, Ad-Bim-infected cells showed no obvious decrease in cell viability compared with Ad-dE1A-infected cells (Fig. 2A, B). Collectively, our results demonstrated that Ad-tBid exerted superior antitumor effects against OC cells compared with Ad-Bim.

Effect of Ad-Bim and Ad-tBid on survival in OC cells.
Ad-tBid and cisplatin synergistically inhibit proliferation of refractory OC
To explore whether Ad-tBid could enhance cisplatin treatment, we first evaluated the response to cisplatin in nine OC cell lines (Fig. 3A) and assessed the half-maximal inhibitory concentration (IC50) (Fig. 3B). Notable cell death was observed in most cells following incubation with cisplatin at relatively low concentrations (<10 μM). In contrast, only about 30–50% death rates were observed in HOC7 or SKOV3 cells after treatment with 10 μM cisplatin for 48 h. Thus, we recognized HOC7 and SKOV3 as cisplatin-refractory OC cell lines in this study.
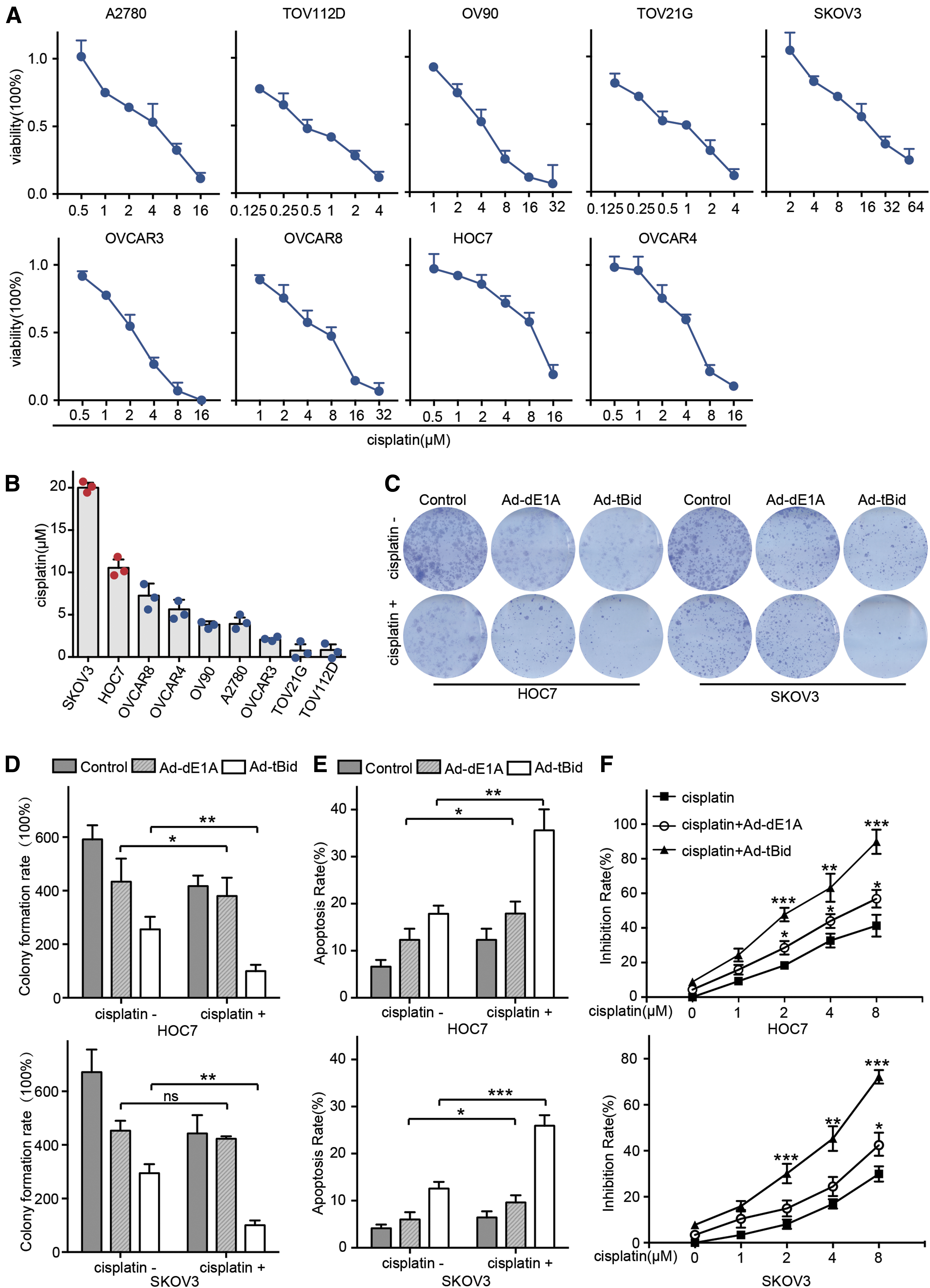
Ad-tBid enhanced the efficacy of cisplatin in refractory OC cells.
The colony formation assay showed that Ad-tBid combined with cisplatin yielded a strong inhibitory effect (Fig. 3C, D). Subsequently, the proapoptotic effect of Ad-tBid in combination with cisplatin on HOC7 or SKOV3 cells was measured. An MOI of 5 for Ad-tBid alone could induce about 18% and 12% cell death after 72 h of incubation in the two OC cells, respectively. Both cell lines failed to respond to cisplatin treatment at the concentration of 2 μM as the apoptosis rate was only 12% and 7%. Interestingly, pretreatment with Ad-tBid at an MOI of 5 led to obvious cell death in response to cisplatin (2 μM). In the Ad-tBid pretreated group, the apoptosis rate increased to 32% and 27% in HOC7 and SKOV3 cells, respectively (Fig. 3E). Consistent with this observation, the inhibition rate of 8 μM cisplatin increased approximately two to three fold in HOC7 and SKOV3 cells after incubation with Ad-tBid in the MTT assay (Fig. 3F). Together, these results suggested that Ad-tBid significantly increased cisplatin-induced apoptosis in OC cells.
Ad-tBid enhances cisplatin sensitivity by activating the mitochondrial apoptosis pathway
Since Bid-peptide pretreatment enhanced chemosensitivity to multiple agents by increasing mitochondrial priming, we hypothesized that Ad-tBid may increase the antitumor effects of cisplatin via a similar mechanism. MOMP is a necessary step to commit cancer cells to programmed cell death and a previous study developed a functional assay to measure it (referred to as BH3 profiling). In this assay, we use JC-1 staining to roughly calculate the degree of MOMP. Besides, we used Adv5/dE1A, an oncolytic Ad constructed previously, 19,22 as the control Ad because it contains no EGFP gene. As shown in Fig. 4A, HOC7 and SKOV3 cells incubated with Ad-tBid exhibited a significant increase in monomer and a decrease in J-aggregate. Besides, treatment with Ad-tBid resulted in obvious cell damage, characterized by nuclear swelling.
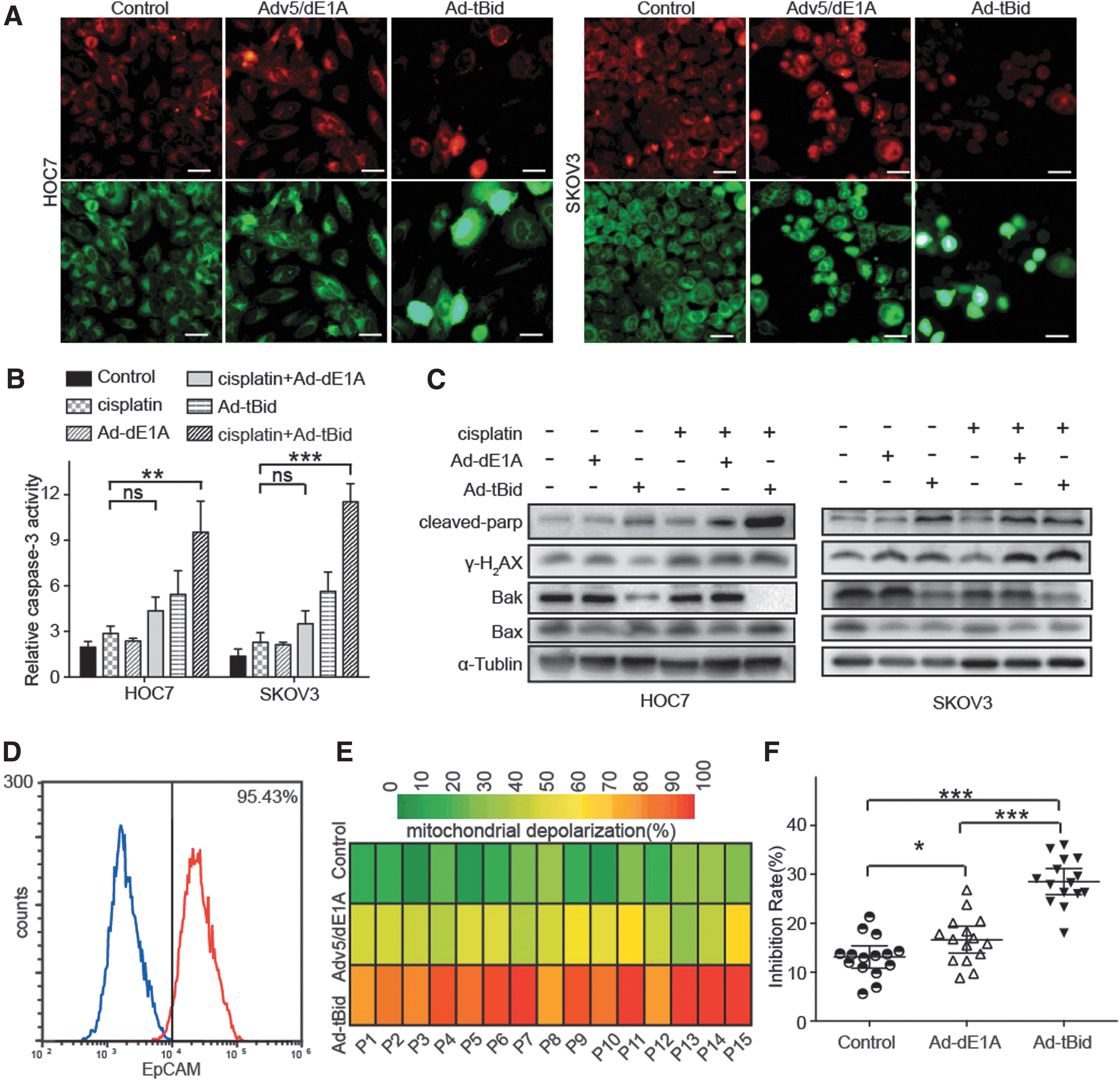
Ad-tBid in combination with cisplatin promoted mitochondrial apoptosis.
Next, we measured the levels of cleaved caspase-3 in the two cell lines. The levels of cleaved caspase-3 in both cells were significantly increased by Ad-tBid plus cisplatin treatment, indicating that cotreatment triggered cell death mainly through an apoptotic manner (Fig. 4B). Immunoblot further indicated that Ad-tBid plus cisplatin treatment led to significant activation of cleaved parp and γ-H2AX and decreased the expression of Bak, while the expression of Bax remained unchanged. Bak reduction may be due to the oligomerization of Bak as previously reported 30,31 (Fig. 4C).
To further verify the mechanism under Ad-tBid incubation, we separated primary OC cells from the ascetic fluid of 15 different OC patients. The purity of primary cells was verified with EpCAM (the epithelium-specific marker)-based flow cytometric analysis; only those with epithelial percentage >90% were used (Fig. 4D). In most cases, Ad-tBid incubation induced significant MOMP compared with Adv5/dE1A incubation (Fig. 4E). Besides, stronger MOMP by Ad-tBid incubation before treatment was strongly associated with better response to cisplatin (Fig. 4F). Altogether, these results indicated that Ad-tBid enhanced cisplatin sensitivity by activating the mitochondrial apoptosis pathway. Altogether, these results indicated that Ad-tBid pretreatment led to increased priming of OC cells and made them more susceptible to cisplatin-induced apoptosis.
Ad-tBid synergizes with cisplatin to inhibit tumor progression in vivo
To investigate whether the effects of the combination of Ad-tBid and cisplatin were also achievable in vivo, mice bearing palpable HOC7 tumors were treated with (1) i.t. phosphate-buffered saline, Ad-dE1A, or Ad-tBid injection, (2) intraperitoneal cisplatin injection, or (3) a combination (Fig. 5A). In keeping with the in vitro observations, the combination of Ad-tBid and cisplatin resulted in smaller tumors than the other five groups (Fig. 5B, C). Moreover, Ad-tBid injection alone also resulted in a remarkable reduction in tumor growth compared with PBS treatment. Meanwhile, the average tumor weights were calculated and indicated that Ad-tBid plus cisplatin treatment efficiently inhibited tumor progression (Fig. 5D).
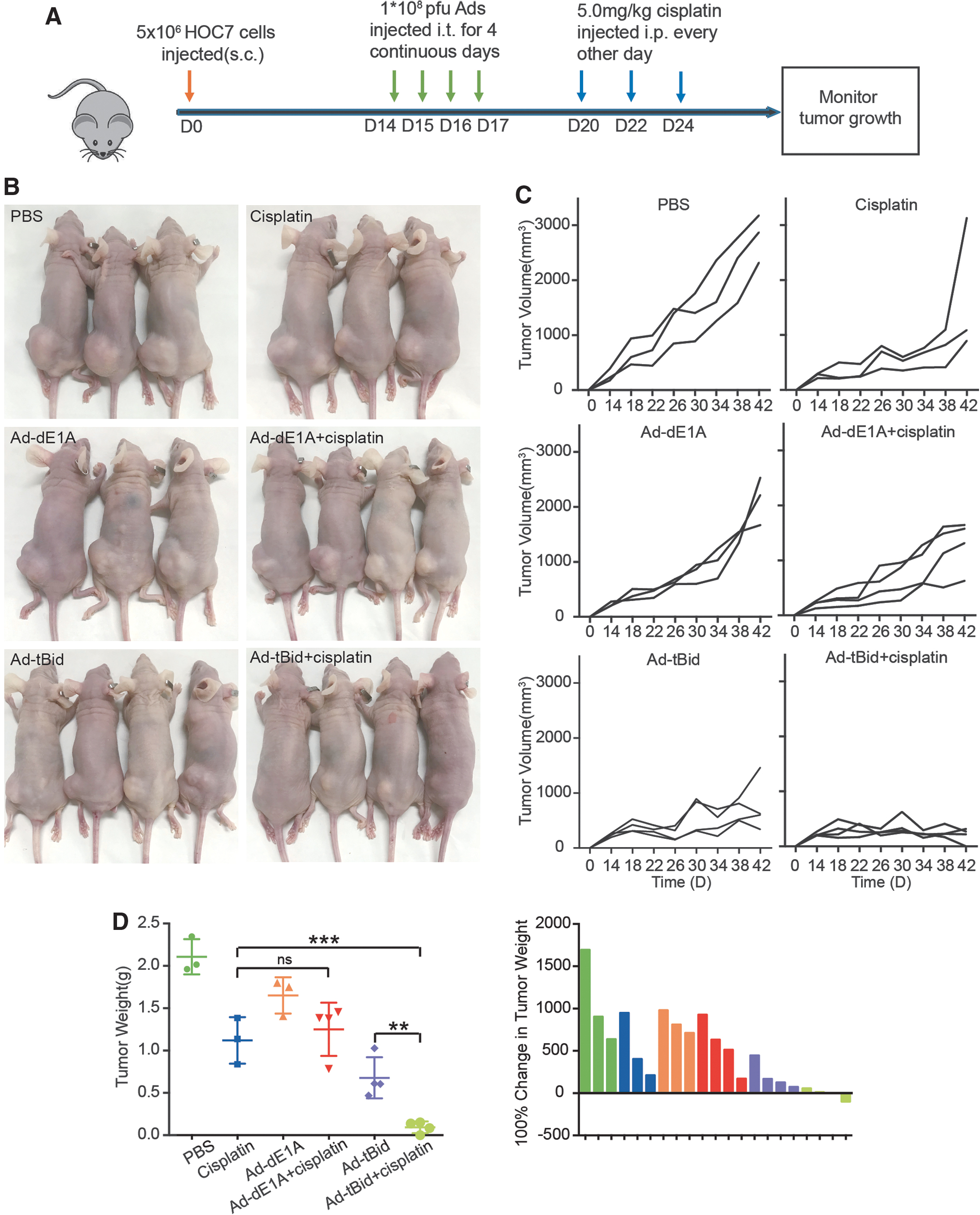
Ad-tBid significantly enhanced the antitumor effects of cisplatin in vivo.
Ad-tBid/cisplatin can safely cure disseminated OC in vivo
To further evaluate the antitumor activity of cotherapy in advanced OC, a peritoneal disseminated OC model that rapidly recapitulates late stage was used. We visualized the growth of individual peritoneal metastases by in vivo imaging system (IVIS) in the same mouse at four time points (0, 7, 14, and 21 days after first injection) (Fig. 6A). Consistent with the earlier results, cisplatin alone had minimal effects on tumor growth, but combination with Ad-tBid therapy markedly reduced the tumor burden in xenograft models (Fig. 6B). The combination of Ad-tBid and cisplatin significantly inhibited tumor growth and delayed tumor progression as mice that received combination treatment showed a 62.5% survival rate, which persisted until the experiment was terminated (Fig. 6C).
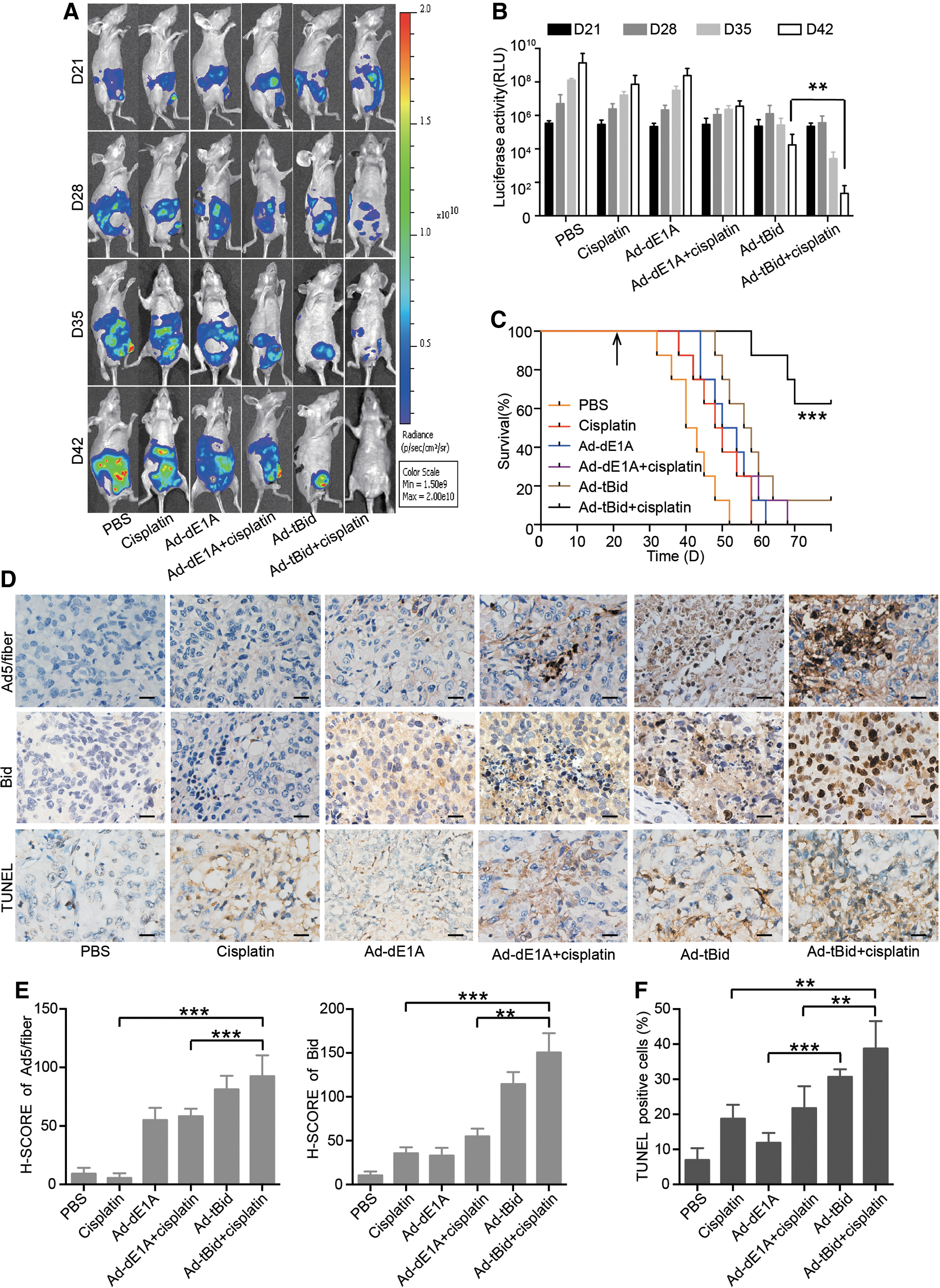
Combinatorial cisplatin and Ad-tBid treatment is efficacious in vivo.
Tumor sections were further examined in xenografts by immunohistochemical staining. As shown in Fig. 6D–F, Ad-tBid in combination with cisplatin increased apoptosis as evidenced by increased terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) staining. Moreover, the replication of Ads and Bid expression was also increased in the combined group (Fig. 6D–F). Together, these data demonstrated that the novel oncolytic Ad Ad-tBid could significantly enhance the antitumor effects of cisplatin in vivo.
Discussion
Despite extensive efforts, the prognosis of patients with OC continues to be poor. In this study, we demonstrated that Ad-tBid was a therapeutic agent that increased the efficacy of cisplatin in vitro, in vivo, and ex vivo. Ad-tBid or cisplatin alone induces only minor tumor death; their cotherapy has a substantial killing effect. The mechanisms underlying the effect have dual functions: (1) increasing the expression of proapoptotic protein in refractory tumor cells and achieving more sensitivity to cisplatin by inducing mitochondrial priming and (2) Ad-tBid serves as an oncolytic Ad that could replicate itself in cancer cells and the progeny viruses cause a cytopathic effect.
Refractoriness to conventional chemotherapy is a major challenge in the treatment of advanced OC. There is increasing evidence that the degree of mitochondrial apoptotic priming determines whether a cell will undergo apoptosis after chemotherapy treatment. 1 However, the regulation of mitochondrial apoptotic priming depends on many factors; both tumor microenvironment and cell lineage are strong modulators. 32 –34 The most critical factor in determining mitochondrial priming is the modification of pro- and antiapoptotic proteins of BCL-2 family that have proceeded during tumorigenesis. For instance, a cell expressing large amounts of antiapoptotic proteins can survive a substantial amount of death signaling, including chemotherapy treatment. Alternatively, a cell loading rich proapoptotic proteins would not be able to weather even a small stressor. A cell that has a huge reserve of proapoptotic proteins is “primed” for apoptosis, whereas those with smaller reserves are “unprimed.” 7 When these two cells are treated with equal doses of chemotherapy, the primed cell seems to be more likely to trigger apoptosis than the unprimed cell. In addition, tumor cells that have gone through standard treatment and then recurred are usually less primed, making them insensitive to subsequent chemotherapy. This may also be a reasonable explanation for the higher intractability of recurrent tumors. 35
Although mitochondrial priming plays a prominent role in chemotherapy, it is unclear how to exploit it for cancer therapy. Recently, a novel class of agents called BH3 mimetics, specifically targeting the antiapoptotic proteins of the BCL-2 family, has already shown promise especially in terms of the treatment of hematological malignant tumors. The first approved BH3 mimetic was ABT-263 (navitoclax), which inhibits BCL-2, BCL-XL, and BCL-w and potently induces apoptosis in several solid and hematological cancers. 36 Then, the BCL-2-specific inhibitor venetoclax (also known as ABT-199) was approved by the Food and Drug Administration for the treatment of refractory chronic lymphocytic leukemia. 37 Recently, BH3 mimetics specifically targeting BCL-XL (A-1331852) and MCL-1 (S63845) have also been developed. 38,39 However, only few successful stories in solid cancers have been reported. Besides, their efficacy has been limited by serious side effects, such as thrombocytopenia. 37
Here, we used another interesting strategy to get that turned around. We armed oncolytic Ads with proapoptotic proteins directly inducing activation of proapoptotic effector Bak/Bax to overcome insensitivity to chemotherapy. Our results indicated that Ad-tBid alone could induce cancer cell death more effectively compared with Ad-Bim. Importantly, the therapeutic window of cisplatin will be extended when combined with Ad-tBid as Ad-tBid could significantly enhance cisplatin-induced apoptosis both in vitro and in vivo. Similarly, when tested on clinical samples, primary OC cells were more sensitive to cisplatin treatment after Ad-tBid administration. Moreover, Ad-tBid combined with cisplatin promoted proximity of tumor cell mitochondria to the apoptotic threshold and Bak oligomerization to kill cancer cells. Of particular note, as Bim peptide was reported to have the potential of inducing mitochondrial priming in multiple myeloma, AML, ALL, and OC, 5 Ad-Bim showed little antitumor effect in OC. Besides, no signaling indicating mitochondrial priming could be investigated in OC after Ad-Bim treatment (data not shown), although overexpression of Bim after Ad-Bim incubation was indeed confirmed. One possible reason is that different drug carriers determine diverse therapeutic windows. Besides, Bim probably requires a larger protein load to induce mitochondrial priming compared with Bid.
Previous studies have involved the insertion of Bid or tBid into Ad, and the powerful efficacy was affirmed in nonsmall-cell lung cancer and hepatocellular carcinoma, respectively. 40,41 However, both of them lack the selectivity to specifically target tumor tissues. Thus, we inserted tBid cDNA into the backbone of new-generation selectively oncolytic Ad to generate Ad-tBid, and confirmed its cytotoxic efficacy in various OC cells. All nine cancer cell lines that were examined showed a marked decrease in viability after exposure to Ad-tBid.
Although we demonstrated a new strategy for combining oncolytic virotherapy with chemotherapy, some limitations of our study need to be acknowledged. First, Ad-tBid is only applicable to superficial tumors, which allow for i.t. injection. Off-targeting, especially liver sequestration, and immune clearance of Ads are major problems that limit its application after systemic or intraperitoneal injection. Second, more precise and comprehensive toxicological and pharmacological experiments are required to assess the safety associated with Ad-tBid administration. Further assessments of these aspects are warranted before translation of this therapy into early-stage clinical trials.
Herein, we show that the administration of Ad-tBid can increase the priming of cancer cells and thus makes other, concurrently administered chemotherapies more effective. Besides, tBid-based virion enhanced cisplatin sensitivity in various tumor cells by mito-priming and we confirmed its effect in two mouse models. In summary, our findings imply that Ad-tBid-cisplatin combination treatment is a potential therapeutic strategy for chemoresistant OC.
Footnotes
Acknowledgments
We thank Sen Xu and Meng-Chen Li for technical help in data analysis; we also thank Ze-Yu Zhang and Yu Xia for help in article preparation.
Author Disclosure
No competing financial interests exist.
Funding Information
This study was supported by the National Science Foundation of China (grant nos. 81802608, 81772787, and 81572570), the National Science and Technology Major Subproject (2018ZX10301402-002), and the “973” Program of China (grant no. 2015CB553903).