
Abstract
High-risk human papillomavirus (HPV) E6 and E7 genes display vital oncogenic properties in cervical cancer. Eliminating HPV driver gene or loss of function by the clustered regularly interspaced short palindromic repeat (CRISPR)/Cas9 system is a promising treatment for the HPV-associated cancer. Thus, this study designed a CRISPR/Cas9 system to target the E6 and E7 genes at once, to detect whether it have efficacy in vitro and in vivo. Meanwhile, CRISPR/Cas9 system was measured after transfection with liposomes but virus. Cervical cancer lines (HeLa and SiHa) were used in this study. Sanger sequencing confirmed that the single CRISPR/Cas9 vector [termed E6E7-knockout (KO)] containing guide RNAs could targeting both HPV18 E6 and E7 genes in vitro. In addition, double-targeting E6 and E7 increased p53 protein expression significantly while compared with E6 or E7 targeting, respectively. Mice with xenografts were divided into four groups: three doses of experimental groups (20, 40, and 60 μg) and one control group. The E6E7-KO through liposome delivery was injected into tumors. Tumor growth was measured and protein expression was observed through immunohistochemistry. The toxic side effects in vivo were also evaluated. E6E7-KO induced cell apoptosis and inhibited cell proliferation markedly in vitro. E6E7-KO downregulated the messenger RNA and protein expression of E6 and E7, whereas p53 and p21 protein levels were upregulated accordingly. Notably, E6E7-KO delivery by liposome exhibited an effect in vivo. Tumor growth was inhibited in the E6E7-KO groups, which was accompanied by decreased E6/E7 protein expression and increased p53/p21 protein expression, especially the level of p53 protein expression. Therefore, E6E7-KO could have synergy efficient by p53 pathway. Furthermore, local injection with CRISPR/Cas9 by nonviral delivery may be regarded as a potential therapy for cervical cancer in the future.
Introduction
Cervical cancer is mainly attributed to persistent infection with high-risk types of human papillomavirus (HPV). An accumulating body of evidence has indicated that E6 and E7 serve crucial roles in the occurrence and progression of cervical cancer. The two viral genes E6 and E7 have been revealed to have oncogenic properties and maintain the malignant phenotype. 1 They regulate the viral life cycle, exploit the host DNA replication machinery to replicate its genome, and stimulate chromosomal instability and DNA damage. 2 The dominant mechanism of E6 action is the degradation of p53, and E7 inactivates retinoblastoma family proteins (pRB) and interacts with the downstream protein p21 and other pathways. 3 –5 In addition, the two driver genes not only induce the immortalization and transformation of cancer cells but also suppress the innate immune response. 6,7 In this regard, knocking out E6 and E7 may be valuable for cervical cancer treatment.
HPV-associated cervical cancer offers a perfect model for clustered regularly interspaced short palindromic repeat (CRISPR)/Cas9 therapies. E6 and E7 are sequence-specific genes that can be discriminated from human genes and are expressed exclusively. 8 Notably, E7 contains genetic conservation, which is critical to carcinogenesis. 1 Small interfering RNA (siRNA) and short hairpin RNA inhibitors could silence the two genes and had an effect in vitro. It was reported that the combination of siRNA anti-E7 and antimyeloid cell leukemia sequence-1 exhibited high efficacy. 9 It was also argued that these approaches have no persistent function. Transcription activator-like effector nucleases disrupted E6 or E7 in vitro. 10 However, the procedure is complex. In recent years, CRISPR/Cas9 has revolutionized the field of gene editing 11 and has been considered for more clinical applications. 12,13 E6 and E7 knockout (KO) using CRISPR/Cas9 has produced promising results for HPV-positive cervical cancer. 8 In addition, CRISPR/Cas9 is required to reduce undesired off-target effects.
Regarding delivery vehicles, adenoviral and adeno-associated viral systems have exhibited good activity in vivo in recent years, 14,15 but their potential immunogenicity should be considered. 16 Despite its efficiency, nonviral delivery has become a new treatment strategy as it can partly avoid the side effects caused by viral delivery. In fact, nonviral delivery of the CRISPR/Cas9 system should be fully explored. Gene editing in vivo requires the optimization of the delivery system, in which the concentration and transfection material are important. 17 To the best of our knowledge, for cervical cancer, the CRISPR/Cas9 system by nonviral delivery has not been detected in vivo yet.
In this study, a single CRISPR/Cas9 vector (viral vector) was constructed and adapted to target the HPV18 E6 and E7 genes through nonviral delivery, leading to the loss of function of the two genes. The effect of E6E7-KO was proven in vitro at first. Next, E6E7-KO mixed with Lipofectamine 3000™ (termed L-3000) was injected into mouse tumors locally. As the majority of cervical cancers can be observed vaginally, it was thought that local injection therapy would be feasible and reasonable. Hence, the aim of this study was to detect its function in vitro and in vivo. Local gene therapy might be helpful and advantageous for the better management of cervical cancer treatment.
Materials and Methods
Plasmid construction and extraction
Plasmid p330x encoding Cas9 and enhanced green fluorescent protein (EGFP) with puromycin (puro)-flag was kindly provided by Cyagen Biosciences, Inc. (Santa Clara, CA). We designed four single guide (sg)-RNAs targeting the HPV18 E6 and E7 genes. The sgRNAs were cloned and inserted into the p330x-plasmids. The transfect efficacy was observed, and then, HPV18 E6 (GAAGCTACCTGATCTGTGCA) and HPV18 E7 (GGAGCAATTAAGCGACTCAG) were chosen to establish a single vector. The plasmid E6E7-KO was produced and verified by T7E1 assays and endonuclease enzyme digestion. Plasmids were transformed into lico and stored long term at −80°C. The negative control (NC)-vector with nonsense sgRNA was regarded as the control. Plasmids were amplified according to the manufacturer's instructions (Cyagen Biosciences, Inc.), and were then extracted and purified (Tiangen Biotech Co., Ltd., Beijing, China).
Cell culture and transfection
The HPV18-positive HeLa and HPV16-positive SiHa cervical cancer cell lines were purchased from the Shanghai Cell Collection (Shanghai, China). The cells were cultured in 1640 modified medium (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA) supplemented with 10% fetal bovine serum (Gibco; Thermo Fisher Scientific, Inc.) and 1% penicillin/streptomycin (HyClone; GE Healthcare Life Sciences, Logan, UT) in a humidified 5% carbon dioxide chamber at 37°C. The cells were cultured in six-well plates overnight and transfected at 80% cellular confluency. The plasmids (2 or 4 μg of E6E7-KO/NC) were added to each well of six-well plates, mixed with Lipofectamine 3000 (Thermo Fisher Scientific, Inc.) and Opti-Minimum Essential Medium (Opti-MEM)-1 Reduced serum Medium (Thermo Fisher Scientific, Inc.). The procedure was performed according to the manufacturer's instructions (Lipofectamine 3000 7.5 μL + p3000 7 μL+ Opti-MEM 250 μL). After 3 days, DNA as extracted for Sanger sequencing and the pictures post-transcription were captured using a microscope system (Olympus Corporation, Tokyo, Japan).
Flow cytometry
This study performed FACS (FACSAria II; BD Biosciences; Becton, Dickinson and Company, Franklin Lakes, NJ) to measure the transfer efficiency according to EGFP expression at 3 days post-transfection. In addition, cells were collected and double stained with Allophycocyanin Annexin V and 7-amino actinomycin D (7-AAD) staining (Apoptosis Detection Kit; BD Biosciences; Becton, Dickinson and Company) according to the manufacturer's instructions. In brief, the cells were trypsinized, washed twice with phosphate buffered saline, put into Annexin V binding buffer, and stained at room temperature with Annexin V for 15 min, then 7-AAD for 10 min. The rates of apoptosis were analyzed using a FACS Calibur system (BD Bioscience; Becton, Dickinson and Company) to calculate the induced cell death. The data were analyzed using BD Cell Quest software (BD Bioscience; Becton, Dickinson and Company).
RNA extraction and reverse transcription-quantitative PCR
Total cellular messenger RNA (mRNA) was extracted after transfection for 3 days using a RNeasy Mini kit (Qiagen, Inc., Valencia, CA). Complementary DNA was produced by Reverse Transcription-PCR (Takara Bio, Inc., Otsu, Japan) and was mixed with SYBR Green (Takara Bio, Inc.), following the manufacturer's protocol. Reverse transcription-quantitative PCR (RT-qPCR) was performed using the Bio-Rad System (Bio-Rad Laboratories, Inc., Hercules, CA). RT-qPCR was conducted with the following thermocycling conditions: 40 cycles of heating at 95°C for 15 s, 58°C for 15 s, and 72°C for 20 s. Reactions were followed using melting curve analysis. The mRNA levels of E6 and E7 were normalized to the reference gene GAPDH. The following primer sequences were obtained from Invitrogen (Thermo Fisher Scientific, Inc.): HPV18-E6 forward 5′-ATCCAACACGGCGACCCTACAA-3′ and reverse 5′-CTGGATTCAACGGTTTCTGG-3′; HPV18-E7 forward 5′-ACCTTCTATGTCACGAGCAAT-3′ and reverse 5′-CGGACACACAAAGGACAGGGT-3′; HPV16-E6 forward 5′-AATGTTTCAGGACCCACAGG-3′ and reverse 5′-TCAGGACACAGTGGCTTTTG-3′; and HPV16-E7 forward 5′-ATGCATGGAGATACACCTACATTGC-3′ and reverse 5′-ACAATTCCTA GTGTGCCCATTAACA-3′.
Western blotting
This study performed western blotting to detect the protein expression levels of E6, E7, P53, P21, and B cell lymphoma 2 (BCL2) in the treated SiHa and HeLa cells. First, E6E7-KO/NC/E6-KO/E7-KO (2 or 4 μg/mL) were placed into six-well plates. After 3 or 5 days of transfection, the cells were harvested, washed, and lysed on ice for 30 min. The protein extracts mixed with loading buffer were subjected to electrophoresis on sodium dodecyl sulfate–polyacrylamide gel electrophoresis gels (Bio-Rad Laboratories, Inc.) and were transferred onto membranes, which were blocked and incubated with primary antibodies overnight. Next, the membranes were washed, incubated with secondary antibody and then washed again. The protein bands were detected and captured using the Bio-Rad System (Bio-Rad Laboratories, Inc.). The anti-HPV16/18 E6 (Abcam, Cambridge, UK), anti-HPV18 E7 (Abcam), anti-p53 (Abcam), and anti-p21 (Abcam) were used. The anti-GAPDH (Abcam) and anti-β-actin (Abcam) were used as the loading controls. Secondary antibodies were also purchased from Abcam.
Immunofluorescence analysis
Cells were cultured in 24-well plates that were loaded with glass slides and allowed to attach for 24 h. Then, the cells were cultured for 12 h and fixed with 4% paraformaldehyde for 30 min and with 0.3% Triton (Sigma Aldrich; Merck KGaA, Darmstadt, Germany) for 10 min. After blocking with 10% bovine serum albumin (Sigma Aldrich; Merck KGaA), the slides were incubated with the anti-E6 (1:100) and anti-E7 (1:100) overnight at 4°C and then with the secondary antibody Alexa Fluor® 594 (1:100) for 1 h at room temperature. The slides were then treated with 4’,6-diamidino-2-phenylindole (1:1,000) for 15 min, washed again, and mounted before examination. Immunofluorescence was detected using a confocal microscope (Carl Zeiss AG, Oberkochen, Germany).
Cell proliferation assay
Cell proliferation was observed using a Cell Counting Kit-8 (CCK-8; Dojindo Molecular Technologies, Inc., Kumamoto, Japan). HeLa cells transfected with E6E7-KO were screened using 0.75 μg/mL of puro for 3 days. Next, 1 × 103 cells were seeded per well of 96-well plates. All experiments were performed at 24, 48, 72, 96, and 120 h. CCK-8 was added at 10 μL per well. After 3 h of incubation at 37°C, the cells were measured with an instrument (Bio-Rad Laboratories, Inc.).
iCELLigence
Cell proliferation in vitro was investigated with iCELLigence software (ACEA Biosciences, Inc., San Diego, CA). Hela cells were transfected with E6E7-KO/NC vector as aforementioned. After 3 days, an equal number of cells (5 × 103cells) were placed into the wells of iCELLigence. The growth of the cells was monitored in the wells for 72 h.
Immunohistochemical staining
The tumors were embedded in paraffin. Immunohistochemistry (IHC) was performed. The slides were incubated overnight at 4°C with the anti-HPV16/18 E6 (Abcam), anti-HPV18 E7 (Abcam), anti-p53 (Abcam), or anti-p21 (Abcam). Antibody detection was performed using diaminobenzidine (DAB). Images were captured using an Olympus camera system.
Animal experiments
Next, 5-week-old nude mice (n = 15) were inoculated with 1 × 107 HeLa cells in the dorsal region of the subcutaneous tumor. When the xenograft reached ∼10 mm, the mice were divided into four groups. A total of three experimental groups were used, including E6E7-KO 60 μg (high dose), 40 μg (middle dose), and 20 μg (low dose) liposome nanoparticles concentration. In addition, there was one control group: the 60 μg NC group. There were four mice in each of the E6E7-KO dose groups and three in the control group. E6E7-KO (60, 40, or 20 μg) were mixed with liposome nanoparticles (Lipofectamine 3000 90 μL + p3000 120 μL; Lipofectamine 3000 60 μL + p3000 80 μL; Lipofectamine 3000 30 μL + p3000 40 μL, respectively). In addition, 60 μg of NC-Vector (Lipofectamine 3000 90 μL + p3000 120 μL) was used as control. Then, these mixtures were injected into the tumor tissues locally. Thereafter, mice weights and tumor sizes were measured, and the tumor volumes were calculated as follows: (length × width 2 ) × 1/2. The mice were sacrificed after 9 days. IHC was then performed. The mice organ weights were also measured.
Statistical analysis
All data are presented as the mean ± standard deviation. Each experiment was performed in triplicate. Data analysis was performed using SPSS 17.0 (SPSS, Inc., Chicago, IL). Student's t-test, with two-sample unequal variance, was used for analysis. p-Value <0.05 was considered to indicate a statistically significant difference.
Results
Construction and verification of E6 and E7 CRISPR/Cas9 plasmids
The four sgRNAs for the HPV18 sequence-specific E6 and E7 genes were identified from the literature
8
and designed using website (
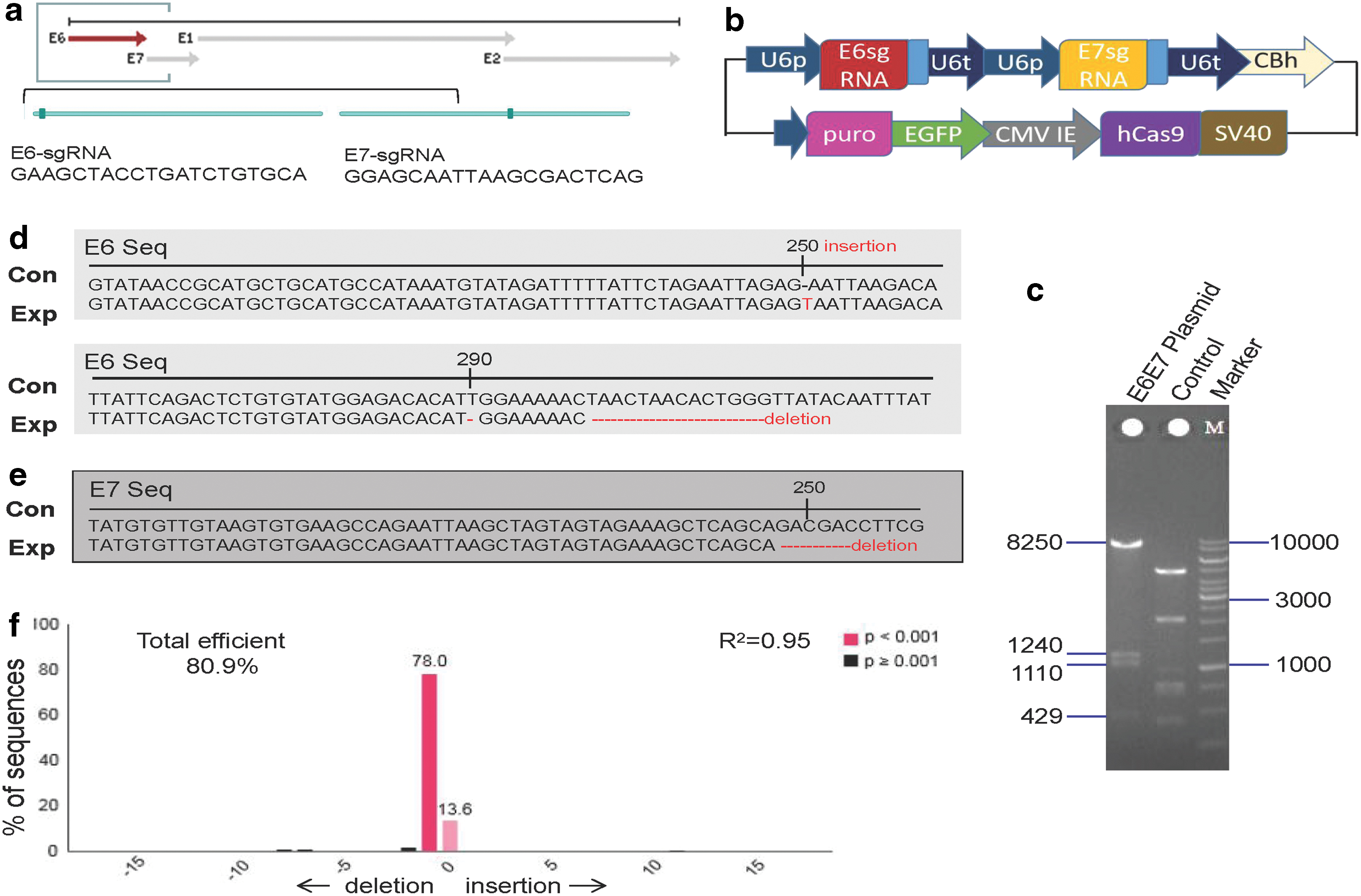
Construction of a single E6 and E7 CRISPR/Cas9 Plasmid and the gene editing efficacy.
Transfection with E6E7-KO induces cell apoptosis and inhibits cell proliferation in HeLa cells
Cells with green fluorescent protein (GFP) accounted for ∼20–30% after transfection for 3 days. Cells with GFP accounted for ∼80% when selected by Puro screening for 2 days (Fig. 2a). HeLa cells transfected with E6E7-KO exhibited marked cell death from 3 to 5 days, whereas SiHa cells exhibited little cell death (Fig. 2b, c). Notably, FACS revealed that the transfection ratios were parallel in HeLa or SiHa cells. There were no significant differences in transfection efficiency, 25% versus 20%, respectively (Fig. 2d).

E6E7-KO transfection rate and the cell viability.
To further verify the difference in the cell viability between the two cervical cancer cell lines, cell apoptosis was measured by FACS after transfection for 3 days. E6E7-KO exhibited increased apoptosis levels when compared with the NC group. By contrast, SiHa cells revealed no significant changes (Fig. 3a, b). In addition, subsequent experiments were performed to evaluate cell proliferation in vitro. The CCK-8 results illustrated that the E6E7-KO cells grew slowly, whereas the untreated cells developed rapidly (p < 0.01; Fig. 4a). Similar to the CCK-8 results, as shown in Fig. 4b, the cell numbers had markedly decreased in the E6E7-KO group when compared with those in the untreated group. iCELLigence methods also revealed the same results (Fig. 4c). Thus, it was believed that E6E7-KO could induce cell apoptosis and inhibit cell proliferation.

CRISPR/Cas9 E6E7-KO cells induce cell apoptosis in HeLa cells.
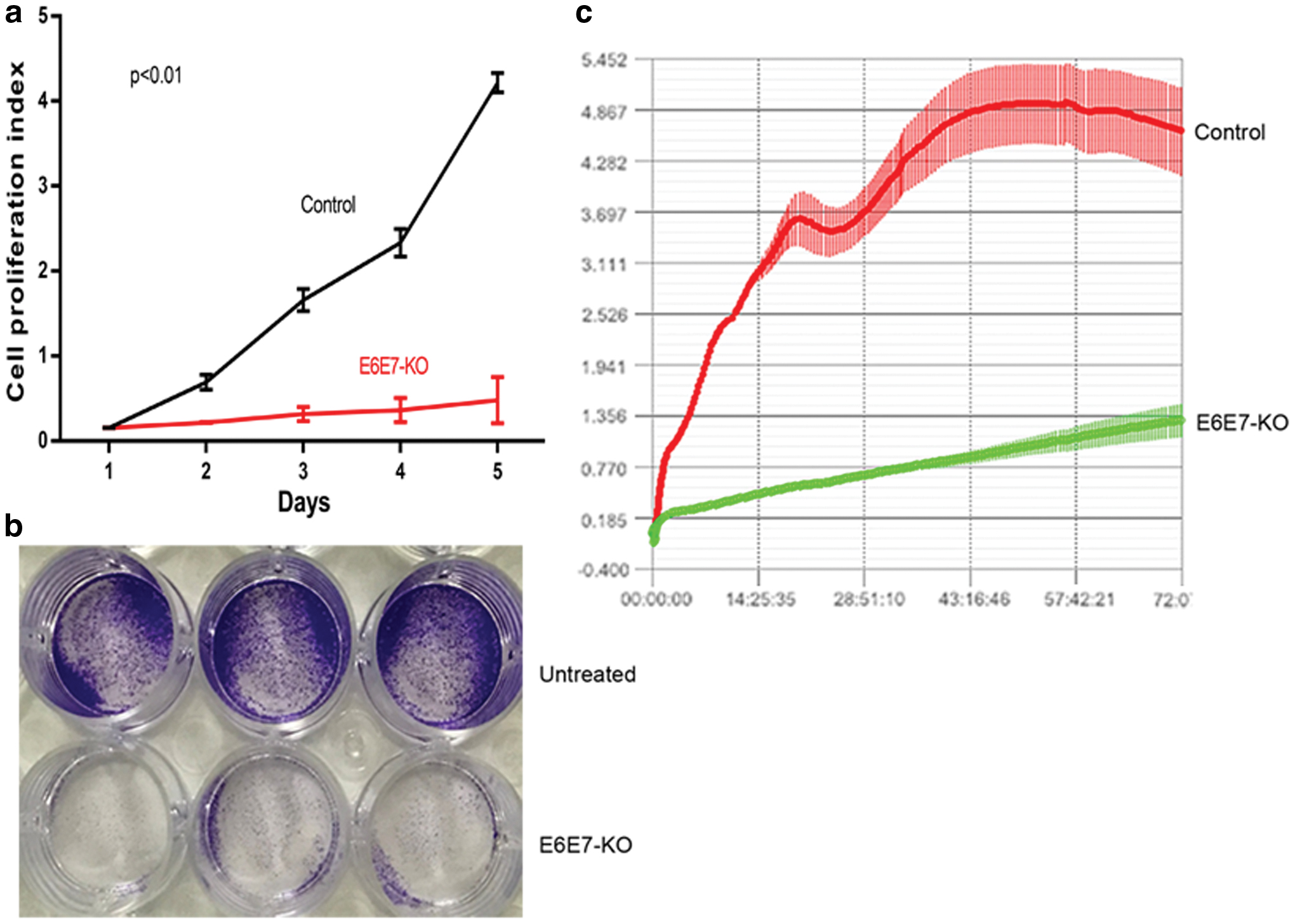
CRISPR/Cas9 E6E7-KO vector inhibits cell proliferation in vitro.
CRISPR/Cas9 E6E7-KO targets E6 and E7 efficiently in vitro
To confirm that E6E7-KO could inactivate or abrogate the function of E6 and E7 genes in HeLa cells, E6 and E7 mRNA levels were detected after transfection for 3 days. E6 and E7 mRNA levels were markedly decreased in the E6E7-KO groups in HeLa cells (p < 0.05; Fig. 5a). In contrast, in the SiHa cells, E6 and E7 mRNA levels were not changed when compared with those of the control group (Fig. 5b). By contrast, the protein levels of p21 and p53 were increased synchronously in the 2 and 4 μg E6E7-KO groups in HeLa cells when compared with the control and untreated groups (Fig. 5c). Therefore, E6E7-KO decreased E6 and E7 mRNA expression, leading to p21 and p53 upregulation correspondingly. Meanwhile, E6E7-KO increased p53 protein expression significantly compared with E6-KO or E7-KO (Fig. 5d).

E6 and E7 expression decrease when transfection with E6E7-KO.
To further validate the specific effect of E6E7-KO targeting E6 and E7 in HeLa cells, this study investigated the protein levels of E6 and E7 by western blotting and immunofluorescence of cells (ICC). For ICC, E6 and E7 levels in the HeLa cells (at 5 days) treated with E6E7-KO were markedly reduced (Fig. 6a, b). Compared with the control, the fluorescence value of the E6 protein declined in the E6E7-KO group, and E7 exhibited the same trend (Fig. 6c). Furthermore, the western blotting results indicated that the protein expression levels of E6 and E7 were decreased after transfection for 3 and 5 days. In addition, 2 or 4 μg of E6E7-KO exhibited the same tendency (Fig. 6d). In conclusion, E6E7-KO could decrease the effect of E6 and E7 gene and reverted the p53 function in vitro.
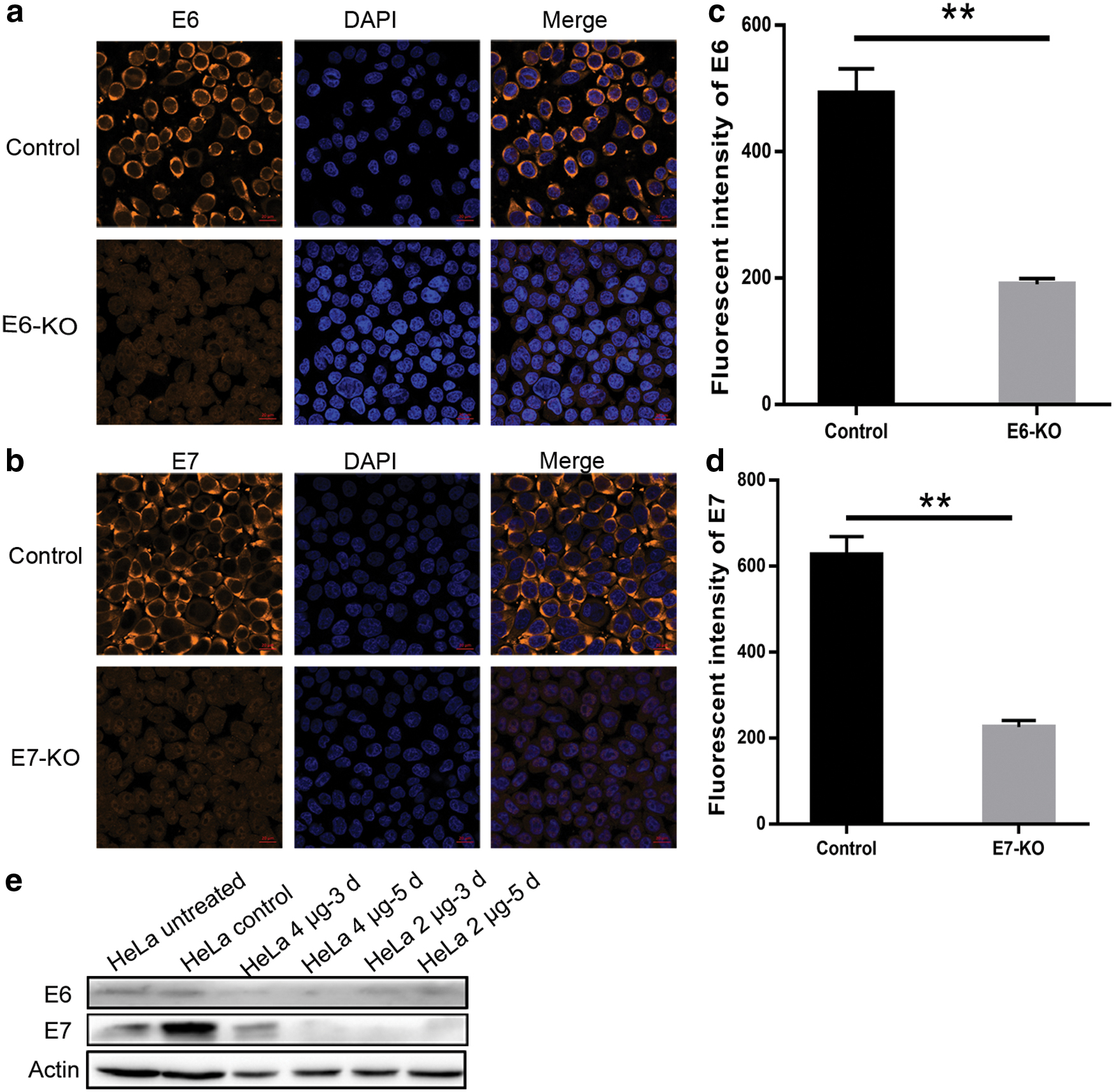
CRISPR/Cas9 E6E7-KO vector targets E6 and E7 efficiently in vitro.
CRISPR/Cas9 E6E7-KO with nonviral transfection suppresses tumor growth in vivo
To detect whether E6E7-KO would inhibit tumor growth in vivo and the effect of nonviral delivery, different doses of liposome nanoparticles were injected into tumors. The tumor size in the experimental groups was significantly smaller than the NC group. Among the experimental groups, the tumor size in the high-dose group was slightly smaller than that of the middle- and low-dose groups (Fig. 7a). In terms of the tumor weight, those of the experimental groups were lighter than the control group (p < 0.01). Nevertheless, there were no significant differences among the experimental groups (Fig. 7b). As shown in Fig. 7c, the tumor growth rates in the experimental groups were slower than the control group, especially in the high-dose group.
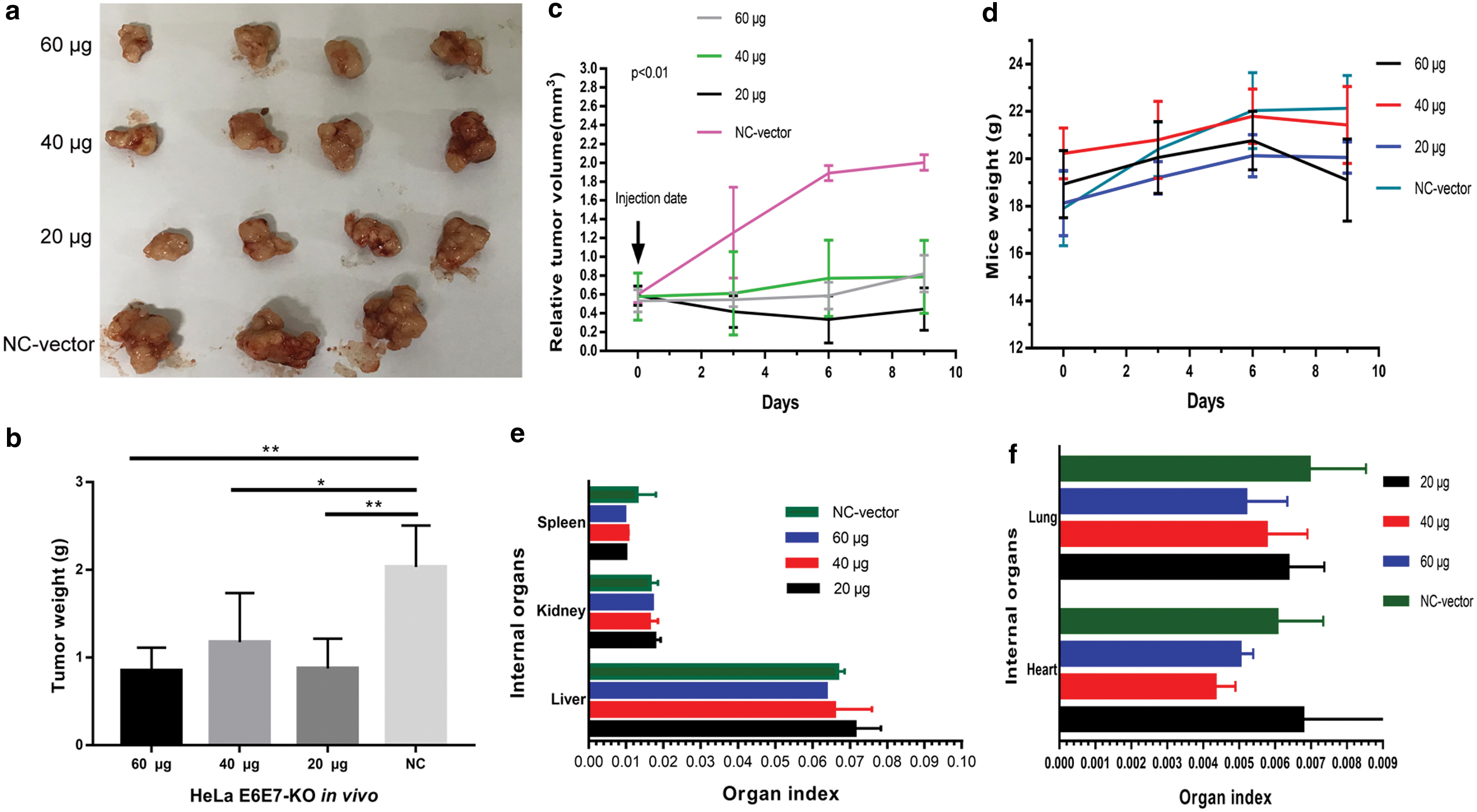
CRISPR/Cas9 E6E7-KO delivery by liposomes suppresses tumor growth in vivo.
Regarding liposome nanoparticles toxicity, the weight and organ index of the mice were compared. The mouse weight did not decrease significantly among the groups (Fig. 7d). For the internal organs, the organ index in the experimental groups showed no difference when compared with control group (Fig. 7e, f). No organ toxicity was observed. Taken together, E6E7-KO inhibited tumor growth without lipid toxicity in vivo. To evaluate the efficacy of E6E7-KO targeting in vivo, and the possible mechanism, the protein expression of E6 and E7 and that of their regulated genes p53/p21 were observed. E6 and E7 expression levels in the experimental groups were reduced when compared with the NC group. Correspondingly, the expression levels of p53/p21 were markedly enriched in the experimental groups, especially in the high-dose group. In addition, p53 protein expressed significantly in experimental groups, which might mean that the antitumor effect was mainly due to the p53 pathway (Fig. 8).
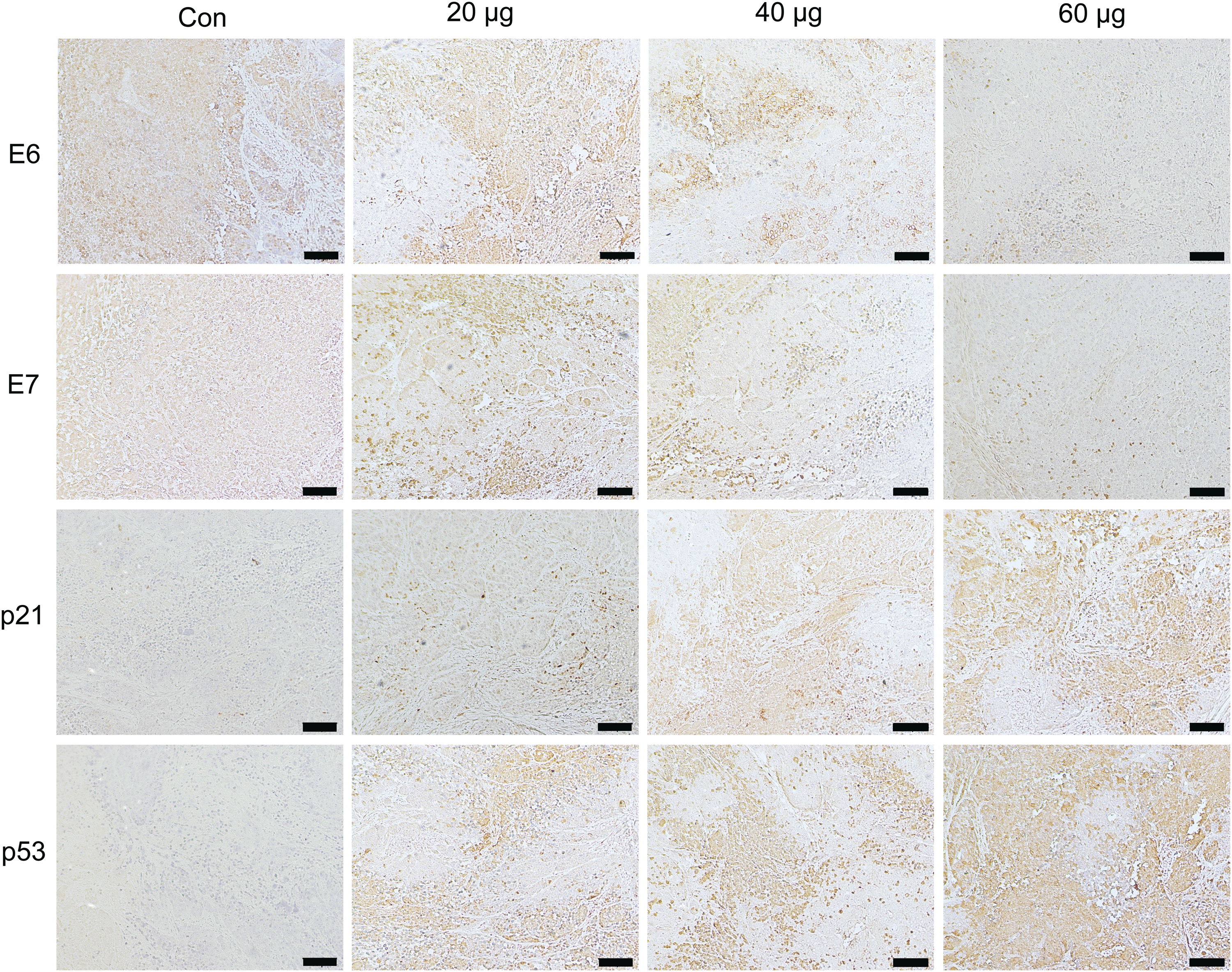
Protein expression was detected by IHC in vivo. IHC indicated that E6 and E7 protein expression in the E6E7-KO groups was reduced when compared with the NC group. p53 and p21 protein expression was enriched in the E6E7-KO groups, particularly in the high-dose group (60 μg). The protein expression of p53 markedly increased in the E6E7-KO groups. Scale bars, 100 μm. IHC, immunohistochemistry; NC, negative control. Color images are available online.
Discussion
Currently, there is a growing interest in gene therapy, which has been developing rapidly with the increased technical progress. The CRISPR/Cas9 system is attractive as a novel gene-editing tool due to its convenience and efficiency. Downregulation or KO of HPV E6 or E7 may be regarded as the ideal therapeutic method for cervical cancer therapy. In this study, a single CRISPR system was constructed to target E6 and E7 and confirmed the effect in vitro. The results were consistent with those of previous studies. 8,18 E6 and E7 mRNA and protein levels were reduced, whereas p53 and p21 proteins accumulated. Consequently, cell apoptosis induction and proliferation inhibition were markedly induced in vitro. Therefore, this study highlights a platform targeting two genes synchronously, and to the best of our knowledge, this plasmid has not been reported for its potential in cervical cancer treatment until now.
All-in-one vector or genome-wide CRISPR-based screening will be a promising blueprint for either research or therapy. 19 Ehrke-Schulz et al. 20 reported that a single viral vector could quickly induce Cas9 and sgRNA expression when it was adapted into high-capacity adenoviral vectors devoid of some viral HPV genes. This process would expand the role of the CRISPR/Cas9 system for HPV-associated disease treatment. However, the constructive approach is required for improvements or remolding. It was speculated that the whole E6 and E7 genome would be cut off if E6E7-KO includes more sgRNAs.
Finally, nonviral delivery or carriers may be potential tools for the clinical genome-editing platform. 21 The CRISPR/Cas9 system delivery by a nonviral carrier, including liposomes, demonstrates improved outcomes for gene therapy. 22 Some studies have confirmed the efficiency of nonviral delivery to be equivalent to that of lentiviral systems, 23 and they represent promising safety and utility of gene editing. 24,25 Nonviral gene delivery could be highly efficient in ex vivo, including that associated with plasmid DNA and mRNA. 26 CRISPR/Cas9 delivery by the nonviral approach has some advantages and could partly avoid various concerns associated with viral-based delivery. Hence, in this study, liposome nanoparticles were used for the nonviral delivery of the CRISPR/Cas9 E6E7-KO plasmid. Concerned about the efficacy in vivo, this study used three different doses to elevate the concentration of the plasmids. All mice treated with E6E7-KO exhibited significant tumor inhibition, especially mice treated with a high dose. IHC revealed that the E6 and E7 protein levels declined. In addition, it was also demonstrated that p21/p53 protein expression markedly increased when treated with E6E7-KO. A limitation of our study was that we should have injected every 3 days regularly in vivo, as serial dosing regimens might eradicate the tumor. Regarding the side effects, the present results suggest that liposome nanoparticles did not show poisonous effects in mice.
In conclusion, the results of this study indicated that HPV-based gene therapy for cervical cancer is feasible, and CRISPR/Cas9 with liposomes or other nanoparticle delivery systems may be applied locally in vivo. Of course, CRISPR/Cas9 also exhibited technical difficulties, including off-target effects and low efficacy of the delivery in vivo. Combination with drugs or other therapies may enhance its function. 18 Therefore, more studies should be performed to make this approach suitable and safe for clinical application in the future.
Footnotes
Acknowledgments
We extend our special gratitude to the technician Tanghong for assistance with our experiment.
Availability of Data and Materials
Data and materials are fully available without restriction.
Authors' Contributions
K.L. and Z.L. conceived and designed the study. K.L., L.Y., S.L., Y.W., P.Y., Y.L., M.C., L.J., and N.Y. conducted the experiments and collected and analyzed the data. K.L., L.Y., and S.L. wrote the original draft. Z.L. reviewed and edited the article.
Ethics Approval
Institutional Review Board of the Southwest Hospital, Third Military Medical University, approved this study.
Author Disclosure
The authors have declared that no competing interests exist.
Funding Information
This study was supported by the National Natural Science Foundation of China (Grant No. 816073011) and the National Key Technology R&D Program of China (Grant Nos. 2019YFC1005202 and 2019YFC1005203).