
Abstract
Regulation of gene expression by viral vectors is an effective method for researchers to explore the function of gene products in a target tissue. The choroid plexus (CP) is an important target for gene therapy of neuropsychiatric diseases such as Alzheimer's disease and major depressive disorder. However, viral tropism in CP has not been well studied as a result of limited viral vector applications. To identify CP-specific viral vectors, we intracerebroventricularly administered six different serotypes of adeno-associated virus (AAV) vectors (AAV2/1, AAV2/5, AAV2/8, AAV2/9, AAV2-BR1, and AAV2-PHP.eB) and lentivirus in adult mice. Tropism in CP was compared among these viruses. We found that AAV2/5 and AAV2/8 displayed remarkable infections in CP, while AAV2/1 infected both ependymal cells and cells in the CP. Except for the low infection intensity of AAV2/9 and lentivirus in the CP, no infection intensity was found for CP tissues injected with AAV2-BR1 or AAV2-PHP.eB. Green fluorescence protein expression in the CP after AAV2/5 infection was confirmed by Western blotting. AAV2/5-mediated tropism in epithelial cells of the CP was verified by immunostaining with transthyretin. In this study, we identified for the first time that serotype-specific AAVs 5 and 8 may be robust research tools for intracerebroventricular gene delivery.
Introduction
Neuropsychiatric disorders such as major depressive disorder, Alzheimer's disease, and schizophrenia impose a severe burden on society. In our previous studies, we found altered metabolites and proteins in the cerebrospinal fluid (CSF) of nonhuman primates (cynomolgus monkey, Macaca fascicularis) with naturally occurring depression in cynomolgus monkeys. 1,2 In addition, gene alterations were identified in the choroid plexus (CP) of depression patients. 3 We also found that dysbiosis of the gut induced depressive-like behaviors by means of the microbiota/gut/brain axis. 4,5
A recent trial showed that CP volume was significantly increased in psychosis probands, 6 suggesting involvement of the CP across the psychosis spectrum with a latent pathophysiological mechanism. However, function of the CP in neuropsychiatric disease remains largely unknown. Moreover, as the CP is exposed to blood and CSF, the secretion of peptides and proteins in the CP affects both blood components in the peripheral environment and CSF components in the central environment. 7 Thus, gene manipulation of the CP is a meaningful way to uncover potential roles of the CP in neuropsychiatric diseases.
To better investigate the function of CP, we describe a viral vector-based approach to make it easier to study the CP at the gene level. Viral vectors, such as adeno-associated virus (AAV) and lentivirus, are currently widely used both in animal model-based neuroscientific research 8 and clinical trials. 9 It was previously shown that lentivirus with a CP-specific promoter can effectively deliver secreted peptides and proteins into the CSF. 10 However, compared with AAV, lentivirus is less used in vivo because of its poor diffusion ability and strong immunogenicity. Moreover, the viral genome of lentivirus may randomly insert into actively transcribed genes of host cells, likely triggering mutagenesis. 11 This instability greatly limits the application of lentivirus in vivo. In contrast, AAV vectors, which can infect both dividing and nondividing cells, are mostly episomal in host cells and apparently lack pathogenicity; thus, AAVs are considered safe for use in vivo. Therefore, we decided to use AAV vectors as an alternative tool for gene intervention in the CP.
The floating structure of the CP makes local injection of virus difficult, which greatly limits methods for gene manipulation in the CP. However, different serotypes of AAV vectors display different tissue-type 12 and cell-type 13 tropisms by recognizing different cell receptors. Thus, it is possible to make use of this property to target the CP via certain serotypes of AAV vectors. In addition, the injection method for virus delivery plays a crucial role in infection efficiency. Systemic delivery of AAV9 and AAV PHP.eB 14 reportedly provides a noninvasive alternative for broad gene delivery to the central nervous system (CNS). 15 Nevertheless, poor specificity of brain regions and low infection rates limit the application of this method in neuroscience. Thus, it is more commonly used to target a specific brain region by local injection.
Based on this approach, we intracerebroventricularly (ICV) administered six different serotypes of AAV vectors (AAV2/1, AAV2/5, AAV2/8, AAV 2-PHP.eB, AAV2-BR1, and AAV2/9) and lentivirus, commonly used in the CNS to identify effective and robust viral tools that could infect the CP and express relevant transgenes.
In summary, we offer a virus-dependent method to manipulate gene expression in the CP and regulate intracerebroventricular substances through CSF secretion, which may be a useful tool for neuropsychiatric investigations.
Materials and Methods
Mice
Our study used C57BL/6 male mice (8–16 weeks; weighing 20–25 g) purchased from the experimental animal center of Chongqing Medical University (Chongqing, China). Animals were singly housed under controlled conditions (12-h light/dark cycle with lights on at 8:00 am, 21–25°C, and 55% ± 5% relative humidity) with ad libitum access to water and food. Permission for all experiments conducted in this study was obtained from the Animal Care and Use Committee of Chongqing Medical University (Approval No. 20160331).
Viral construct and packaging
All viral vectors were produced according to the standard method. 16,17 Briefly, 293T cells were transfected with an AAV vector containing a reporter gene cDNA driven by a cytomegalovirus (CMV) or chicken beta actin (CAG) promoter, helper plasmids containing the AAV2 rep gene, and capsid gene plasmids of different serotypes. Purification of viral particles was conducted with an iodixanol step-gradient ultracentrifugation method. After transfection of the lentivirus transfer vector and helper plasmids into 293T cells, virions from the cellular media went through subsequent purification and were finally harvested. Titers of AAV vectors, as determined by quantitative PCR, were over 1 × 1012 viral genomes (vg)/mL. The titer of lentivirus was over 1 × 109 transduction units (TU)/mL. Vectors used in this study were all single-stranded AAVs.
Stereotaxic surgery
For virus injection, each mouse was fixed in a stereotaxic frame (RWD; Life Science, Shenzhen, China) after being anesthetized with isoflurane. Five microliters of virus (titers of AAVs were ∼1.6 × 1010 vg/mouse; titer of lentivirus was 5.95 × 106 TU/mouse) was slowly injected into the right lateral ventricle [anterior/posterior (AP): −0.4 mm from bregma, medial/lateral (ML): −1.0 mm, dorsal/ventral (DV): −2.0 mm from the brain surface]. AAV2/8 with an SYN promoter (1 μL) was injected into the hippocampus (AP: −3.3 mm from bregma, ML: ±3.0 mm, DV: −1.5 mm/−2.5 mm/−3.5 mm from the brain surface). The pulled glass capillary was left in place for an additional 3 min after injection. The wound was sutured and animals were fed for 21 days until sufficient expression of fluorescent protein could be observed.
Brain slice preparation and fluorescence detection
All brain slices used in this study were processed under the same condition. After 21 days of transgene expression, mice were anesthetized with 1% pentobarbital sodium and transcardially perfused with ice-cold phosphate-buffered saline (PBS; pH 7.4) followed by 4% paraformaldehyde. Next, brains were fixed in 4% paraformaldehyde solution for 6–8 h and then dehydrated in 30% sucrose at 4°C until they sank. Coronal brain sections (20 μm) were cut on a freezing microtome (Leica, Wetzlar, Germany) and mounted on adhesive microscope slides. Brain sections were stored at −80°C before fluorescence detection and immunostaining. Fluorescent signals of all brain slices were detected under the same exposure settings after staining with 4′,6-diamidino-2-phenylindole (DAPI). All images were acquired under a Nikon A1R confocal microscope (Tokyo, Japan).
Immunofluorescence
Brain sections were first incubated with primary antibodies at 4°C overnight. After washing with PBS three times (10 min each), sections were incubated with an appropriate secondary antibody at room temperature for 2 h. Antibodies used in this study were rabbit anti-green fluorescence protein (GFP) (1:200; GeneTex, Irvine, CA), rabbit anti-FLAG (1:200; GeneTex), rabbit anti-transthyretin (TTR; 1:100; Proteintech, Rosemont, IL), Alexa Fluor 488 donkey anti-rabbit IgG, Alexa Fluor 555 donkey anti-rabbit IgG, and Alexa Fluor 555 donkey anti-mouse IgG (1:1,000; Invitrogen, Carlsbad, CA).
CP tissue isolation
Mouse brains were collected and quickly immersed in ice-cold PBS after animals were anesthetized with 1% pentobarbital sodium. CP in the lateral ventricles was carefully isolated with sharp forceps under a stereomicroscope 18 and then rapidly frozen in liquid nitrogen. All tissues were stored at −80°C until use.
Western blotting
Western blotting was used to evaluate the expression of the fused fluorescent protein. As reported in our previous study, 19 CP tissue was lysed with radioimmunoprecipitation assay buffer (Beyotime, Nanjing, China) containing protease inhibitor cocktail (Roche, Basel, Switzerland). After separation by 10% sodium dodecyl sulfate/polyacrylamide gel electrophoresis, proteins were transferred to polyvinylidene difluoride membranes (Millipore, Billerica, MA) for 2 h of blocking. Next, membranes were incubated with rabbit polyclonal anti-GFP (1:10,000; GeneTex) and mouse anti-GAPDH (1:10,000; Proteintech) overnight at 4°C. After washing with tris-buffered saline containing Tween 20 (Sigma-Aldrich, St. Louis, MO), membranes were incubated with secondary antibodies at room temperature for 2 h. Proteins were detected with horseradish peroxidase-conjugated secondary antibodies using an ECL Kit (Millipore). Signals were visualized using ChampChemi (Sage Creation Science, Beijing, China).
Results
Tropism of different viral vectors in CP
We constructed six serotypes of AAV vectors and lentivirus, which were all driven by broad-spectrum promoters (Table 1). To infect the CP, we ICV injected these viruses independently. Intracerebroventricular administration was chosen to directly expose the CP to these viruses. To reduce the effect of CSF on diluting viruses, we ICV injected 5 μL of each vector. Brain slices of each group were imaged at 21 days postinjection. Images of different groups in all fluorescent channels were acquired under identical exposure settings. The results showed strong and obvious fluorescence signals in the CP after intracerebroventricular injection of AAV2/1, AAV2/5, and AAV2/8, but there was no observable fluorescent protein expression (GFP) after injection of AAV2-BR1 or AAV2-PHP.eB (Fig. 1). In addition, CP infected by AAV2/9 and lentivirus exhibited a low fluorescent intensity (Fig. 1).
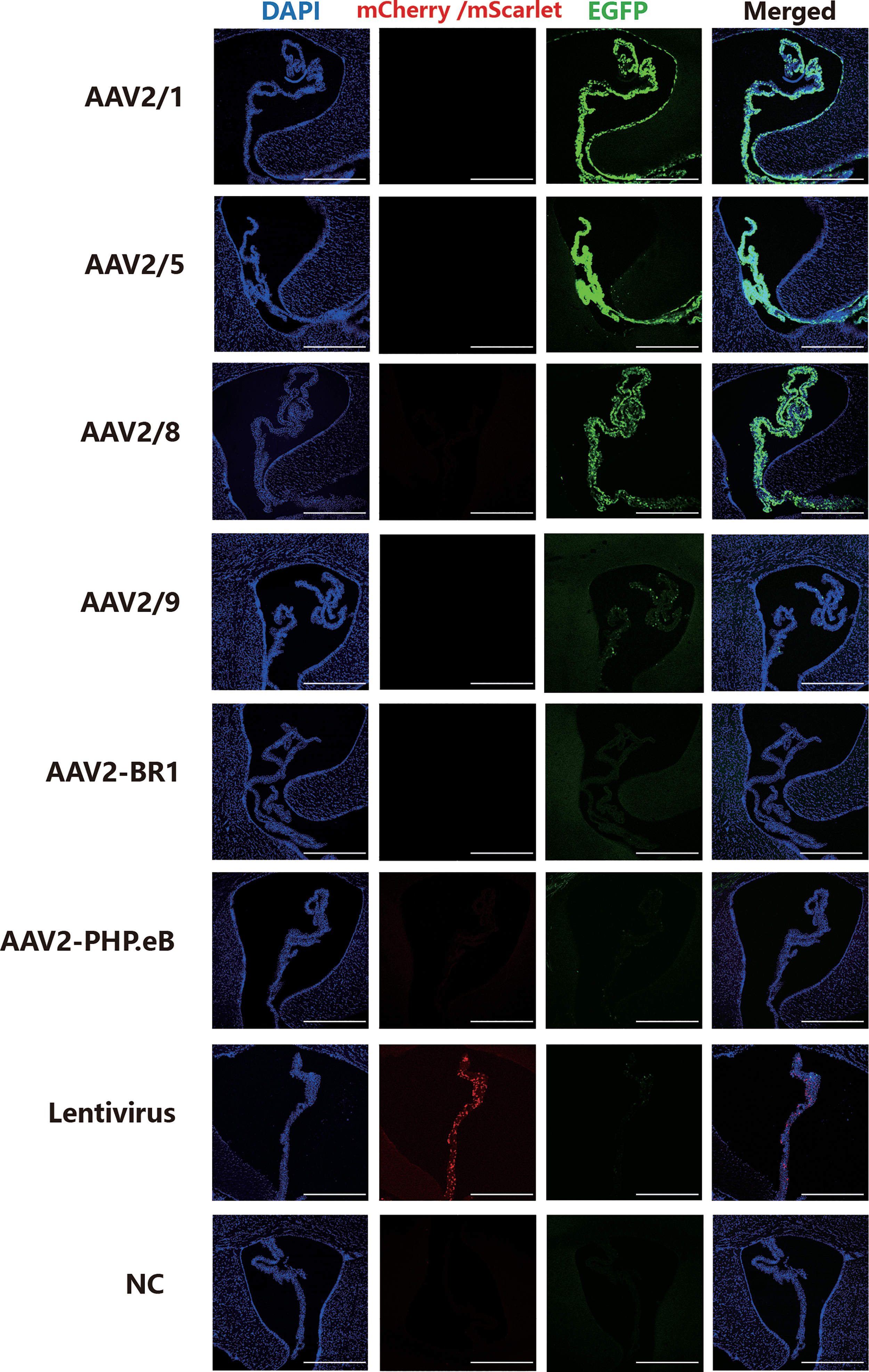
Tropisms of six serotypes of AAV vectors and lentivirus in CP. Fluorescent images of the distribution of report gene expression 21 days after viral vector injection. High-infection group: AAV2/1, AAV2/5, and AAV2/8; low-infection group: AAV2/9, lentivirus; no-infection group: AAV2-BR1 and AAV2-PHP.eB. Equal volume of PBS was injected as a negative control. Scale bar: 500 μm. n = 3 mice per group. AAV, adeno-associated virus; CP, choroid plexus; NC, negative control; PBS, phosphate-buffered saline. Color images are available online.
Detailed vector information of the viruses
AAV, adeno-associated virus; CAG, chicken beta actin; CMV, cytomegalovirus; EGFP, enhanced green fluorescence protein; Ubc, human ubiquitin C.
To magnify fluorescence signals of viruses and avoid the quenching of fluorescence during experimental processes, immunostaining was used to detect protein expression of the relevant transgene in the low-infection and noninfected group. The results of immunostaining with a DDDDK tag (FLAG) antibody indicated little positive fluorescence signal in AAV2-PHP.eB- and AAV2-BR1-infected CP (Supplementary Fig. S1). Moreover, little GFP expression was found in AAV2/9-infected CP (Supplementary Fig. S1). Hence, we confirmed that AAV2/9, AAV2-BR1, and AAV2-PHP.eB failed to effectively infect the CP. Whole-brain images of all the serotypes were also provided (Supplementary Fig. S2).
Infection characteristics of AAV2/1, AAV2/5, AAV2/8, and lentivirus in CP
After costaining with a nuclear marker (DAPI), we confirmed that intracerebroventricular administration of AAV2/1 resulted in GFP expression both in ependymal cells (which line the brain ventricles) and CP cells (Fig. 2a). Even in the third ventricle, strong fluorescent signals were also observed in these cells (Fig. 2a). In contrast, intracerebroventricular administration of AAV2/5 and AAV2/8 specifically infected CP cells rather than ependymal cells (Fig. 2b, c). Imaging of the whole brain clearly demonstrated that only CP cells, not ependymal cells or other surrounding regions of CP, expressed GFP in AAV2/5-injected mice (Fig. 2b).

Infection characteristics of AAV2/1, AAV2/5, AAV2/8, and lentivirus in CP.
Lentivirus also effectively infected CP cells, although the infection intensity was not as good as the aforementioned viruses (Fig. 2d). We also found that AAV2/8 driven by a synapsin (SYN) promoter instead of CMV was unable to effectively infect CP, although AAV2/8 with an SYN promoter successfully infected neurons in the hippocampus as a positive control (Fig. 2e). As both epithelial cells of the CP and ependymal cells are glial cells, it is reasonable that AAV2/8 under a neuronal SYN promoter resulted in negative transduction of the CP. Thus, both serotype and promoter codetermined viral tropism in tissue.
Validation of stable and long-term infection in CP after AAV2/5 injection
First, after 21 days of AAV2/5 infection, we carefully isolated the CP under a microscope (Fig. 3a). Next, expression of GFP in CP was validated via Western blotting. The hippocampus, which is structurally adjacent to the CP, was compared as a negative control. As expected, GFP was expressed only in the CP and not in the hippocampus (Fig. 3b). We then examined whether there were epithelial cell tropisms of AAV2/5 infection in the CP. First, anatomical structure of the CP was observed (Fig. 3c). To test our hypothesis, we used TTR (an important transporter synthesized and secreted by CP epithelial cells in mouse brains) to conduct immunostaining in AAV2/5-infected CP. The results showed colocalization of TTR and GFP, illustrating that AAV2/5 was able to infect epithelial cells of CP (Fig. 3c). In addition, after intracerebroventricular administration of AAV2/5 at titers of 4.55 × 1010, 1.37 × 1011, and 2.28 × 1011 vg/mouse, we found that different titers of viral vectors showed differences in infection intensity, demonstrating that AAV2/5 infection of CP had a dose-dependent effect (Fig. 3d). Evident GFP fluorescent signal in the CP was revealed both in lateral ventricles and the third ventricle, even 6 months after intracerebroventricular administration of AAV2/5, indicating stable and long-term transgene expression in the CP (Fig. 3e). Moreover, we were surprised to find GFP expression in ependymal cells 6 months after ICV injecting AAV2/5, which was different from the result observed 21 days after intracerebroventricular injection (Fig. 3e).
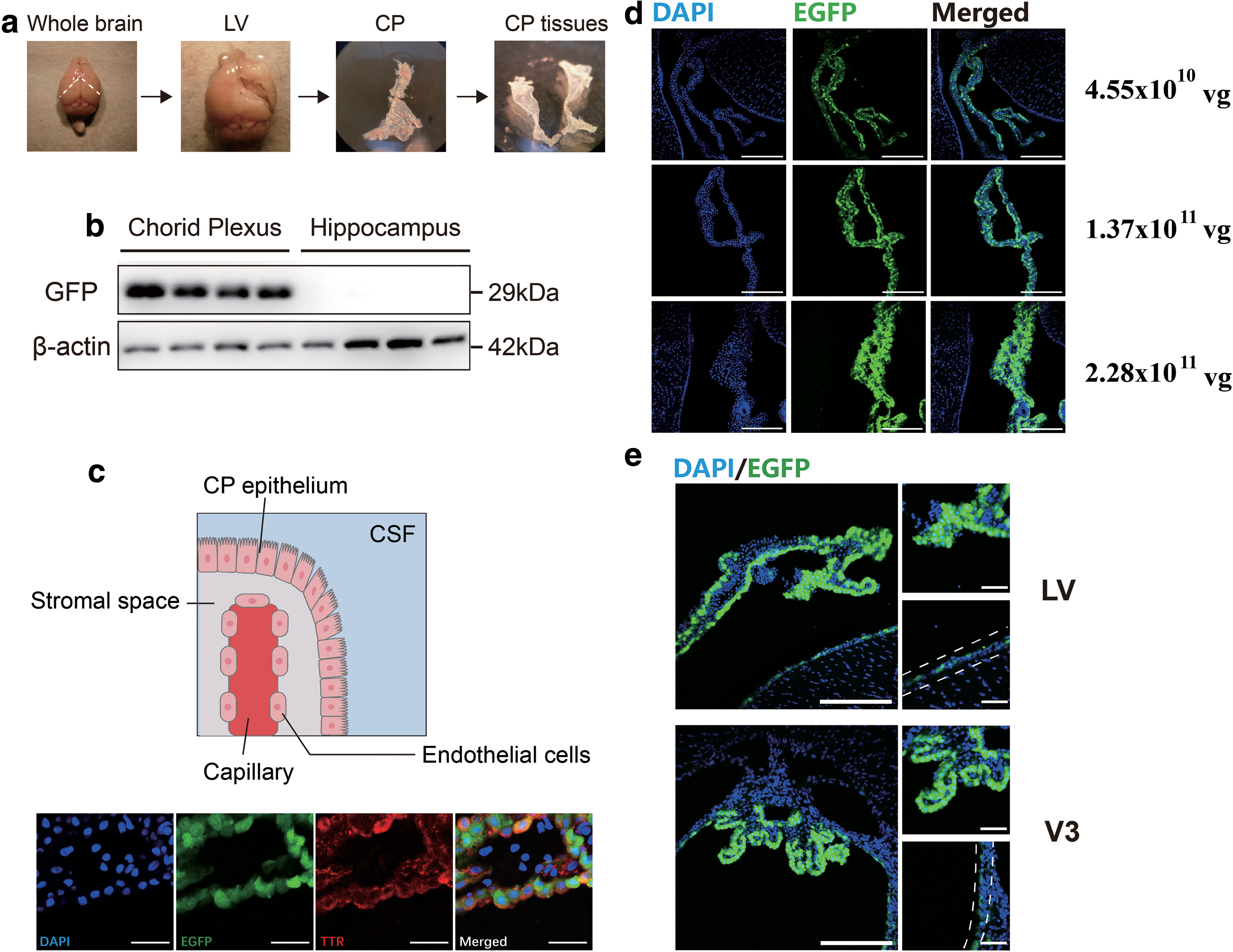
Validation of the persistent infection of AAV2/5 in CP.
Discussion
Despite an abundance of studies 6,20,21 reporting molecular changes in the CP of neuropsychiatric patients, few studies have investigated potential physiological mechanisms in the CP underlying neuropsychiatric disorders. To address this issue, we validated reliable serotypes of viral vectors to offer a method to intervene in gene expression in CP, which might be valuable for further applications in gene therapy of neuropsychiatric diseases.
In this context, we compared the tropism of different serotypes of viral vectors in the CP of adult mice for the first time. AAV2/5 and AAV2/8 were confirmed to be effective among the six examined serotypes of AAV vectors (AAV2/1, AAV2/5, AAV2/8, AAV 2-PHP.eB, AAV2-BR1, and AAV2/9) and lentivirus. Consistent with a previous study, 10 we verified that lentivirus was able to infect CP. However, considering the immunogenicity and low infection intensity of lentivirus, we here provided evidence that the prevailing AAV vectors (especially AAV2/5 and AAV2/8) were effective and robust tools to replace lentivirus for specific manipulation of gene expression in the CP.
It was previously reported that both AAV1 22 and AAV4 23 were able to infect ependymal cells and constantly express a transgene in the CSF after intracerebroventricular injection. Our findings confirmed AAV1-mediated infection in the CP and ependymal epithelium. Intracerebroventricular delivery of AAV5 24 in the prenatal period led to transgene expression in the CP, while delivery of AAV9 tended to spread throughout the brain. 25 Our results showed that AAV9 was unable to effectively infect the brain parenchyma of adult mice because of the maturation of ependymal cells, which prevented AAV9 from transferring from the CSF to the brain. Although AAV5 infection of CP was observed after intrathecal injection in adult mice, 26 low specificity and technical difficulties limit the wide use of this method.
Moreover, although AAV2-PHP.eB and AAV2/9 are widely used to cross the blood–brain barrier through tail vein injection 27 –29 —we here presented evidence that they failed to infect CP effectively when ICV injected. AAV2-BR1 was reported to have a high specificity for endothelial cells of the brain and spinal cord vasculature via intravenous injection, 30 however, we did not find observable transduction of endothelial cell in the CP by intracerebroventricular injection. Tropism of AAV2/5 in epithelial cells, rather than ependymal cells, disappeared after 6 months of infection. This phenomenon probably resulted from varying infection efficacies of each vector in different cell types.
Despite the high cost of manufacturing viral vectors and existence of immunological barriers in the human population, 31 AAV-centric gene intervention via gene addition and replacement has been conducted in multiple clinical trials of neuropsychiatric diseases. For example, AAV-mediated glutamic acid decarboxylase delivery in subthalamic nuclei of patients with Parkinson's disease reportedly elicited significant clinical benefits persisting for 12 months. 32
Considering the mounting evidence for involvement of the CP in inflammatory-driven neuropsychiatric disorders, 33 treatments to directly target CP cells, such as the delivery of compounds to reduce brain inflammation and leukocyte brain entry, 34 might help. Moreover, transplantation of CP epithelial cells can reportedly provide neuroprotective effects. 35 Based on our work, the success of this approach suggests that AAV-mediated gene therapy in CP is viable. Indeed, infection of CP with viral vectors enables persistent gene delivery in vivo, providing a new strategy for treatment of neuropsychiatric diseases.
The CNS is susceptible to disturbances in peripheral blood through the dysfunction of CP. Hence, neuropsychiatric symptoms are also found in manifold non-neuropsychiatric diseases. Human lupus patients with neuropsychiatric symptoms exhibited increased leukocyte migration through the CP in histological evaluation. 36 The CP itself plays a vital role in multiple pathophysiological processes. Age-related cognitive decline in mice could be reversed by blocking type I interferon signaling in CP. 37 Moreover, the CP is an essential structure in the migration of peripheral inflammatory cells toward the CNS; for example, increased lymphocyte adhesion in the vessels and stroma of CP has been observed in multiple sclerosis patients. 38 Therefore, the CP is also a candidate target for gene therapy and genetic manipulation in the study of non-neuropsychiatric diseases with psychiatric manifestations.
However, there are limitations that we only examined viral vectors without target genes. The results would be more convincing if we had explored functional changes in CP after infection with vectors carrying a specific target gene. In addition, as the present experiments were conducted in mice, it remains to be seen whether similar conclusions occur in other species. Moreover, while we verified epithelial cell tropism of AAV2/5 in infecting the CP through colabeling with TTR, endothelial cells were not analyzed. Ideally, it would be more credible if vectors in this study were under an identified promoter. However, our findings emphasize the potential of AAV2/5 and AAV2/8 for gene operation of the CP. Future studies are needed to prove that this AAV-based method of gene transfer into the CP can effectively interfere with its function.
Taken together, the results of this study revealed the different tropisms of viral vectors in CP, demonstrating that AAV2/5 and AAV2/8 are effective viral tools to intervene in gene expression in CP. Owing to the CSF secretory function of CP, this AAV-mediated tool may help establish a prospective method for drug delivery in the CNS. Furthermore, modulating gene expression in CP by means of viral vectors is a promising method for biological and clinical applications in the future.
Footnotes
Acknowledgments
We thank Dr Shanquan Sun from the Chongqing Medical University for consultation on anatomical structure and functional character of CP.
Author Disclosure
No competing financial interests exist.
Funding Information
This study was supported by the National Key R&D Program of China (2017YFA0505700) and the National Key Program International Cooperation Project (No. 81820108015).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.